
Abstract
Osteoporosis is a silent disease caused by low bone mineral density and is complicated by fractures. This study was designed to examine the differentiation of uterine stem cell-derived osteoprogenitor cells (UOPCs) both in vitro and in vivo, assessing their effectiveness in treating osteoporosis. CD271+/CD45− UOPCs were isolated from the endometrial tissue of inbred Balb/c mice through magnetic activated cell sorting. Stem cell differentiation assays were used for CD271+/CD45− UOPCs in vitro. In vivo, the UOPCs were implanted into mouse osteoporosis models through tail-vein injection for 8 weeks. Osteogenic differentiation was examined by X-rays and computed tomography (CT) scans. Enhanced green fluorescent protein (EGFP)-labeled UOPCs, obtained from C57BL/6-Tg (ACTb-EGFP) 1Osb/J mice, were used to assess cell survival in the osteoporosis model. The levels of osteogenic markers were assessed by enzyme-linked immunosorbent assay. In vitro, UOPCs were able to form into typical spheres and various differentiations. In vivo, implantation of UOPCs into osteoporosis model significantly increased bone mineral densities and bone microstructure parameters. The levels of a biochemical marker of bone metabolism, Semaphorin-3A, increased significantly. However, levels of receptor activator of nuclear factor kappa-B ligand decreased. Immunofluorescence staining of osteoporosis mice injected with green fluorescent protein+ UOPCs showed their survival for up to 7 days. In conclusion, stem cells with osteogenic differentiation potential can be isolated from uterine or endometrial tissue. These UOPCs can stably proliferate and differentiate in vitro or in vivo, which can inhibit bone resorption and osteoclast marker expression. In vivo, UOPCs significantly improved reduction in bone density caused by reduced estrogen levels. Such cell transplantation approach is potentially useful in the treatment of osteoporosis.
Introduction
Osteoporosis is the most prevalent bone disease in humans. It is well known that menopause is significantly associated with the risk of osteoporosis, 1,2 which is complicated by fractures that seriously endanger the life and health of elderly people. 3 Therefore, novel treatment methods that prevent osteoporosis after menopause are urgently needed. Existing treatments against osteoporosis mainly include administration of antiresorptive and anabolic agents. 4 However, no maintenance agents with long-term effectiveness are available. 5 Stem cell therapy is considered as a potential treatment for osteoporosis.
Stem cells can be derived from various tissues, including the uterus. 6 Stem cells exhibit the abilities for self-renewal and multipotent differentiation. 7 Several studies have demonstrated the importance of stem cells in treating diseases 8 such as myocardial infarction, ischemic kidney injury, and spinal cord injury, among others. 9 –11 Previous studies reported that transplanted mesenchymal stem cells (MSCs) could migrate into the host abnormal tissue and replace damaged and lost cells, which was investigated by nanoparticle imaging technology. 9,12,13 However, other studies reported that MSCs did not spontaneously implant and differentiate at the relevant sites after perfusion. 14
Human bone marrow-derived CD271+ MSCs show similar functional abilities as stem cells in phenotype expression, clonal growth, and multilineage differentiation. 15 A previous study revealed that CD271 inhibited osteogenic differentiation. 16 However, another study found that CD271+ cells could prevent osteoporosis. 17 It is generally known that one of the main causes osteoporosis in menopausal women is a declining estrogen secretion, which results in an increase in osteoclasts. 18,19 During the process of bone formation, the action of several molecules such as the bone morphogenetic protein (BMP) and semaphorin 3A (Sema3A) stimulate the activity of osteoblasts, 20,21 maintaining bone balance. 22 The potent antiosteoclastogenic function of Sema3A is regulated by receptor activator of nuclear factor kappa-B ligand (RANKL) signaling that suppresses osteoclast precursor cell migration. 20
Recently, stem cells have been detected in uterine or the endometrial layer and found to be similar to MSCs in terms of their phenotype, clonogenic growth, and multilineage differentiation. 23,24 However, whether uterine stem cell-derived osteoprogenitor cells (UOPCs) can differentiate into osteoblasts is unclear. In this study, we investigated whether CD271+/CD45− cells isolated from the mouse uterine could differentiate into osteoblasts and the microenvironment which could secrete estrogen or molecules to reduce the apoptosis of osteoblasts and decrease the risk of osteoporosis. The main aim of this study was to develop a novel strategy for treating osteoporosis using stem cells or the stem cell microenvironment.
Methods and Materials
Animals
All animal procedures were approved by the Animal Care Committee of the Shanxi Medical University Animal Institute. The following strains of mice were obtained from The Nanjing University animal laboratory: wild-type female Balb/c and female C57BL/6 mice (8-weeks old), which were used to establish the osteoporosis animal model into which uterine stem cell-derived osteoprogenitor cells (UOPCs) were implanted; and C57BL/6-Tg (ACTb-EGFP) 1Osb/J mice (8-weeks old), which express enhanced green fluorescent protein (EGFP) in all tissues except erythrocytes and hair.
Isolation of CD271+ cells by magnetic-activated cell sorting
Female Balb/c mice (8–10 weeks old) were anesthetized, intubated, and injected intraperitoneally with 10% chloral hydrate. Mice were continuously perfused with saline through the aorta at physiologic pressure until the liquid flowing from the right atrium was clear. Uteri were collected, minced, and digested twice with 0.25% trypsin, 2 mg/mL collagenase, and 0.01% DNase at 37°C for 1 hour. The uterine cells were then passed through a 70 μm filter. Red blood cells were removed from the blood samples using a 1% ammonium chloride lysis buffer. The magnetic activated cell sorting (MACS) analysis was performed by EasySep™ PE Positive Selection Kit (Catalog #17684, STEMCELL, Vancouver, British Columbia, Canada) and using the following antibodies: fluorescein isothiocyanate-conjugated anti-mouse CD271 (Santa Cruz Biotechnology, Dallas, TX) and phycoerythrin-conjugated anti-mouse CD45 (Abcam, Inc., Cambridge, MA). The CD271+/CD45− cell population (mean of 10,000 cells) was then analyzed for the purity by flow cytometer. The labeled cells were analyzed with an EPICS XLMCL flow cytometer equipped with Expo32 ADC Xa software (Beckman Coulter, Sacramento, CA). Overlay histograms were generated using FlowJo software.
Cell proliferation and differentiation
Individual CD271+/CD45− cell-derived blast colonies were carefully isolated from female Balb/c mice and transferred into differentiation media (Dulbecco's modified Eagle's medium-low glucose supplemented with 10% fetal bovine serum (FBS) and cell primary culture media containing 0.1 μmol/L dexamethasone, 10 mmol/L β-glycerophosphate, 2 mmol/L glutamine, 100 U/mL penicillin, and 100 g/mL streptomycin). Subcultured cells were seeded into 100-mm culture dishes (1 × 104 cells = dish) and cultured for the indicated periods in the differentiation medium. The cells were harvested by trypsinization and resuspended in the differentiation medium, and the cells were counted using a microscope counting chamber (hemocytometer). The cultures were maintained at 37°C, 5% CO2 in air, and more than or equal to 95% humidity.
BMP2 and β-catenin immunofluorescence staining
Immunofluorescence staining of BMP2 and β-catenin of in vitro differentiated UOPCs was carried out using the streptavidin-biotin peroxidase complex (SABC) method. After differentiation, the cultured cells were fixed and mounted on slides. The slides were deparaffinized and hydrated using a consecutive series of xylene and ethanol and treated with 9 mg/mL hydrogen peroxide to inactivate endogenous peroxidase activity. Slides were then incubated with 50 mg/mL bovine serum albumin (BSA) for 1 hour to block nonspecific staining. Subsequently, sections were incubated with a rabbit polyclonal antibodies to BMP2 (ab214821; Abcam, Inc.) or β-catenin (ab2365; Abcam, Inc.) at a dilution of 1:400 overnight at 4°C, and then incubated with goat anti-rabbit IgG (ab150078, ab150077; Abcam, Inc.), for 2 hours at room temperature. Immunoreactive signals were visualized by the addition of 3,3′-diaminobenzidine substrate (Sigma Chemical Co., St. Louis, MO). Negative control sections were incubated with normal rabbit serum omitting the primary antibody. To quantify the BMP2 or β-catenin staining, the slides were scored using a method similar to that described previously: −, no staining; +, weak staining; ++, moderate staining; and +++, strong staining. Relative immunostaining levels were evaluated at least three times.
UOPC sphere formation inhibition assay
UOPCs (1000 cells/well) were seeded in a 96-well black plate (Corning, New York, NY) and treated with fresh media. On the 3rd day, a image reader (Lion-heart, Biotek, Winooski, VT) was used to count the number of spheres (≥100 μm). Each measurement was repeated in a total of 14 wells, as shown in the results section.
In vitro differentiation potential assay of UOPC
The cells in each well were cultured in osteoprogenitor cell stimulation medium (Gibco; Thermo Fisher Scientific, Waltham, MA) for 2 weeks, after which bone nodules appeared around the stem cells. Aspirate medium from the cultures and rinse once with phosphate-buffered saline (PBS). Fix the cells for 10 minutes at room temperature using 100% methanol (−20°C). Following incubation, remove the fixative and rinse once in PBS. Remove the PBS and add a sufficient volume of freshly prepared Alizarin Red staining solution (Sigma Chemical Co.) to cover the dish. Incubate at 37°C for up to 20 minutes, protected from light, until the desired intensity of staining is reached. Aspirate and discard the stain, rinse with water, and air dry. At the end of the time course, all dishes were stained for Alizarin Red activity. Alizarin Red staining was visualized with an Olympus CKX 41 inverted light microscope.
The cells in each well were cultured in Adipose cell stimulation medium (Gibco; Thermo Fisher Scientific). When the cells reached about 80% confluence, 1 mM dexamethasone (Sigma-Aldrich, St. Louis, MO), 0.5 mM isobutyl-methyl-xanthine (Sigma-Aldrich), 10 μg/mL human recombinant insulin (Sigma-Aldrich), and 100 mM indomethacin were added. Lipid drops were visualized with Oil Red staining (Sigma-Aldrich) according to the manufacturer's instructions.
The cells in each well were cultured in chondrocyte differentiation stimulation medium (Gibco; Thermo Fisher Scientific). When the cells reached about 80% confluence,100 nM dexamethasone, 50 mM ascorbic acid, 40 mg/mL proline (Sigma-Aldrich), and 10 ng/mL transforming growth factor-β1 (PeproTech, Rocky Hill, NJ) were added. Chondrogenic differentiation was assessed that cells were incubated by 0.1% Alcian Blue solution prepared in 0.1 N HCl for 30 minutes according to the manufacturer's instructions of Chondrogenesis Differentiation Kit (Sigma-Aldrich).
Flow cytometry
One million cells from each sample were taken for antibody staining with cell surface markers or isotype-identical IgG (PE Rat Anti-Mouse CD90, Cat No. 553006; PE Rat Anti-Mouse CD105, Cat No. 562759; PE Rat Anti-Mouse CD73, Cat No. 550741; PE Rat Anti-Mouse CD45, Cat No. 561087; PE Rat Anti-Mouse CD34, Cat No. 551387; PE Rat Anti-Mouse CD14, Cat No. 553740; PE Mouse Anti-Human HLA-DR Cat No. 555559; all from BD Biosciences) for half an hour. The cells were then washed and resuspended in PBS supplemented with 2% FBS and 0.1% sodium azide. Cells were analyzed using a Becton Dickinson LSRII flow cytometer. The fluorescence intensity of 10,000 cells for each sample was quantified.
Surgical procedure
Before surgery, sodium penicillin (22,000 U/kg) and gentamicin (6.6 mg/kg) were administered to Balb/c mice or C57BL/6 mice intravenously. Under general anesthesia, the hair along both sides of the spine was shaved. A scalpel was used to create a 1 cm incision on each side of the spine to expose the ovaries, which were then ligated with an absorbable suture, after which the ovariectomy surgery was performed (OVX, n = 28, including 7 C57BL/6 mice). Sham-operated mice were used as the control group (n = 14, including 7 C57BL/6 mice). Wounds were tied with absorbable gut sutures. After this, a small amount of erythromycin ointment was applied to the sutures and the wounds. Six weeks after surgery, radiology was used to verify the establishment of osteoporosis.
Radiology examination
Balb/c mice and C57BL/6 mice were anesthetized with 10% chloral hydrate, stably placed on the scanning stage, and then scanned with a Hologic Discovery system using dual-energy X-ray absorptiometry (Bedford, MA). The image analysis software allowed for a calculation of the overall bone density values and the bone mineral content. Regional High-Resolution Version 4.76 (Hologic) was used to analyze the data.
Following anesthesia with 10% chloral hydrate and fixation to the scanning stage, mice were also scanned by micro-CT (eXplore Locus SP; GE Healthcare, WI). A bone thickness of 1.55 mm was selected as the region of interest (ROI) for the femur cancellous bone distal growth plate. For the cortical bone analysis, a bone thickness of 3 mm was selected as the ROI. A 8.0 × 8.0 × 8.0 μm voxel was used for three-dimensional (3D) reconstruction, and the image information was extracted by computer to automatically generate the domain values. The Microview 2.0+ ABA software was used for quantitative analysis of the reconstructed micro-CT images.
Diffusion chamber implantation
CD271+/CD45− UOPCs of Balb/c mice were isolated, trypsinized, and resuspended at a definitive density. Under sterile conditions, the cells were then loaded into a Theracyte immunoisolation device (Design Engineering Group, Baxter Healthcare, Round Lake, IL) with the use of a gas-tight syringe and blunt-tipped stainless-steel needles. The loading port was then sealed with sterile silicone adhesive (Dow Corning, Midland, MI). The 1 × 105 cells/injection, resuspended in 50 μL of medium, was then implanted into the tail vein of Balb/c model mice. Cells in the continuously injected UOPC group (CIUCs, n = 7) were implanted immediately after OVX surgery, once a week up to a total of 8 weeks. The tonifying UOPCs group (TUCs, n = 7) were implanted 8 weeks after OVX surgery, also the 1 × 105 cells/injection, resuspended in 50 μL of medium, once a week. The sham group (n = 7) and the OVX group (n = 7) were implanted using the same volume of PBS used in the CIUC group once a week to a total of 8 weeks. Femur sections were analyzed by light microscopy, and images were digitized by using a color digital video camera (Olympus, Center Valley, PA) adapted to a BX71 microscope (Olympus). Morphometric analyses were performed 8 weeks after treatment by using software Image Pro Plus version 7.0 (Media Cybernetics). The number of trabecula was determined by counting 10 fields (100 × ) per femur in hematoxylin and eosin-stained sections. All the analyses were blinded.
Isolation of CD271+/CD45−/EGFP+ UOPCs by MACS
Female C57BL/6-Tg (ACTb-EGFP) 1Osb/J mice (8–10 weeks old) were anesthetized, intubated, and injected intraperitoneally with 10% chloral hydrate. Uteri were collected, minced, and digested, the same as Balb/c mice. The uterine cells were passed through 70 μm filter. Then, red blood cells were removed. The MACS analysis was performed by EasySep PE Positive Selection Kit (Catalog #17684, STEMCELL) and using the following antibodies: fluorescein isothiocyanate-conjugated anti-mouse CD271 (Santa Cruz Biotechnology) and phycoerythrin-conjugated anti-mouse CD45 (Abcam, Inc.). The CD271+/CD45−/EGFP+ UOPCs population (mean of 10,000 cells) was then analyzed the purity by flow cytometer.
Confocal immunofluorescence analyses
In addition, after 1 week, the C57BL/6 mice of osteoporosis model were implanted with EGFP+ UOPCs CD271+/CD45− that were from uterus of C57BL/6-Tg (ACTb-EGFP) 1Osb/J mice by MACS, which were 1 × 105 cells/injection, resuspended in 50 μL of medium. They were anesthetized with 10% chloral hydrate and sacrificed to analyze the trace of the target cells.
Femoral heads obtained from osteoporosis or sham of C57BL/6 mice were preserved in paraffin embedded slides, and 10 μm sections were obtained using a Thermo HM325 microtome (Thermo Fisher Scientific Ltd., Newton Drive, Carlsbad, CA). To evaluate the presence of CD271+/CD45−/EGFP+ UOPCs in the femoral head, the tissue sections were blocked in 3% H2O2 for 30 minutes at room temperature, and then blocked for 30 minutes in 4% normal goat serum in 0.1 M PBS with 1% polyvinylpyrrolidone, 1% BSA, and 0.3% Triton X-100. The primary antibody- anti-p75 nerve growth factor receptor antibody (CD271) (ab52987; Abcam, Inc.) were incubated overnight at 4°C, washed with PBS, and then incubated with the secondary antibody- goat anti-rabbit IgG (ab150078, ab150077; Abcam, Inc.) at room temperature for 20 minutes. Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI) using VECTASHIELD HardSet mounting medium and DAPI H-1500 (Vector Laboratories, Burlingame, CA). Sections were then analyzed using confocal microscope (FV1200; Olympus). Images were then captured with FluoView 10 (Olympus) using Adobe Illustrator C53 software.
Enzyme-linked immunosorbent assays for Sema-3A and RANKL
Blood was collected from mice by the eyeball puncture method. The blood was centrifuged at 17,968 g to 20 minutes to obtain serum. Enzyme-linked immunosorbent assay (ELISA) kits (Biomotic, Inc., Kansas City, MO) were used to measure the Sema-3A and RANKL levels according to the manufacturer's recommendation.
Statistics
Data are presented as mean ± standard deviation. Statistical analyses were performed using GraphPad Prism 5 (GraphPad, La Jolla, CA). A two-tailed Student's t-test was used for two-group comparisons. All other comparisons among three or more groups were analyzed using a one-way analysis of variance followed by a Tukey post hoc test. Differences were considered statistically significant at p < 0.05.
Results
Radiological analysis of the mouse model of osteoporosis induced by bilateral OVX
The OVX of female mice used to establish the osteoporosis model is a simple and safe surgical approach (Fig. 1A–C). A reduction in bone mineral density (BMD) has been reported 6 weeks after OVX in the lumbar spine and can be detected 4 weeks after OVX in the distal radius. We found that the spinal BMDs and bone cortex thickness of the control group (n = 7) were significantly higher than the OVX group (n = 7) (p < 0.05) (Fig. 1D). Six weeks after OVX, we used both X-ray photography and CT-imaging (Fig. 1E–H) to examine the femur and spine. The presence of osteophytes was also found in femurs in the OVX mice compared with control mice (Fig. 1I, K); Vertebral bone cortex damage was also evident in the OVX mice compared with the control mice (Fig. 1J, L), which indicated that the model of osteoporosis was successfully established.
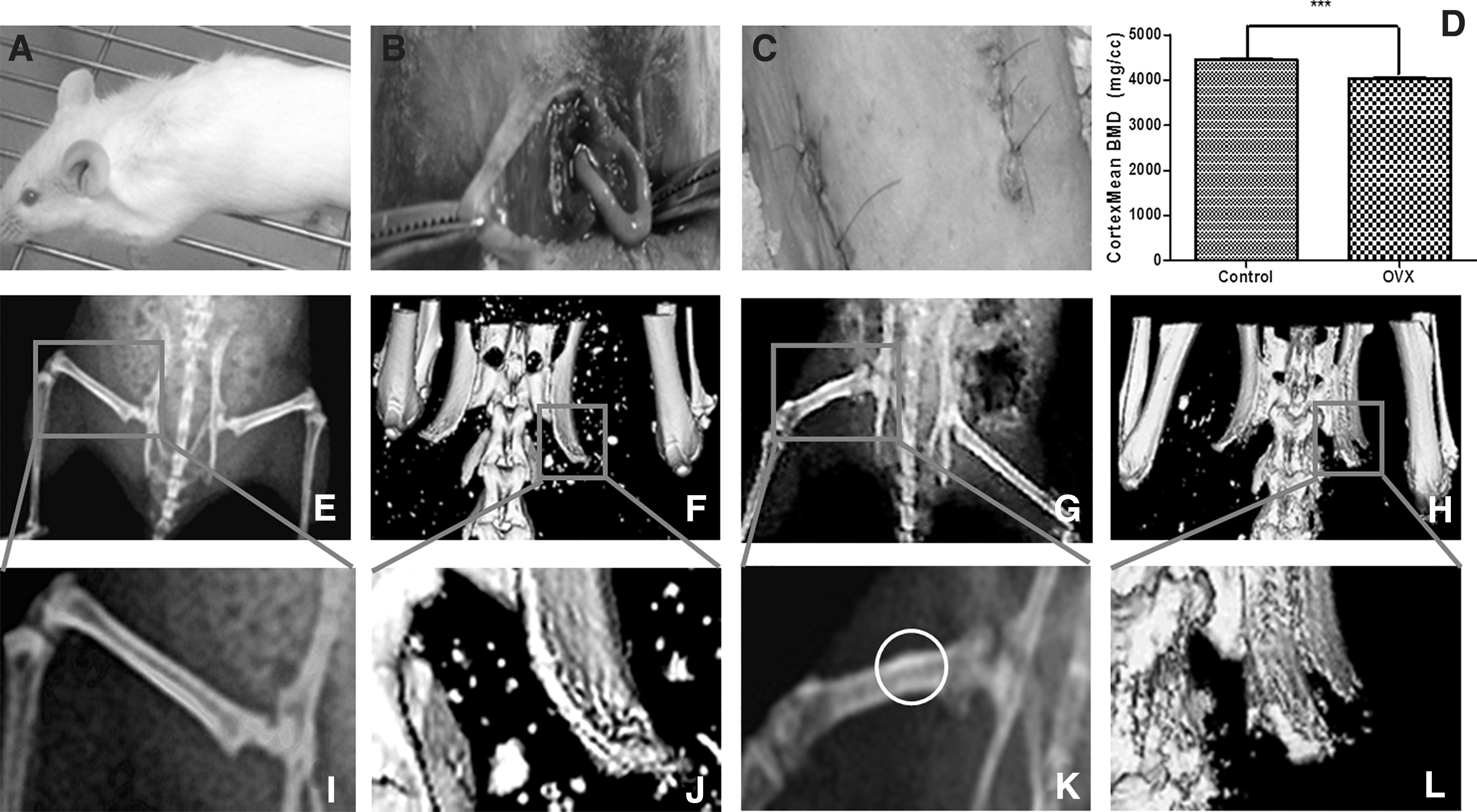
Radiology-imaging detects the animal model of osteoporosis induced by bilateral OVX in mice.
UOPCs can be used to treat osteoporosis
After established the osteoporosis model, we explored the therapeutic effect of UOPCs against osteoporosis. The uterus contains contractile myometrial tissue and extensive vascularized endometrial tissue, which are periodically shed and regenerated. CD271+/CD45− UOPCs were obtained from the uterus of Balb/c mice using MACS, and were of 1 × 105 cells/injection/mouse then implanted into OVX female Balb/c mice (8–12 weeks old), once a week. After 8 weeks, spinal and trabeculae BMDs were assessed by X-ray photography and CT imaging. The number of lumbar spine trabeculae was assessed using a 3D reconstruction imaging technology and hematoxylin-eosin staining. We found that, compared to OVX mice, the femur cortex mean BMD of mice with CIUCs was increased significantly (p < 0.01). The femur cortex mean BMD of sham group was significantly higher than OVX groups (p < 0.01) and higher than other groups. The femur cortex mean BMD of TUCs group was significantly lower (p < 0.01) than CIUCs group. Osteophytes were found on the femoral backbone of mice in the OVX group and the TUC group. Similar data were observed for the trabeculae mean BMDs (Fig. 2B1, D1).
The 3D reconstruction images allow for a calculation of the bone volume (BV) to tissue volume (TV) ratio. The BV/TV ratio in the sham group was the highest of all the experimental groups (Fig. 2A2). In the OVX group (Fig. 2B2), the BV/TV ratio was significantly lower than sham group (p < 0.01) and CIUCs group (p < 0.01) (Fig. 2C2) (Fig. 2E). The CIUCs group BV/TV ratio was higher than the ratio in the TUCs group. The ranks for the BS/BV ratios were the inverse of the BV/TV ratios for the four groups (Fig. 2F). The BS/BV ratio in the sham group was the lowest of all the groups (Fig. 2G) (p < 0.05), whereas the OVX group ratio was significantly higher than sham group (p < 0.01) and CIUCs group (p < 0.05). The ratio in TUCs group was higher than CIUCs group.

UOPCs have preventive functions to osteoporosis. Detected femur BMD of
The data from the number of bone trabeculae evaluated using the 3D reconstruction imaging technology and by femoral head hematoxylin and eosin staining were consistent. The number of bone trabeculae in the sham (Fig. 2A3, A4) was higher than in the CIUC group (Fig. 2C3, C4). It was also higher than in the TUCs group (Fig. 2D3, D4), and also higher than the OVX group (Fig. 2B3, B4). These were all significantly different (p < 0.05).
In vitro differentiation of UOPCs
CD45− UOPCs were obtained from the uterus by MACS, cultured to form a clonogenic blast colony (Fig. 3A), and then selected for CD271+ also by MACS (Fig. 3B). From the 26 blast colonies examined, 7% were CD45− and 0.06% were CD271+/CD45−. These UOPCs were addressed by extensive flow cytometry analysis, purity of sorted CD271+/CD45− cells from uterus were 93.1% (Supplementary Fig. S1A).
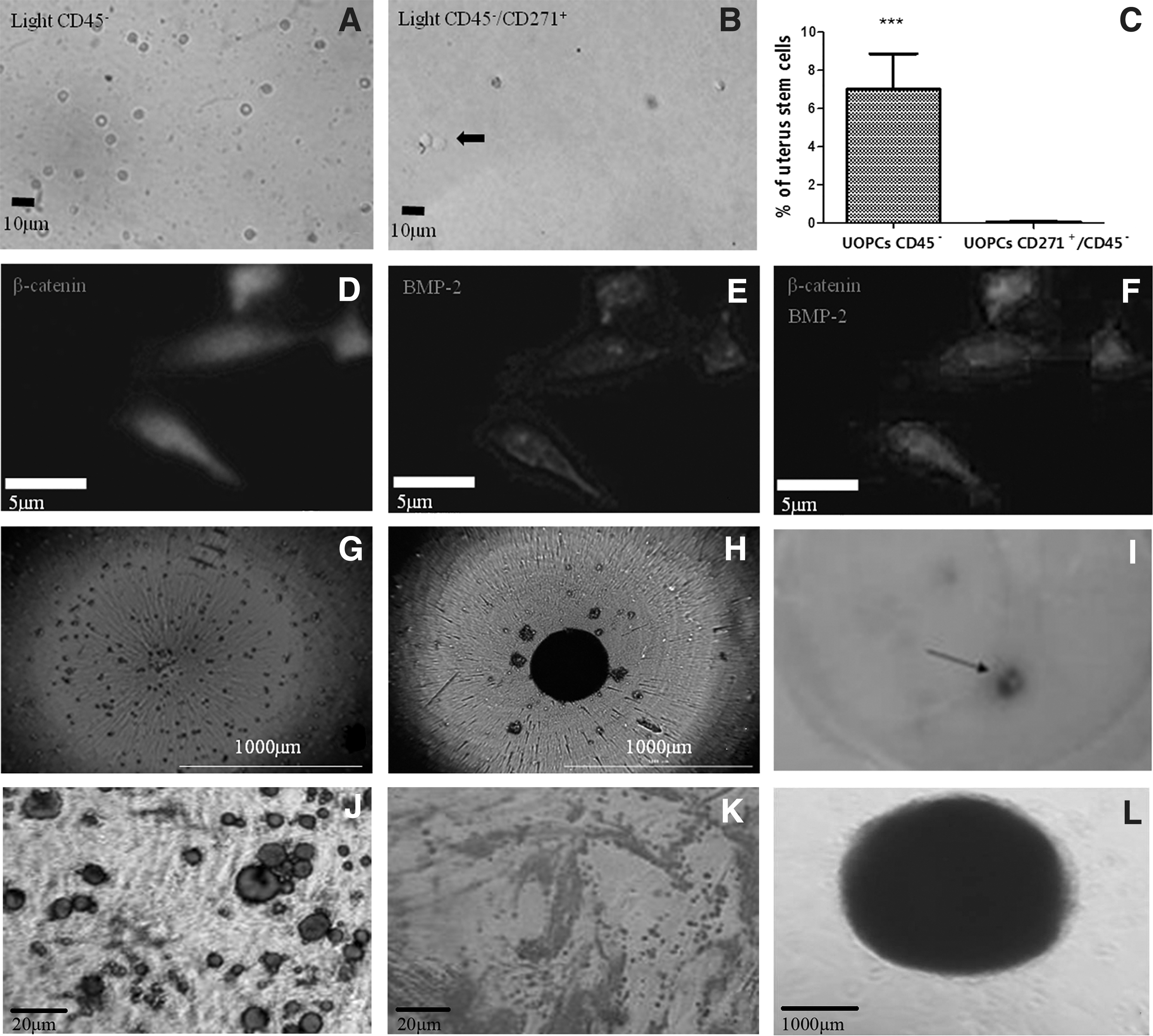
UOPCs CD271+/CD45− have multidifferentiation potential in vitro.
The adherent cell population expressed bone morphogenetic protein 2 (BMP-2) and β-catenin by immunofluorescence staining (Fig. 3F). The presence of strong positive expression of β-catenin (Fig. 3D) and BMP-2 (Fig. 3E), which are important pathways of osteogenic differentiation, confirmed that the CD271+/CD45− UOPCs can differentiate into osteoblasts.
Unsorted uterine or endometrial cells were not formed typical spheres in 3 days (Fig. 3G). However, The CD271+/CD45− UOPCs formed typical spheres in the 3rd day (Fig. 3H), suggesting that stem-like cells were enriched in the spheres. The osteogenic differentiation was confirmed by Alizarin Red staining (Fig. 3I, J). The adipogenic differentiation was confirmed by Oil Red staining (Fig. 3K). The chondrogenic differentiation was confirmed by Alcian Blue staining (Fig. 3L).
Characteristics of UOPCs in vitro
After, the CD271+/CD45− UOPCs were cultured and confirmed with notable self-renewal and multilineage differentiation capacities. UOPCs were addressed by extensive flow cytometry analysis to characterize stromal cells at the protein level. The CD271+/CD45− UOPCs expressed highly positive for CD90 (98.01%), CD105 (98.40%), and CD73 (99.12%), whereas CD45, CD34, CD14, and HLA-DR were not positively detected by fluorescence-activated cell sorting analysis (Fig. 4).

MSC surface marker assay in UOPCs. MSC, mesenchymal stem cell.
Osteoprogenitor cells derived from the uterus have the ability for short-term self-renewal
To test CD271+/CD45− UOPCs have the characteristics of MSCs, we also examined osteogenic differentiation in vivo to examine the retention and renewal properties of these cells. EGFP+ UOPCs CD271+/CD45− (1 × 105) (Supplementary Fig. S1B) were extracted from C57BL/6-Tg (ACTb-EGFP) 1Osb/J mice uteri (Fig. 5A) and transplanted into wild-type C57BL/6 mice, which had been verified by the establishment of osteoporosis before (Fig. 5B). These EGFP+ UOPCs were allowed to differentiate in the wild-type C57BL/6 osteoporosis mice for 7 days (Fig. 5C), and could be clearly seen in the femoral head of the recipients on the 8th day (Fig. 5D).
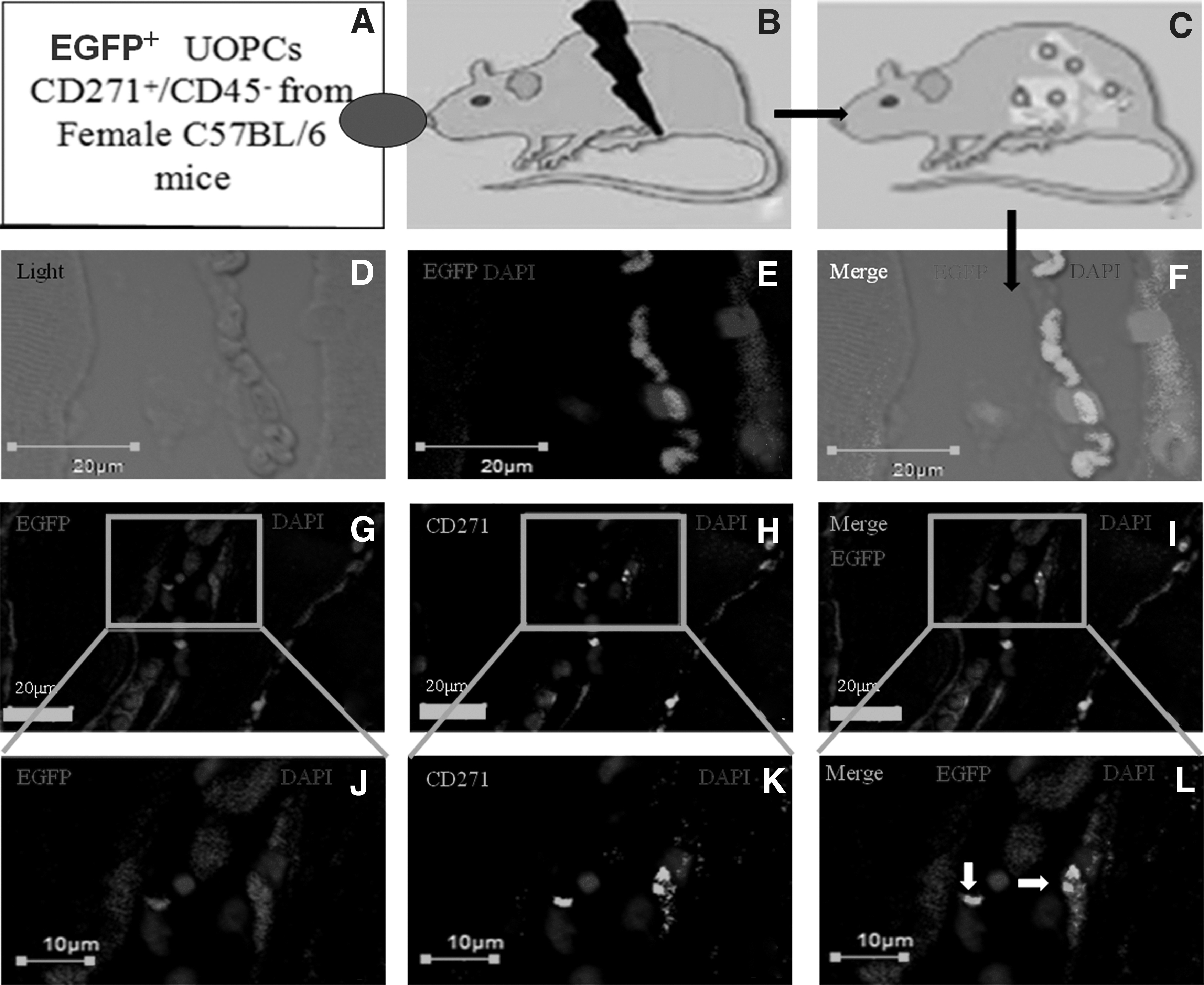
The UOPCs CD271+/CD45− have the abilities of short-term self-renewal in vivo.
The transplanted EGFP+ UOPCs improved osteogenic function in the osteoporosis mice. By the 8th day, EGFP+ UOPCs could be detected in the femoral head regions (Fig. 5E). The femoral head was removed from the recipient C57BL/6 mice and was found to contain both wild-type and EGFP+ cells capable of osteogenic formation (Fig. 5F). The cells from the EGFP+ mouse uterus could be identified by double-staining for CD271 and EGFP+ (Fig. 5I). The femoral head slices indicated EGFP+ cells (Fig. 5G) that also expressed the CD271+ at 7 days (Fig. 5H). These results showed that uterine CD271+/CD45− cells retain their capacity for osteoblast differentiation and extensive engraftment in vivo.
UOPCs affect Sema3A and RANKL levels
Sema3A exerts an osteoprotective effect by both inhibiting bone resorption and promoting bone formation. 20 Sema3A levels in sera from the CIUCs group were significantly higher than in the TUCs group (p < 0.005) and were significantly higher than the OVX group (p < 0.005). The levels of Sem3A in TUCs group were significantly higher than the levels in the OVX group (p < 0.005). The Sema3A levels in the sham group were significantly higher than OVX group (p < 0.005) (Fig. 6A).
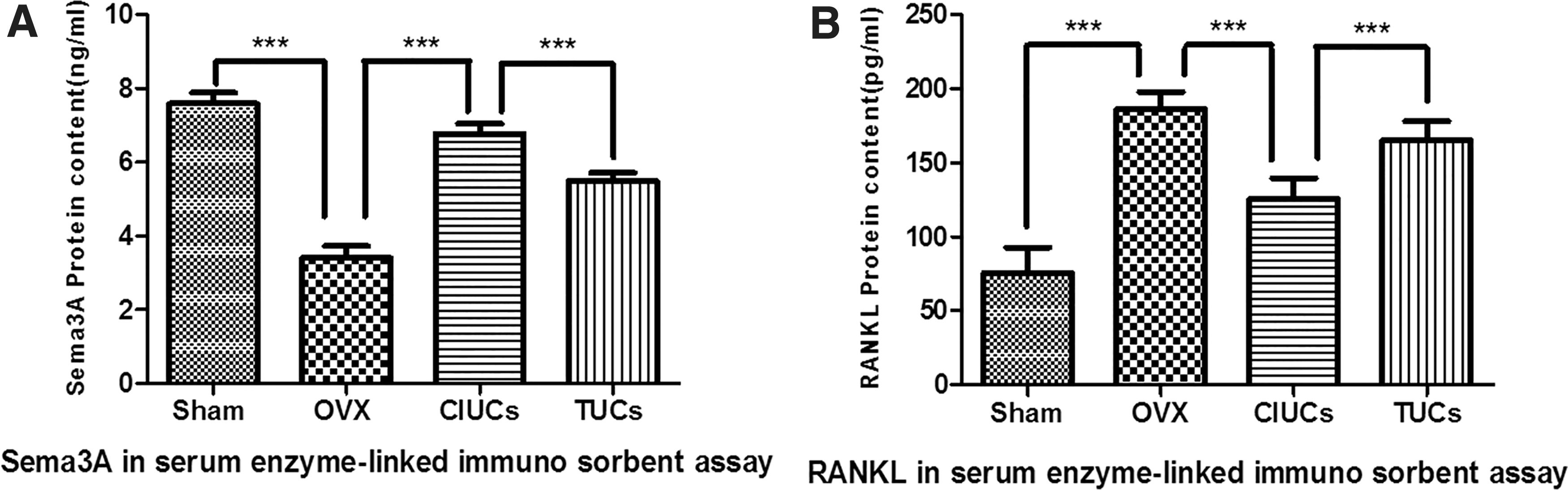
UOPCs CD271+/CD45− promoted the secretion of Sema3A protein in vivo, which have the effect on pathway for osteogenesis.
RANKL is an important factor in osteoclast differentiation and formation. The RANKL levels in the sham group were significantly less than CIUCs group levels (p < 0.005), the TUCs group levels (p < 0.005), and the OVX group levels (p < 0.005). The RANKL levels were highest in the OVX compared to sham group and CIUCs group (p < 0.005). The RANKL levels in TUCs group were also significantly higher than the CIUCs group (p < 0.005) (Fig. 6B). These results indicated that CD271+/CD45− UOPCs can be derived from the uterine or endometrium. Therefore, we can better predict that the UOPCs implantation could be one treating or preventing method against osteoporosis (Supplementary Fig. S2)
Discussion
In this study, we isolated CD271+/CD45− UOPCs from endometrial tissue by MACS and then explored the potential use of these osteoprogenitor cells to treat osteoporosis. We used a mouse model of osteoporosis by OVX and injected the UOPCs through the tail vein. We assessed osteogenesis by examining the cortex and trabecula in mice as well as assessing the levels of serum osteogenic biomarkers. The data confirmed that CD271+/CD45− UOPC treatment can reduce the damage caused by osteoporosis.
The animal model and UOPC injection
To create the osteoporosis model, we performed an OVX whereby the bilateral ovaries were surgically removed, as previously described. 25 This model has the advantage that it does not use glucocorticoids which often require individual doses. 26 After OVX, although there is currently no clear standard diagnosis for osteoporosis. 27 The symptoms of osteoporosis were assessed after 4 weeks at least by both X-ray photography and CT scan. We used 4–6 weeks to allow osteoporosis to fully develop.
To avoid any immune tolerance issues, the UOPCs were derived from the same batch of Balb/c mice. 28 Our results indicated that both the bone cortex BMD and trabecular BMD in the CIUCs group were higher than in the TUCs group. This suggests that CD271+/CD45− UOPC transplantation should be conducted before trabecular damage has occurred. Once the symptoms of osteoporosis are established firmly, treatments can have very limited effects. 29
From the X-ray and micro-CT scans, we concluded the following: (1) the lack of estrogen leads to a decline in both femur cortex BMD and trabeculae BMD; (2) destruction of the trabecular structure causes osteoporosis; and (3) the implantation of UOPCs can inhibit bone reabsorption and increase bone trabecular thickness. This result therefore suggests a new cell-based treatment for menopausal women suffering from osteoporosis. 30 Currently, we cannot explain the precise mechanism by which the addition of osteoblasts inhibit osteoclasts. Furthermore, it is also uncertain if the lack of estrogen plays a role in inducing bone resorption, since bone can be affected by other factors, such as IL-6, 31 which should be clarified in the future by advanced imaging or gene level. 13,32
Differentiation and homing of UOPCs
The CD271+/CD45− UOPCs were shown to be able to differentiate into osteoblasts by immunofluorescence staining for BMP-2 and β-catenin, stemness assays by formed sphere assays, Alizarin Red staining, Oil Red staining, and Alcian Blue staining. We did not assess whether Wnt/β-catenin signaling or other pathways were important in stromal cell differentiation. 33 The current study were able to identify stem cells in uterus tissue that had pluripotent differentiation capabilities such as MSCs, and analyzed the stromal cell characteristics at the protein level by flow cytometry, 34 this would support the theory that the uterus can act as a stem cell pool. 35,36 Although UOPCs appeared to be able to treat osteoporosis, it is not clear whether this was a direct effect in bone or an indirect effect on other pathways to provide appropriate proliferation conditions for osteoblasts. 37
We observed that CD271+/CD45− UOPCs have the ability for short-term self-renewal. We also addressed if these cells were capable of long-term retention and self-renewal. An obstacle to detect the accuracy of treatment was whether these cells of interest were distinguished from host cell or organism. 38 We used CD271+/CD45− UOPCs with both the expressions of the EGFP marker gene (green) and CD271+ (red) to establish that these stem cells were UOPCs and not derived from the marrow. In addition, if these uterine cells were not derived from stem cells, the EGFP+ cells would likely be destroyed by phagocytosis for they were without self-renewal ability, 39,40 which proved CD271+/CD45− UOPCs is one of the stem cells. We have also tried to determine the kinetics of cell survival over the period of 1 week. We used imaging which avoids the sampling biases and errors, which occur when groups of animals are euthanized at different time points.
Biochemical markers of osteogenesis
In our study, the data showed that the CD271+/CD45− UOPCs implanted into mice via the tail-vein implantation not only increased Sema3A levels in serum but also decreased RANKL levels which would be expected to inhibit bone resorption, which results were consistent with other studies. 41,42 These results suggested that this treatment for osteoporosis could be a pathway that increases Sema3A levels and inhibit the RANKL pathway. A recent study has shown that stem cell transplantation can not only enhance BMP levels in the lumbar spine and the femur but also increase the Sema3A levels in serum. 20 However, in current study, although Sema3A could play a reparative role in damaged bone caused by osteoporosis, after osteoporosis had completely formed, the serum Sema3A levels would not obviously rise after this treatment. Regardless, the data suggest that there may be a RANKL/Sema3A pathway which could participate in the coregulation of bone metabolism.
RANKL is an osteoclast marker. Previous studies have shown similar effects on RANKL levels using the implantation of MSCs derived from bone marrow mononuclear cells. 43,44 There were different results that treatment of animal models of osteoporosis established by OVX or injection of IL-6 or IL-17 antibodies also decreases RANKL levels, 31,45 which were not clear as to whether RANKL levels were mediated by osteoblasts, or a key pathway in osteoporosis, 46 which should be explored in the future work.
UOPC injection against corticosteroids-induced osteoporosis
Recent research found that stem cells in the endometrial layer were detected from endometrial tissue that obtained from women by biopsy (curettage), which can be obtained without anesthesia, invasive, and painful interventions. 47 However, whether CD271+/CD45− UOPCs that were obtained by curettage can be used in the treatment to osteoporosis in human needs future study. Severe acute respiratory syndrome (SARS) had a negative impact on health in China in the 21st century. 48 To protect against SARS, it was initially suggested that prolonged, high-dose, corticosteroids be used. 49 However, long-term treatment with corticosteroids has many side effects, mainly including osteoporosis. 50 In the current study, we did not investigate any systemic side effects in mice. The reasons could be that the stem cells improve cellular microenvironments, which could play a key role against bone resorption, activating both proliferation and differentiation of host cells. Whether UOPC injection could be more secure and efficient treatment against osteoporosis in human need more experiments.
Conclusions
Our study has proved that inducible osteoprogenitor stem cells can be derived from the uterine or endometrium and appear effective in a murine model of osteoporosis, perhaps by increasing Sem3A levels and decreasing RANKL levels. CD271+/CD45− UOPCs implantation could therefore be useful for treating or preventing osteoporosis, particularly in postmenopausal patients.
Footnotes
Acknowledgments
The authors' contributions were as follows—M.H.: guided the study; J.X.: designed, supervised, and guided the study; Z.W.: conducted the experiments and wrote the article; D.W.: conducted the experiments; Y.L.: statistical analysis; and all authors: contributed to the design of the statistical analysis, interpreted data, critiqued and revised the article, and read and approved the final article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by National Natural Science Foundation Projects (grant no. 81671462). This study is also supported by the Special Public Welfare Industry Research of National Health and Family Planning Commission of China (grant no. 201402010); This study is supported by the applied basic research project of Shanxi province (grant no. 201601D011086).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.