
Abstract
Intermittent fasting can be effective in reducing metabolic disorders and age-related diseases. However, there remain questions about the effects of fasting with respect to the age in which fasting begins, the fasting models, and the mechanisms involved. We investigated the effects of age of beginning fasting and chronic mild and severe fasting models on blood pressure (BP), insulin/glucose profile, and expression of klotho, sirtuin1 (SIRT1), and sirtuin3 (SIRT3) in male Wistar rats. Young (3 months), middle-aged (12 months), and old (22 months) animals were randomly divided into three subgroups and fed as ad libitum (AL), AL with fasting 1 day per week (FW), and AL with fasting every other day (EOD), respectively, for 3 months. The FW reduced the weight gain in young animals (p < 0.001 vs. AL), whereas EOD induced weight loss in all three age categories (p < 0.001). Aging was associated with high BP, high glucose, and insulin levels. Both FW and EOD feedings decreased BP and blood glucose level (p < 0.001) and EOD decreased insulin level (p < 0.05 vs. AL) in old animals. Parallel to aging, the expression of SIRT1 and klotho significantly decreased in plasma and EOD feeding recovered this defect. Both FW and EOD feedings increased the expression of SIRT3 in middle-aged and old rats. Age is a determining factor for the effectiveness of fasting and old animals respond more desirably to fasting. The effect of EOD fasting is more effective than FW fasting in improving the metabolic factors, partly through the recovery of SIRT1 and klotho.
Introduction
Aging is considered as one of the major risk factors for the development of metabolic and cardiovascular disease. 1,2 Some interventions, such as caloric restriction (CR) or CR mimetic agents like metformin, spermidine, and rapamycin, and also non-CR interventions, such as angiotensin II blockade, may increase survival and retard age-related diseases in mammals and nonmammal organisms. 3,4 Intermittent fasting (IF) is one of the methods of energy deprivation, 5 which is distinct from CR. Unlike CR, the energy intake through IF does not need to be limited; in turn, the frequency of food intake is controlled. 6
It is reported that intermittent or periodic fasting protects and delays the onset of metabolic and age-related diseases, such as diabetes, cancers, heart disease, neurodegeneration, obesity, hypertension, asthma, and rheumatoid arthritis in rodents and human. 7 It increases longevity 7,8 and minimizes the side effects caused by chronic dietary regimens. 7 IF increases the myocardial tolerance and protects the rat heart against ischemic injury, besides being able to attenuate post-myocardial infarction cardiac remodeling likely through antiapoptotic and anti-inflammatory effects. 9 In addition, IF can reduce the heart rate (HR) and blood pressure (BP), 10 and can improve cardiac diastolic parameters. 11 It is also known to extend the lifespan in mice and rats. 12,13
The signaling pathways of CR are well studied and a part of its beneficial outcomes on longevity, improving lifestyle and decreasing cardiovascular and metabolic disorders are attributed to the improving of sirtuins gene/protein expression 14 –17 and antiaging gene of klotho expression. 18 Sirtuins are NAD+-dependent protein deacylases that cleave off acetyl, as well as other acyl groups, from ɛ-amino group of lysines in proteins like histones. 19 They regulate important biological pathways, such as mitochondrial biogenesis, energy efficiency, aging, transcription, apoptosis, inflammation, and stress resistance. 20
It is suggested that sirtuins, especially Sirt1, 2, 3, 6, and 7, can be some of the mediators of the antiageing effects of calorie restriction in skeletal and cardiac muscles. 21 Klotho is a protein with multipotential effects. For instance, it suppresses the insulin and Wnt signaling pathways, inhibits oxidative stress, and regulates phosphatase and calcium absorption, and induced antiaging traits. 22,23 Kidneys are the principal providers of klotho, which is intended to circulate in the bloodstream. 23 Due to its antiapoptotic and antioxidant properties, klotho is considered as a novel protective factor in the cardiovascular system. 24
IF is not only more acceptable to patients, but it also prevents some of the adverse effects of chronic calorie restriction, such as malnutrition, anemia, weakness, and muscle wasting. However, molecular mediators and mechanisms, which involve the effects of IF on mammals, need to be investigated further, as there is little information in this regard. We hypothesized that klotho and sirtuins may change during fasting and in turn, mediate its beneficial effects on improving metabolic and cardiovascular systems. Therefore, the present study was conducted to test this hypothesis and also evaluate the effect of ages in which two fasting models begins, on BP, insulin/glucose profile, and mentioned molecules. Young, middle-aged, and old male rats were subjected to two model of chronic mild (fasted 1 day per week [FW]) and severe (every other day [EOD]) IF. Then, BP, genes, and protein expression of klotho, sirtuin1 (SIRT1), and sirtuin3 (SIRT3) in the heart, kidney, plasma, and also insulin and blood sugar were measured.
Materials and Methods
Materials
Chemical materials were prepared as Urethane from Sigma-Aldrich Co., the QuantiTect Reverse Transcription Kit (Qiagen, GmbH), the Qiagen RNeasy Mini Kit (Qiagen, GmbH), SYBR Green Supermix (BioFACTCO, South Korea), sirtuins, the α-Klotho and Insulin ELISA Kit (Eastbiopharm; Hangzhou, China), and the Glucose ELISA Kit (Pars Azmon, Tehran, Iran). Specific forward and reverse primers were selected and designed by Sinaclon CO (Tehran, Iran).
Ethics approval
This study was conducted in accordance with the national laboratory practice guidance and was approved by the Ethics Committee (permission No IR.KMU.REC.1395.313) of the Kerman University of Medical Sciences, Kerman, Iran.
Animals and diet
Male Wistar rats with three initial different ages (3, 12, and 22 months) were obtained from the Animal House of Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran. Upon arrival, all rats were maintained on a standard diet and had free access to water, along with a 12-h light/12-h dark cycle. Ten days later, animals were randomly assigned to standard laboratory food ad libitum (AL), feeding AL and FW, and feeding AL and fasting EOD. The rats were housed separately in isolated cages (one rat per cage) under controlled conditions of optimum temperature (22°C) and, food was provided or removed at 8.30 a.m. every day. Body weights were recorded weekly and the fasting protocol lasted for 3 months before the experimental examination.
Sampling and recording
Twenty-four hours after the completion of the course, fasting blood samples were collected from the tail vein of each rat and drawn into a tube containing K3EDTA as an anticoagulant. The plasma was isolated and stored at −80°C for measuring insulin, glucose, SIRT1, SIRT3, and α-klotho. The next day, the animals were anaesthetized with the urethane (Sigma) (1000 mg/kg body weight) and placed on a heated surgical table to maintain rectal temperature at 37°C. The animal's trachea was cannulated to allow free breathing without any obstruction. The carotid artery on one side was exposed, and a polyethylene catheter was inserted into the artery for recording arterial BP. The catheter was attached to a pressure transducer (Statham-P23D). The patency of the catheter was maintained with a heparinized saline solution (5000 IU m/L, v/v). The pressure recording system was calibrated with the help of a mercury manometer before each experiment. Systolic, diastolic, mean arterial pressure (MAP), and HR were recorded on a Power-Lab (AD Instruments, Australia) data acquisition system with a computerized analysis program. Rate pressure product (RPP) was calculated as: RPP = (HR × systolic pressure)/1000.
Preparation of tissues
After recording, the animals in each group were sacrificed to have their organs collected. The whole heart and kidney cortex were removed and immediately frozen on dry ice, then stored at −80°C, and they were processed for further experiments.
Quantitative real-time PCR
We used real-time PCR (RT-PCR) to determine the transcriptional induction of antiaging genes, including SIRT1 and SIRT3. About 30 mg of the samples obtained from the heart and kidney was homogenized and total RNA was extracted according to the RNA Isolation Protocol of the manufacturer's instruction (Qiagen, GmbH). The total RNA was extracted from the frozen heart and kidney tissue samples using the Qiagen RNeasy Mini Kit (Qiagen, GmbH). The purity and concentration of the extracted RNA were determined spectrophotometrically at 260 and 280 nm wavelengths (Eppendorf; BioPhotometer Plus, Germany). Then, strand cDNA was synthesized from 1 μg total RNA using the QuantiTect Reverse Transcription Kit (Qiagen, GmbH) according to the manufacturer's instruction. After determining the concentration and purity of total RNA by a NanoDrop spectrometer (Eppendorf; BioPhotometer Plus), extracted RNA was reverse transcribed using the QuantiTect Reverse Transcription Kit (Qiagen, GmbH) in gradient thermal cycler in a final volume of 10 μL according to the manufacturer's protocol. All PCR amplifications were performed in duplicate reactions and in final volume of 10 μL containing 1 μL of diluted cDNA, 5 μL SYBR Green Supermix (BioFACT CO, South Korea), and 3.2 μL deionized H2O and specific forward and reverse primers with a final volume of 0.8 μL selected and designed by Sinaclon CO, IRN. The sequences of primers used in this study are shown in Table 1. PCR was carried out in a StepOne Plus Real-Time PCR System (Applied Biosystems) using the following protocol: incubation at 95°C for 10 minutes to activate DNA Taq polymerase, 40 two-step cycles with 15 seconds at 95°C for denaturation, and annealing/extension at 60°C for 1 minutes. In addition, the no-template negative control was routinely run in every PCR. The melting curve was examined at the end of the amplification process to ensure the specificity of PCR products.
Sequences of the Different Primers
SIRT1, sirtuin1; SIRT3, sirtuin3; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
The relative levels of SIRT1 and SIRT3 mRNAs were obtained from a standard curve generated using serial dilution of cDNAs. Glyceraldehyde 3-phosphate dehydrogenase was used for normalization. To determine the relative quantification of gene expression, the comparative cycle of threshold (Ct) method was used with arithmetic formulae (E−ΔΔCt). 25
Enzyme-linked immunosorbent assay
The levels of SIRT1, SIRT3, α-klotho, insulin, and glucose in the blood plasma were assayed by commercially available Enzyme-Linked Immunosorbent Assay (ELISA) Kits. The ELISA tests were processed according to the manufacturer's instructions on the basis of the biotin double-antibody sandwich technology. Briefly, 100 μL of standard, diluted samples (1:5 in sample diluent) was added to the well of a ready-to-use microtiter plate coated with rat protein monoclonal antibody. Then, the plate was incubated for 60 minutes at 37°C. The seal plate membrane was carefully removed, the liquid was drained and the remaining liquid was shook off, and then each well was filled with the washing solution. After washing, 100 μL of chromogen solution A and B were added first to each well and then were incubated for 10 minutes at 37°C away from light to allow color development. Then, 50 μL of the stop solution was added and absorbance was read at 450 nm.
A standard concentration/absorbency curve was generated for each batch of samples and protein concentrations were calculated with a standard curve ranging from 0 to 48 ng/mL using the optical density detected by a Microplate Reader (ELX808; BioTek).
Statistical analysis
All data are expressed as the mean ± standard error of the mean. The data were analyzed by using the Statistical Package for the Social Sciences (SPSS) software program, version 22.0. The values were first assessed for normality using a Shapiro/Wilk test. Weight change comparisons during study were tested by one-way analysis of variance (ANOVA) and difference between the basal and final weights in each group was measured by paired sample t-test. Since the results were not normality distributed, the data were analyzed using a Wilcoxon test.
Other data were analyzed with two-way ANOVA followed by Bonferroni post-hoc test and one-way ANOVA. First, two-way ANOVA was carried out to evaluate the effects of age, diet, and their interactions. When one or both of the main effects were statistically significant, the one-way ANOVA was used for determining the differences between the means. When only the interaction between the diet and age was statistically significant according to the two-way ANOVA model, the one-way ANOVA was carried out to compute pairwise comparisons between groups.
Results
The effects of age and different patterns of fasting on body weight
Young and middle-aged animals on the AL regimen showed 41.91% (p < 0.05) and 2.52% weight gain, respectively, at the end of the 3-month period of experiment (Fig. 1A, B). However, the weights of old animals that fed on the AL regimen had no significant difference when compared with its base weight (Fig. 1C). FW fasting suppressed the slope of weight gain, however, induced 22.73% weigh gain in young animals at the end of 3 months. The patterns of weight changes in middle-aged and old animals on the FW fasting were somewhat similar to the AL regimen. The EOD fasting was associated with significant weight loss almost from the third to fourth week onward in all three age categories of animals, that is, young, middle aged, and old (p < 0.05 vs. corresponding base weight, at the end of 3 months). Between groups, the comparison at each age level revealed that the basal weights had no significant difference. In young animals, FW fasting induced a lower weight gain than AL (p < 0.001) but caused a higher weight gain than EOD (p < 0.001) regimens. However, 3 months of FW fasting had no significant effect on weight of middle-aged and old rats. On the other hand, in comparison with AL and FW regimens, EOD fasting significantly decreased the body weight in the three age groups in our study (p < 0.001 vs. corresponding AL and FW and middle groups, p < 0.01 vs. corresponding AL and FW old group (Fig. 1A–C). The percent change in body weight also is shown in Figure 2. In the presence of both FW and EOD regimens, old animals' group showed the greatest decrease in body weight (p < 0.001 and p < 0.01 vs. young group and p < 0.05 and p < 0.01 vs. the middle-aged group, respectively) (Fig. 2). In comparison with the AL regimen, the FW and EOD regimens were associated with decrease in total food consumption (−4.6% and −30.3%, respectively, for young; −3.1% and −31.7%, respectively, for middle-aged; and −3.9% and −36.3%, respectively, for old animals).

Effect of age and different patterns of fasting on body weight in

Effect of different patterns of fasting on percent change in body weight of animals with different age. AL, ad libitum regimen; FW, AL and fasted 1 day per week; EOD, fed AL and fasted every other day. Data are represented as mean ± SEM, analysis was done by one-way ANOVA, 10–12 animals per group, **p < 0.01 and ***p < 0.001 versus corresponding young group. # p < 0.05 and ## p < 0.01 versus corresponding middle-aged group.
The effects of age and different patterns of fasting on HR, BP, and RPP
Two-way ANOVA indicated the significant effect of diet pattern (df = 2, F = 3.23, p < 0.05) on HR. In comparison with the AL regimen, only the FW feeding decreased the HR, whereas the effect of age as well as the interaction between the age and the diet treatment were not significant. However, for systolic blood pressure (SBP), diastolic blood pressure (DBP), and mean arterial pressure the two-way ANOVA showed significant effect for age (df = 2, F = 12.50, p < 0.001 for SBP; df = 2, F = 3.48, p < 0.05 for DBP; and df = 2, F = 4.83, p = 0.012 for MAP), diet (df = 2, F = 52.11, p < 0.001 for SBP; df = 2, F = 9.61, p < 0.001 for DBP; and df = 2, F = 21.24, p < 0.001for MAP), and age–diet treatment interaction (df = 4, F = 13.57, p < 0.001 for SBP; df = 4, F = 3.52, p = 0.012 for DBP; and df = 4, F = 3.78, p = 0.009 for MAP).
In comparison with the AL regimen, the FW and EOD feeding had no significant effect on the hemodynamic indices of young animals. In the middle-aged group, BPs were not affected by different fasting patterns. However, all of these parameters (SBP, DBP, and MAP) reduced significantly in old animals which experienced FW fasting (p < 0.001 for all of them, vs. age-matched AL group). A similar pattern was observed for BP and RPP when old animals fasted EOD (p < 0.001 for BP, p < 0.05 for RPP) (Table 2).
Effect of Different Patterns of Fasting and Also Age on Blood Pressure and Heart Rate
Data are represented as mean ± SEM; N = 6–8.
p < 0.05, ** p < 0.01 and *** p < 0.001 versus age-matched AL group.
p < 0.005 versus age-matched FW group.
p < 0.05 and ‡ p < 0.01 versus regimen-matched young group.
p < 0.01.
HR, heart rate; SBP, systolic blood pressure; DBP, diastolic blood pressure; MAP, mean arterial pressure; RPP, rate-pressure product; AL, ad libitum; FW, fasted 1 day per week; EOD, every other day; SEM, standard error of the mean.
Among the animals with the AL diet pattern, old rats were hypertensive and had higher MAP, systolic and diastolic pressure (p < 0.01 vs. young and middle-aged). In the EOD fasting condition all three age groups of animals were normotensive and there was no significant difference between the mentioned parameters (Table 2).
The two-way ANOVA indicated significant effects of diet treatment (df = 2, F = 9.12, p < 0.001), and age–diet treatment interaction (df = 4, F = 3.67, p = 0.010) on RPP. On the other hand, age factor had no significant effect on RPP.
As shown in Table 2, the RPP decreased in EOD middle-aged rats compared with the age-matched AL group (p < 0.05), and in FW old rats compared with the age-matched AL group (p < 0.05).
The effects of age and two patterns of fasting on glucose and insulin of plasma
At the end of the study, the plasma levels of fasting glucose and insulin were assessed in three age categories of rats.
The two-way ANOVA showed that plasma glucose level is significantly affected by diet (df = 2, F = 17.98, p < 0.001), and also indicated age–diet treatment interaction (df = 4, F = 13.93, p < 0.001).
In comparison with the AL regimen, the FW and EOD feeding patterns had no significant effect on circulating glucose levels of young and also middle-aged animals. However, both FW and EOD significantly decreased the levels of glucose (p < 0.001 vs. AL group) in old rats. In addition, the value of this parameter was lower in EOD than in the FW group. Comparison between three age categories with AL food pattern showed a higher level of glucose in old rats (p < 0.001 vs. young and middle-aged animals) (Fig. 3). FW and EOD food patterns abolished these aging-dependent difference, so that the level of glucose was lower in old rats treated with EOD fasting than in old rats of the AL group (p < 0.05) (Fig. 3).

Effect of different patterns of fasting and age on blood
For plasma insulin level, the two-way ANOVA showed significant effect by diet (df = 2, F = 4.21, p < 0.05), but not by age or diet–age interaction. The EOD feeding decreased plasma insulin concentration in old animals only (p < 0.05, in comparison with matched AL regimen) (Fig. 3).
The effects of age and different patterns of fasting on gene expression and plasma protein levels
The two-way ANOVA showed significant increase in the expression of SIRT1 mRNA in the heart as a result of the different patterns of diet (df = 2, F = 4.78, p = 0.012). However, different groups of age (df = 2, F = 2.55, p = 0.085) and age–diet patterns (df = 4, F = 4.78, p = 0.488) did not lead to any significant differences in the expression of SIRT1 mRNA in heart tissue.
Analysis showed a main effect of diet (df = 2, F = 92.69, p < 0.001), and an age–diet interaction (df = 4, F = 10.81, p < 0.001) on SIRT1 mRNA in the kidney. A main effect of age (df = 2, F = 13.05, p < 0.001), a main effect of diet (df = 2, F = 17.72, p < 0.001), and an age–diet interaction (df = 4, F = 8.12, p < 0.001) were observed in the plasma level of SIRT1.
When young animals were maintained on the FW or EOD, the heart expression of SIRT1 mRNA and also SIRT1 protein in plasma did not change significantly. However, the SIRT1 mRNA was higher in kidney (p < 0.001) in comparison with the corresponding AL group. In middle-aged rats under FW, only the expression of SIRT1 mRNA in kidney was more than that of the corresponding AL group (p < 0.001). EOD significantly increased the SIRT1 protein in plasma and SIRT1 mRNA of kidney of middle-aged rats (p < 0.001 vs. age-matched AL group). In old rats treated with EOD, the SIRT1 mRNA in the heart (p < 0.01), kidney (p < 0.001), and plasma SIRT1 protein were more than the corresponding AL groups (p < 0.001); however, the FW only increased this parameter in the kidney of these rats (p < 0.001) (Fig. 4).

Levels of SIRT1 mRNA in the
With AL feeding, the values of mRNA SIRT1 was lower in samples of middle-age and old rats, but were significant in the heart and kidney of old rats (p < 0.05) and the values of plasma SIRT1 protein decreased in both middle-aged and old animals (p < 0.01).
FW fasting did not cause significant differences in the expression of mRNA SIRT1 in heart and kidney of middle-aged and old than the young rats, but plasma levels of SIRT1 in middle-aged (p < 0.01) and old (p < 0.001) animals decreased versus the young group. EOD fasting significantly enhanced the expression of SIRT1 in the heart of old (p < 0.001) and in the kidney of middle (p < 0.001) and old (p < 0.001) animals and also in the plasma of middle (p < 0.01) and old rats (p < 0.001) (Fig. 5).
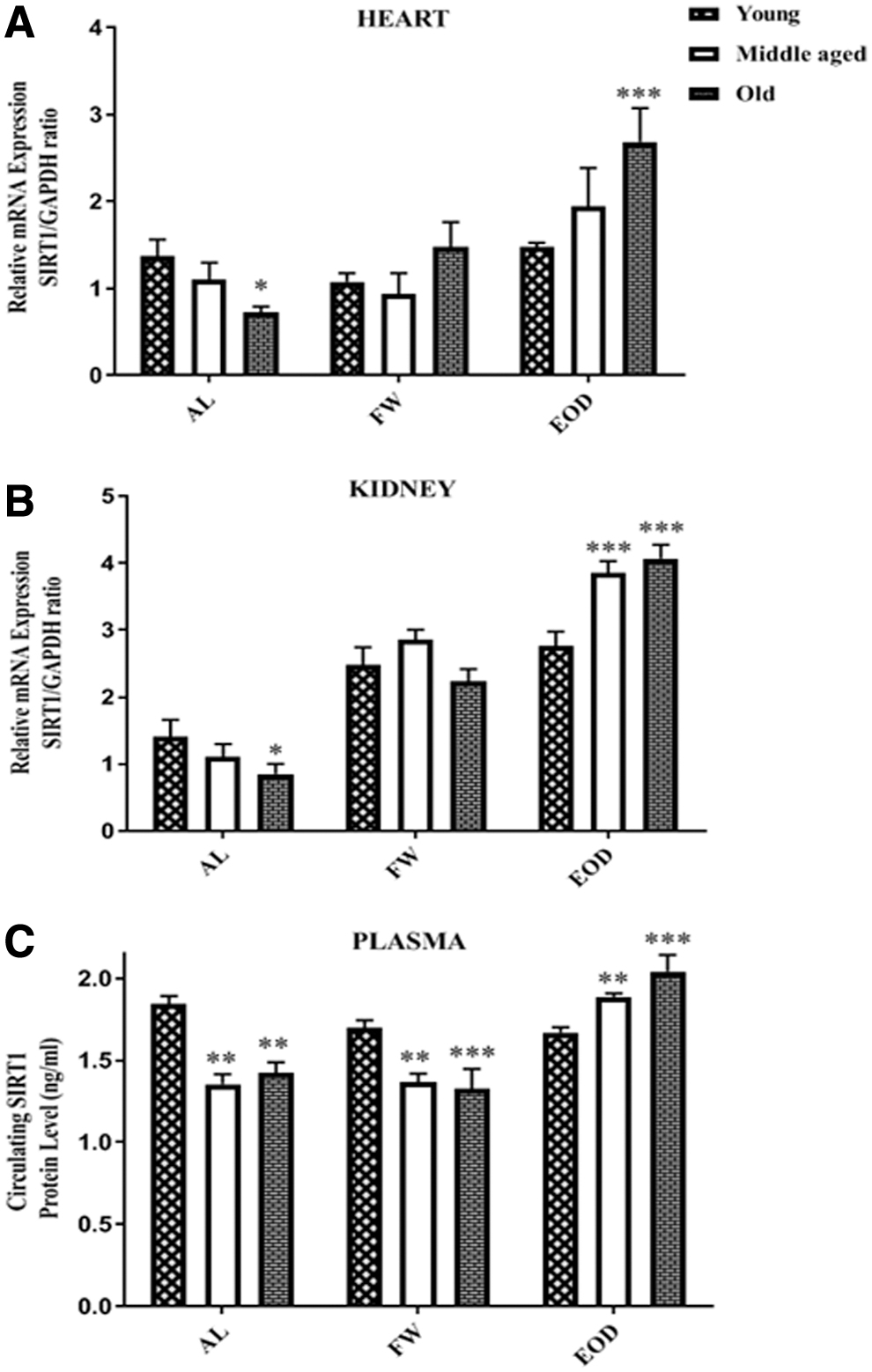
Levels of SIRT1 mRNA in the
The two-way ANOVA analysis showed a main effect of age (df = 2, F = 6.5, p < 0.01), diet (df = 2, F = 23.99, p < 0.001), and an age–diet interaction (df = 4, F = 4.00, p < 0.01) on SIRT3 mRNA in the kidney. There was also a main effect of age (df = 2, F = 29.70, p < 0.001), diet (df = 2, F = 24.93, p < 0.001), and age–diet interaction (df = 4, F = 9.5, p < 0.001) on SIRT3 protein of plasma. The analysis also indicated the main effect of age (df = 2, F = 22.97, p < 0.001), and age–diet interaction (df = 4, F = 8.07, p < 0.001) on SIRT3 mRNA of heart.
Under FW and EOD fasting, young animals showed lower level of SIRT3 mRNA expression in the heart (p < 0. 05 vs. AL regimen). SIRT3 mRNA in kidney and plasma SIRT3 enhanced significantly as a result of the FW treatment only (p < 0.01). In the presence of FW and EOD feeding, the middle-aged and old animals showed greater levels of SIRT3 expression in their samples when compared with corresponding AL regimen. The SIRT3 mRNA/protein was significantly higher in the kidney of middle-aged FW group (p < 0.01), in the plasma of middle-aged EOD group (p < 0.01), in the heart, kidney, and plasma of old FW group (p < 0.001), and also in the kidney (p < 0.001) and plasma (p < 0.01) of old EOD group versus the corresponding AL regimen (Fig. 6).
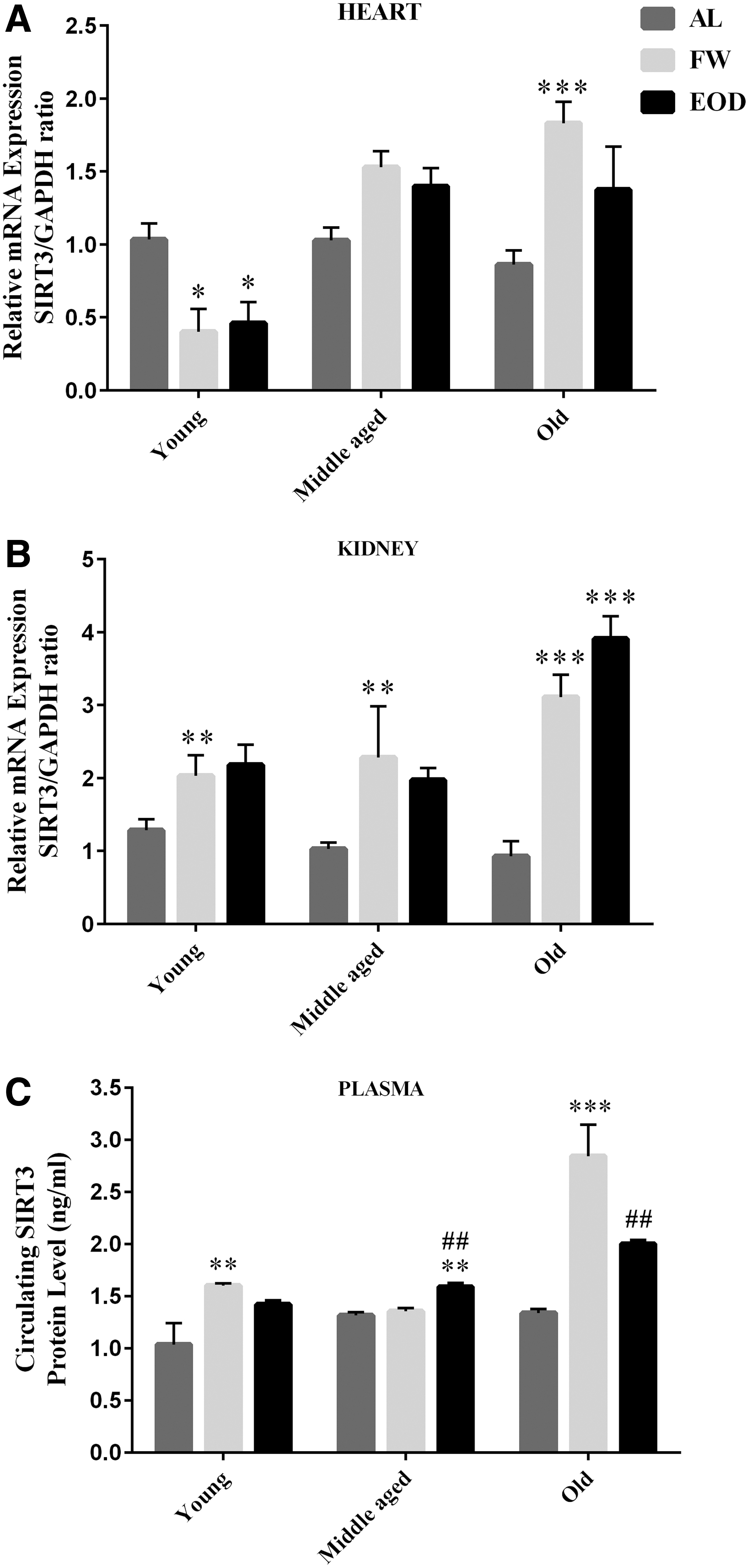
Levels of SIRT3 mRNA in the
The results showed that under AL feeding, the gene expression of SIRT3 was not affected by the aging process and that there was no significant difference among the groups in this regard.
In the presence of FW fasting, the SIRT3 mRNA was more expressed in the heart of middle-aged (p < 0.001 vs. young) and old rats (p < 0.001 vs. young). The SIRT3 plasma level was higher in the heart of old rats (p < 0.001 vs. young and middle aged). EOD fasting promoted the SIRT3 mRNA expression in the heart of middle-aged (p < 0.001) and old rats (p < 0.001) and in the kidney of old rats (p < 0.001) and also in the plasma SIRT3 of middle-aged (p < 0.05) and old (p < 0.001) rats when compared with young animals (Fig. 7).

Levels of SIRT3 mRNA in the
The two-way ANOVA analysis showed a main effect of age (df = 2, F = 5.19, p = 0.01), diet (df = 2, F = 8.09, p = 0.001), and age–diet interaction (df = 4, F = 9.62, p < 0.001) on plasma levels of α-klotho.
Plasma levels of α-klotho did not show significant changes in the FW group versus age-matched AL groups among the three age categories in our study. The value of this parameter was significantly greater in old animals who experienced EOD fasting than FW (p < 0.01) and AL (p < 0.001) age-matched groups (Fig. 8). The circulating levels of α-klotho was less with increasing age, so that it was lower in old animals in comparison with middle-aged (p < 0.001) and young animals (p < 0.001). FW fasting had no effect on blood α-klotho in middle-aged and old rats compared with the young group. However, EOD fasting increased the level of α-klotho in old rats compared with the regimen-matched middle-aged group (p < 0.05) (Fig. 8).
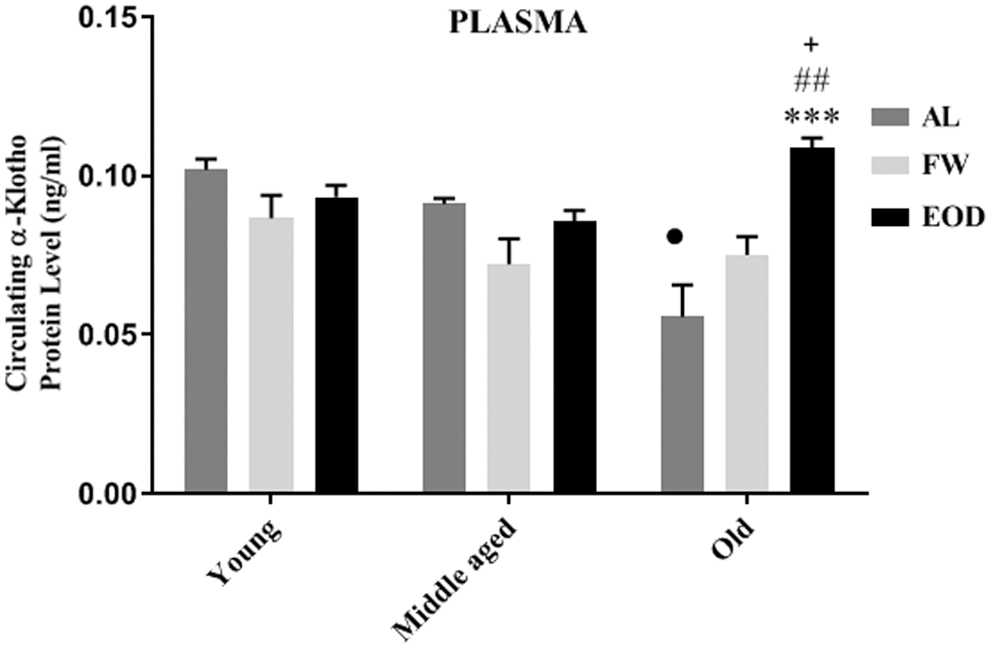
Effect of different patterns of fasting and age on blood α-klotho level in the rats blood, α-klotho protein level was measured using a rat α-Klotho Elisa Kit. Data are represented as mean ± SEM, analysis was done by one-way ANOVA, 5–6 animals per group, ***p < 0.001 versus age-matched AL group. ## p < 0.01 versus age-matched FW group. • p < 0.001 versus regimen-matched young and middle-aged groups. + p < 0.05 versus regimen-matched middle-aged group. AL, ad libitum regimen; FW, ad libitum and fasted 1 day per week; EOD, fed ad libitum and fasted every other day.
Discussion
In the present study, male rats were considered in three different age categories (young, middle-aged, and old) and they experienced two fasting regimens for 3 months. Then, their weight; HR; BP; the levels of SIRT1 and SIRT3 mRNA of heart and kidney; and SIRT1, SIRT3, and klotho proteins of plasma and blood insulin and glucose were compared with corresponding control groups.
The FW regimen had no significant effect on weight gain in middle-aged and old animals, but it reduced the slope of weight gain in young animals. The EOD regimen was associated with significant weight loss in all animal groups. Therefore, the negative effect of EOD on weight is more prominent than the effect of FW. This finding is in line with previous reports. Goodrick et al. demonstrated that body weight and growth rate can be reduced in rats under the EOD regimen from weaning to death and they concluded that dietary restriction increases longevity through retarded development. 26 However, in another study on mature rats of 10.5 and 18 months, Goodrick et al. showed that EOD can increase survival even beyond ages at which body weight growth usually ceases. 13 The results of other studies also showed that maintaining the EOD regimen for 3 months 9 and 6 months 27 is associated with low weight gain in young Sprague-Dawley rats. In the present and previous studies, 27 there are indications that animals with EOD tend to consume less food compared with rats fed on AL. These findings suggest that in long periods the animals are not able to double their dietary intake on the feast day to compensate the calorie of the fasting day, which results in a loss of body weight. However, some factors such as housing conditions, palatability or energy density of diet and genetics can be influenced on compensation.
One of the prominent physiological effects of calorie restriction in mammals is the decrease in insulin and glucose of plasma and the increase in insulin sensitivity. 28 The present study showed that both insulin and glucose were significantly higher in old rats with AL regimen than the young and middle-aged rats. Interestingly, both FW and EOD regimens eliminated this undesirable effect in old animals in a way that made the levels of glucose and insulin reach a similar level observed in young animals. However, these interventions had no significant effect on the levels of insulin and glucose in young and middle-aged groups when compared with their AL-matched groups. This finding suggests that similar to calorie restriction, fasting, especially EOD, has a beneficial modulatory effect on glucose metabolism. It is well known that in the long run, high levels of plasma glucose lead to high levels of nonenzymatic glycation, a form of protein damage and also cardiovascular and neurological disorders. In addition, prolonged exposure to glucose affects the glucose-regulated gene expression, including genes that control glucose metabolism itself. One of the outcomes of this phenomenon that is named glucose-induced transcriptional hysteresis is the induction of genes that increases glycolysis and inhibits gene expression of alternative metabolic pathways, such as the pentose pathway, beta oxidation, and the tricarboxylic acid (TCA) cycle. It is believed that following the cumulative exposure to glucose, similar metabolic shift occurs in aging. 29 Therefore, it can be proposed that fasting by correction of insulin responsiveness and glucose effectiveness can improve the glucose metabolism in the presence of low levels of glucose and insulin and induce its beneficial effect on lifespan.
Few studies have investigated the effect of EOD on the course of BP in physiological conditions and none has assessed the impact of FW on BP. Consistent with the current knowledge, 30 the present study revealed that old animals have significantly higher MAP, and systolic and diastolic pressures in comparison with middle-aged and young animals. On the other hand, FW and EOD regimens reduced MAP, and systolic and diastolic pressures of old animals to the extent that they reached the corresponding values of BP in young animals. In addition, EOD decreased the RPP as an index of myocardial oxygen consumption in middle-aged and old animals. This finding is similar with results of previous studies which showed that both CR and IF with similar magnitude can attenuate the BP of rats. 31 In the same context, studies on humans demonstrated the beneficial effect of fasting and CR on BP, regardless of whether the subjects are male or female, normal weight, overweight, obese, have diabetes type 2 or metabolic syndrome, or are hypertensive or prehypertensive. 32 Potential mechanism candidates for this beneficial effect are improving nitric oxide production and endothelium-dependent vasodilatation, 33 increasing levels of brain-derived neurotrophic factor and its positive effect on HR variability, 34 improvements in glucose and insulin regulation and reducing insulin resistance, and also the upregulation of SIRT1. 32 Improvements in insulin sensitivity through restoration of the activity of phosphatidylinositol-3 kinase 35 can attenuate BP. SIRT1 deacetylase can increase endothelial nitric oxide synthase (eNOS) deacetylation through lysines in the eNOS calmodulin-binding domain, thereby stimulating endothelial nitricoxide (NO) production 36 and reducing BP. In the present study, the increasing effect of EOD on SIRT1 mRNA levels of heart, kidney, and plasma SIRT1protein of old rats was confirmed that is in line with the effect of calorie restriction and can justify the positive effect of EOD fasting on BP in these animals.
Results showed a decreasing trend in SIRT1 and α-klotho but not in SIRT3 with the passing of time from youth to old age. However, fasting, especially EOD, induced positive effects on the expression of SIRT1, α-klotho, and even on SIRT3 particularly in old rats. In agreement with our findings, previous studies indicated that the level of SIRT1 in the heart is highest in the embryonic stage and then progressively reduces with aging. 37,38 An experimental study revealed that following the regimen of up to 24 hours fasting/24 hours, refed young mice showed enhanced levels of SIRT1 in the liver. 39
Previous findings have emphasized the requirement of SIRT1 as an important regulator of energy metabolism for the lifespan extension effects of calorie restriction. 40,41 Apparently, the effect of SIRT1 on glucose homeostasis is mediated through peroxisome proliferator-activated receptor gamma co-activator 1-alpha. 39 Therefore, similar to calorie restriction, it seems that EOD induces a part of its metabolic regulatory effect through improving the expression of SIRT1in old but not in young rats. In addition, our finding led to the speculation that fasting-induced SIRT1 overexpression can occur at certain ages with diverse patterns in heart, kidney, and plasma.
Endogenous SIRT3 is a soluble protein located in the mitochondrial matrix and is found in small amounts in the nucleolus. Involving in adaptive thermogenesis, cellular respiration and protection of cardiomyocytes from stress are some suggested roles for SIRT3. 42,43 The present study demonstrated that in the presence of FW and EOD regimens, the expression of SIRT3 in the heart was decreased in young rats, however, it showed an increasing trend in middle-aged and especially in old animals.
Therefore, similar to SIRT1, the effect of different fasting patterns on the expression of SIRT3 can vary in different ages and in different tissues. It has been reported that CR 44 and CR-mimetic agents, such as resveratrol, 45 upregulate SIRT3 mRNA expression in the adipose tissue of mice and in H295R cells, respectively. Therefore, similar to CR, fasting can modify the expression of SIRT3 in different tissues and thereby apply a part of its effects. The harmfulness of decreasing levels of SIRT3 in young hearts requires additional studies. However, it should be noted that the plasma level of SIRT3 showed an increasing trend in young animals that experienced FW and EOD when compared with AL. The increasing trend may compensate for the reduction of SIRT3 in the hearts of these animals.
The level of soluble α-klotho is inversely related to age and it decreases with aging. 46 In agreement with previous studies, our findings confirmed that the circulating level of α-klotho is significantly lower in old rats than in young rats under the AL regimen. Upregulation of the klotho gene in the kidney has been reported previously in CR adult male rats. 18 We showed that in comparison with the AL regimen, having 12 weeks of fasting can improve the protein levels of klotho in the plasma of old rats. This finding is consistent with the effect of fasting on the plasma level expression of SIRT1 in this study. The klotho gene was first introduced as an antiaging gene. Klotho is known to have a wide range of physiological effects, including antiaging, antioxidant, antiatherosclerosis, antiapoptosis, anti-inflammatory, tumor-suppressing properties, and also the maintenance of the endothelial integrity and function, NO production, and the maintenance of calcium homeostasis. 47 Klotho can reverse the aging process and provides cardiovascular/renal protection, possibly by inducing resistance to oxidative stress and protecting tissues from oxidative damage. 47,48
In klotho (+/−) mice, klotho deficiency promoted vascular dysfunction and hypertension that were mediated by mammalian target of rapamycin (mTOR) upregulation. 49 Also, klotho deficiency reduces SIRT1 activity in arterial endothelial and in smooth muscle cells, 50 thereby leading to arterial stiffening and hypertension. Therefore, klotho is likely to serve as an upstream regulator of vascular SIRT1 in the vasculature.
Conclusion
Taken together, this study clearly demonstrated that first, consistent with previous studies, 30,51 –53 the old animals showed higher levels of BP, insulin and glucose levels of blood, and lower levels of SIRT1 and klotho. Second, the effect of EOD fasting was prominent than FW fasting in improving the mentioned parameters. Third, age is a determining factor for rate of cellular and physiological responses to fasting, so that the old animals showed more desirable response to fasting. This effect may emanate from the fact that the aging process induces disturbance in molecular and physiological functions and some interventions such as fasting can become more effective. Fourth, similar to CR, a part of positive effect of fasting, especially EOD, apparently is mediated through improvement in the expression of SIRT1, SIRT3, and klotho proteins.
Footnotes
Acknowledgments
The authors are thankful to Dr. Alireza Sarkaki and Dr. Ali Khodadadi from Ahvaz University of Medical Sciences, Ahvaz, Iran, for their technical supports and their provision of animals, their house, and laboratory equipment for RT-PCR and other molecular tests.
Author Contributions
S.J. devised the main conceptual ideas and designed the study with F.B. F.B. performed the hemodynamic evaluation with the supervision of S.J. and M.B. F.B. purified all RNA and performed the real-time PCR and ELISA experiment with the supervision of M.R. T.D. helped in the performing of data analysis. S.J. supervised all parts of the study and edited the article with input from F.B.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by a grant (IR.KMU.REC.1395.313) from Kerman University of Medical Sciences, Kerman, Iran, and provided from the results of PhD thesis of Mrs. F.B.