
Abstract
Aging is strongly correlated with several noncommunicable disorders such as diabetes, obesity, cardiovascular disease, and neurodegenerative conditions. Glucosamine (2-amino-2-deoxy-
Introduction
Aging is characterized by a continuous decline in all physiological functions. 1 The process of aging involves altered cellular homeostasis, elevated oxidative stress, and accelerated cellular senescence. These changes make the body susceptible to several age-related diseases. 2 There is also widespread decline and deregulation of membrane-mediated signal transduction in different cells and organs during aging. 3
Calorie restriction (CR) is one of the most effective interventions for improving lifespan and slowing the rate of aging in lower species. 4 CR intervention involves a reduction in food consumption (up to 30%–50%) without inducing malnutrition. 5 Despite its established efficacy in lower animals, the implementation of CR as a regular lifestyle in humans is still a subject of investigation. 6 Caloric restriction mimetics (CRMs) that include synthetic or natural substances imitating the effects of CR are being studied for their antiaging benefit. Similar to CR, CRMs also activate stress response pathways, generate CR-like longevity outcomes, and reduce the likelihood of aging-induced diseases. 7,8 CRMs are reported to modulate several cell signaling pathways and have been classified as sirtuin stimulating compounds, AMP-activated protein kinase (AMPK) activators, mammalian target of rapamycin (mTOR) antagonists, and autophagy inducers in various animal models. 9
Glucosamine (2-amino-2-deoxy-
GlcN has been shown to induce insulin resistance and mimic hyperglycemic condition in adipocytes by rapidly increasing flux through the HBP, resulting in increased O-linked glycosylation of regulatory proteins. 15
When administered orally, it has also been documented to influence the metabolism of carbohydrates and reduce body fat in rodents. 16 and lead to improved oxidative stress tolerance, accompanied by AMPK activation. 17 The compound has also been documented to induce autophagy in mammalian cells through an mTOR-independent signaling pathway. 18
In 2007, Ristow and colleagues first reported mitohormetic extension of lifespan in Caenorhabditis elegans exposed to 2-deoxy-
In recent years, a rat model of accelerated senescence has been developed, based upon treatment with
Erythrocytes provide a good model system to study age-dependent biomarkers. 24 In the circulation, erythrocytes are in constant exposure of both endogenous and exogenous sources of ROS that can impair its functions. In the microcirculation, where the erythrocytes are in proximity to the vasculature, ROS released from neutrophils, macrophages, and endothelial cells are taken up by erythrocytes. 25,26 Erythrocytes are especially vulnerable to oxidative damage due to the heavy content of polyunsaturated fatty acids in their membranes and a large amount of cellular oxygen and hemoglobin. 27
Several redox parameters in erythrocytes and plasma have been established as reliable biomarkers of aging, these have been frequently used to study age-dependent alterations. 28,29
In this study, an attempt has been made to explore the potential role of GlcN as a possible CRM, based on its property of a potent glycolytic inhibitor, in an accelerated senescence model of rat. The proposed study is expected to provide an understanding of what is the mechanism of action of glycolytic inhibitors when this class of compounds exerts their effects as CRM.
Materials and Methods
Chemicals and reagents
2,4-Dinitrophenylhydrazine (DNPH), dithiobis nitro benzoic acid (DTNB), glucosamine hydrochloride was purchased from TCI Chemicals (India) Pvt. Ltd. All other chemicals were of analytical grade procured from Merck, Germany, and SRL, India.
Animal model and the study protocol
Male Wistar rats were used in the analysis and were collected from the CSIR-Indian Institute of Toxicology Science, India. Rats were independently housed in temperature-controlled environments (temperature 20°C–25°C and relative humidity 55% ± 15%) with a 12-hour light/dark period and ample exposure to drinking water and nutrient-rich pellets. Twenty-four rats were used for experimentation. For each study, rats in four groups (n = 6) were randomized to the following groups:
Group I young control: rats (4 months old, b.w. = 150 ± 20 g) received orally 0.9% saline (as a vehicle) for 12 weeks.
Young glucosamine hydrochloride II (Y-GlcN): young rats (4 months old, b.w. = 150 ± 20 g) treated orally with (300 mg/kg b.w.,) once daily for 12 weeks.
Accelerated senescence model III (aged): rats (4 months old, b.w. = 150 ± 20 g) treated with
Senescent rats + GlcN IV (aged GlcN): rats (4 months old, b.w. = 150 ± 20 g) were administered with
All the protocols and procedures were followed as mandated by the ethical committee of the University of Allahabad, Allahabad, India. Approval # IAEC/AU/2017(1)/011.
After completion of the treatment procedure, rats were anesthetized with intraperitoneal administration of pentobarbital 50 mg/kg body weight.
Isolation of red blood cells
After the treatment schedule, blood was obtained through a cardiac puncture in dry tubes and heparinized syringes. The whole blood was centrifuged (800 g) at room temperature to separate plasma from Packed RBCs (PRBCs). After removal of the plasma (immediately kept at −80°C), the concentrated leukocyte band (the buffy coat) including the top 15% of concentrated red blood cells was discarded. This was to minimize the leukocyte contamination from PRBCs. The erythrocytes thus obtained were washed three times with ice-cold phosphate-buffered saline (PBS, pH 7.4).
The protein content was measured using bovine serum albumin as a standard according to the method of Lowry. 30
Measurement of ROS in erythrocyte by using 2,7-dichlorodihydrofluorescein diacetate dye
ROS generation in the erythrocyte of all experimental groups was measured using 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA) dye following our earlier protocol. 31 The generation of ROS in treated groups is expressed as a percentage change with respect to control.
Measuring the antioxidant ability in terms of FRAP
Ferric reducing ability of plasma (FRAP) is based on the reduction of ferroin analogue by antioxidants in the acidic medium by following the developed Benzie and Strain protocol. 32
Reduced glutathione level in erythrocyte
Erythrocyte reduced glutathione (GSH) was measured using 5.5-dithiobis-(2-nitrobenzoate) (DTNB) at 412 nm. 33 In short, 1 mL of PRBCs (10%; w/v) and 1 mL of 0.15 M KCl were combined and deproteinized by introducing 3 mL of metaphosphoric acid solution. Tubes were centrifuged at 3000 g for 20 minutes and 0.5 mL of supernatant was applied to 2 mL of 0.3 M of Na2HPO4 2H2O and 0.5 mL of DTNB. GSH amounts are reported in μM/mg protein.
Estimation of PMRS activity
The activity of the erythrocyte plasma membrane redox system (PMRS) was determined by the reduction of ferricyanide as mentioned previously. 34 The findings are represented as micromol ferrocyanide per milliliter PRBCs per 30 min.
Estimation of superoxide dismutase
Superoxide dismutase (SOD) was measured following the reported method 35 using nicotinamide adenine dinucleotide hydrogen as a substrate and the activity is expressed as units per milligram protein.
Estimation of catalase activity
Catalase (CAT) activity of whole experimental groups in plasma was measured by the H2O2 degradation assay following the reported protocol 36 and data are expressed as μM H2O2 decomposed per minute per milligram protein.
Estimation of advanced glycation end products
The determination of advanced glycation end products (AGEs) was based on spectrofluorimetric detection following the protocol previously published. 20 In brief, serum was diluted 1:50 with PBS and the fluorescence intensity was recorded at a maximum emission of 440 nm at 350 nm. Fluorescence intensity is expressed as an arbitrary units per gram protein in arbitrary units.
Determination of plasma sialic acid
Plasma sialic acid levels were calculated using a procedure stated earlier by us. 37 Ten times diluted plasma was treated with 0.04 M of periodic acid and incubated in an ice bath for 30 minutes. Then 1.25 mL of resorcinol working solution (5 mL of 6.0% resorcinol solution and 0.125 mL of 0.1 M copper sulfate solution delivered to a final volume of 50 mL with 10 M HCl) was added and incubated in a water bath for 5 minutes. After that, 3.25 mL of n-butanol was added and centrifuged for 5 minutes. The resultant supernatant was read at 625 nm. Plasma levels of sialic acid are reported in micromolar.
Statistical analysis
Statistical analysis was performed using the software PRISM version 5.01. All the values are expressed as mean ± standard deviation for six animals (experiments performed in duplicates) in each group. The statistical significance of differences was calculated by one-way analysis of variance followed by post hoc Bonferroni test, p < 0.05 was considered significant.
Results
Spectrofluorometric data show a significant increase in erythrocyte intracellular ROS (p < 0.05) in GlcN-treated young and senescent rats (
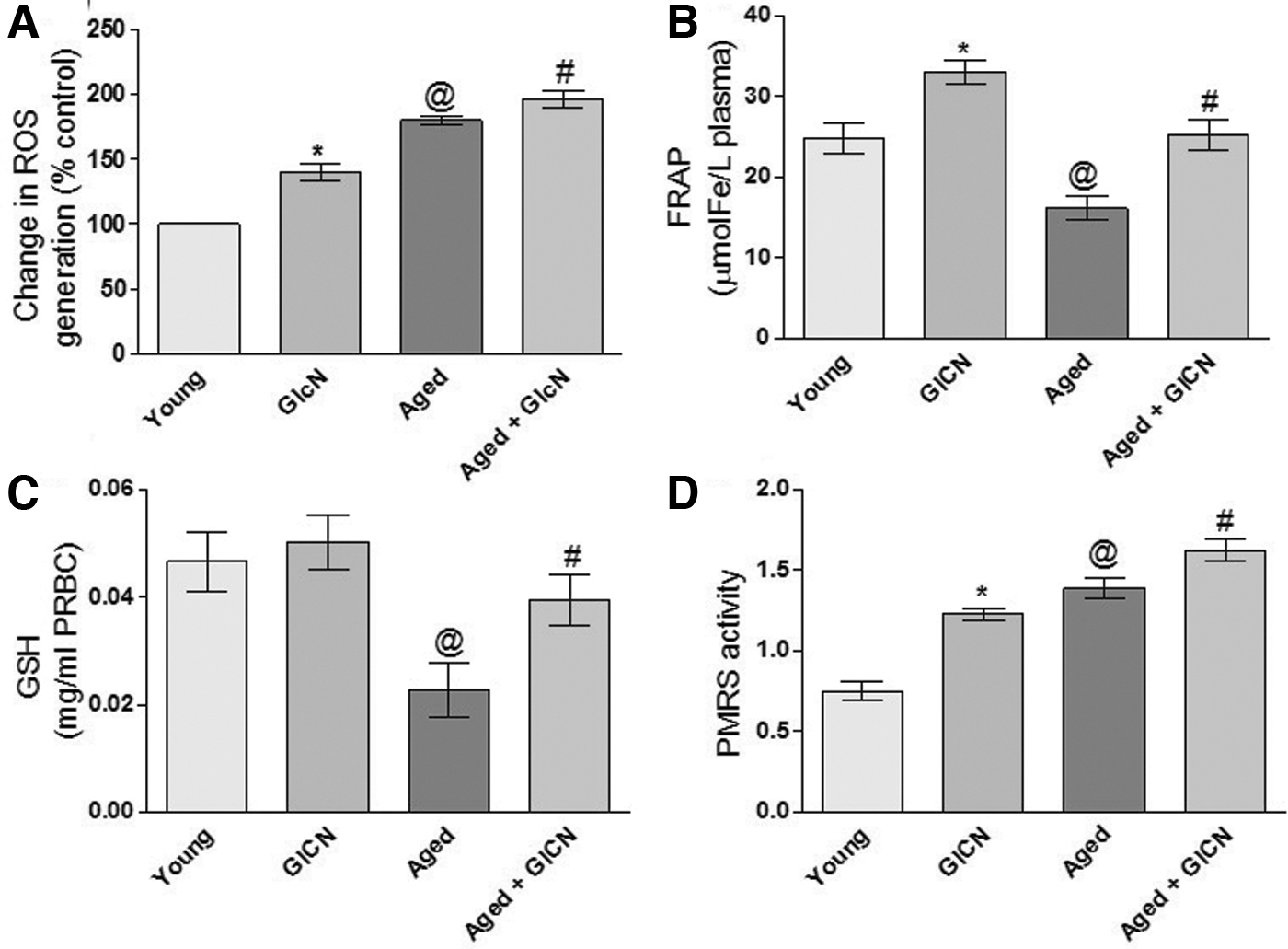
Figure 1B shows the ferric reducing ability of plasma (FRAP). There was a significant increase (p < 0.05) in plasma antioxidant capacity in GlcN-treated young and aged rats with respect to their controls. In control rats, the value of FRAP decreases with age. When compared with GlcN-treated young and aged rats, the FRAP values decreased significantly (p < 0.05).
Intracellular GSH levels are depicted in Figure 1C, there was a nonsignificant change in the erythrocyte intracellular levels of GSH in GlcN-treated young and aged rats groups with respect to their control groups. Significant (p < 0.05) decreases have been found in young and aged GlcN-treated rats and their respective controls.
Erythrocyte PMRS activity is shown in Figure 1D. There was significantly increased PMRS activity (p < 0.05) in young GlcN-treated rats compared with the control group. Similarly, higher values of PMRS activity were confirmed in GlcN-treated aged rats groups. Significant (p < 0.05) increase is found in GlcN-treated young and aged rats, with respect to their respective controls.
Figure 2A and B shows the SOD and CAT activity. There is a significant increase (p < 0.05) in antioxidant enzymes SOD and CAT in GlcN-treated young rats with respect to the young control. However, there is a significant decrease in SOD activity in aged treated rats when compared with young treated rats. Whereas CAT activity is significantly (p < 0.05) increased in both young and GlcN-treated aged groups with respect to their control.

Figure 3A shows the formation of AGEs that is decreased nonsignificantly in young rats after the treatment with GlcN. In aged rats, there is a significant (p < 0.05) decrease in AGEs after GlcN supplementation compared with aged rats. There is a nonsignificant increase in AGEs in GlcN-treated young and aged rats. However, there is a significant (p < 0.05) change in young and aged controls.
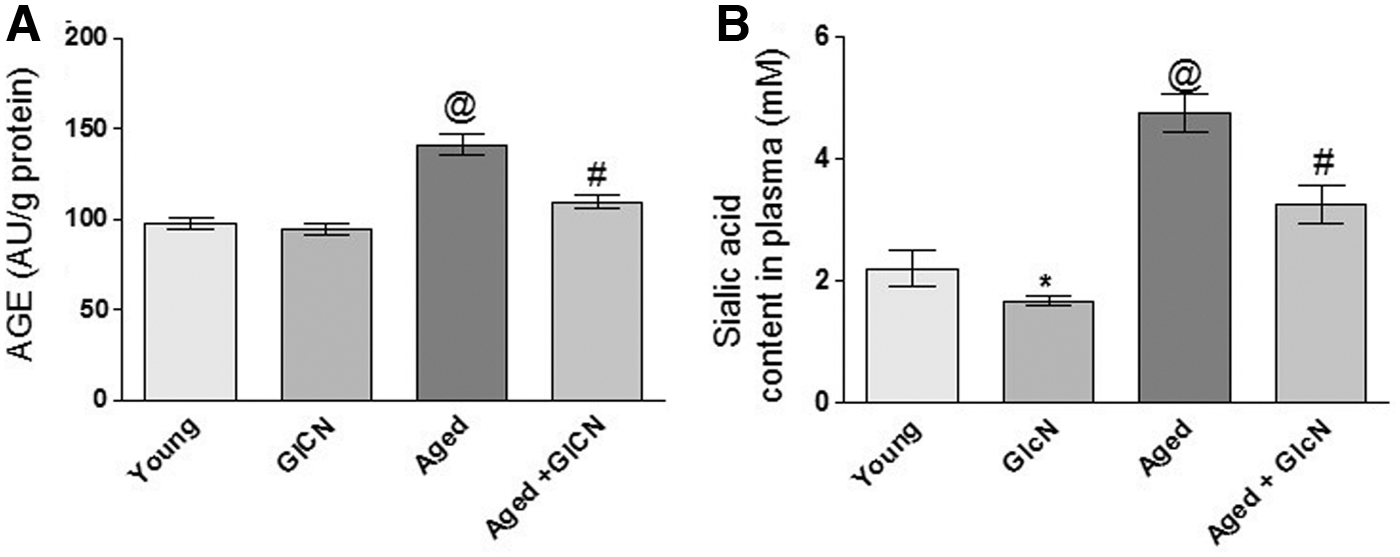
Plasma sialic acid is represented in Figure 3B, as a function of age, sialic acid in plasma increases. However, our result shows a significant decrease (p < 0.05) in plasma sialic acid in GlcN-treated senescent rats as compared with their controls. A similar result has been followed in GlcN-treated young rats. Sialic acid was found to be increased significantly (p < 0.05) in senescent GlcN-treated rats when compared with young treated rats.
Discussion
Although CR is documented to prolong lifespan in many model organisms, it is difficult to implement long-term CR in humans due to serious side effects and ethical considerations. It is due to these concerns that CRMs are being intensively investigated. In this study, our selection of GlcN was based on the assumption that a reduction in intracellular glucose availability could mimic the effects of CR. 12 Initial reports show that GlcN provides protection against age-associated damages in erythrocytes, increases mitochondrial biogenesis, lowers blood glucose, and increases amino acid catabolism and membrane polarity as reported for other glycolytic inhibitors such as 2-deoxyglucose. 18 Similar to resveratrol, another CRM that potentially slows down aging and certainly slows down age-related diseases, GlcN, is known to activate sirtuins as well as induce mitohormetic reactions. 38,39
Our finding of low FRAP values in old rats corroborates previously published results, 20 a higher FRAP value in senescent rats after GlcN treatment indicates there is an improvement in redox homeostasis and this effect may contribute to the CRM effect. However, a higher ROS, post-GlcN treatment in both young and senescent rats, appears to be at variance with elevated FRAP. Higher ROS would be predicted to lower the antioxidant level. A plausible reason for the paradoxical higher FRAP despite increased ROS could be a hormetic effect induced by ROS, resulting in an augmented antioxidant resistance. 40 GlcN treatment upregulates mitochondrial respiration and short-term induction in mitochondrial ROS production. This initial ephemeral ROS induction encourages mitochondrial biogenesis and ameliorates antioxidant response resulting in GlcN-mediated lifespan extension in the long term. 18 SOD and CAT are the most important antioxidant enzymes. Our analysis showed an improved antioxidant mechanism evaluated in terms of SOD and CAT activity. Significantly increased SOD and CAT activity have also been reported in mitochondria during CR. 41 Thus, we propose that the restriction of glucose metabolism contributes to the ROS protection through a secondary rise in SOD and CAT activity after a primary rise in ROS levels.
The plasma redox membrane system (PMRS) is a defense mechanism employed by cells to mitigate oxidative stress and has been hypothesized to provide defense against oxidative stress during aging. 34,42,43 A higher erythrocyte PMRS has been hypothesized to be a contributing factor for a longer lifespan. 44 Exogenous antioxidants including tea catechins and resveratrol have been reported to display their antioxidant effects by providing electrons to the PMRS to activate its function 45 ; we hypothesize that GlcN may activate PMRS by a similar mechanism. GlcN-treated rats display increased PMRS activity in both young and senescence rats, this observation explains that GlcN may adopt the same pathways as other CRM compounds such as resveratrol, which activate PMRS during aging. 1,46
GSH levels decline during the progression of age and age-related diseases, which also contribute to age-dependent impaired redox balance. 47,48 GlcN-treated rats do not show significant changes in GSH levels, this may be attributable to the presence of disulfide reductase, thioredoxin, which converts the oxidized (sulfonic) form of cysteine back into thiolate anions. The membrane of erythrocyte has a net negative surface charge contributed by sialic acid. Proof of this decreased erythrocyte surface charged in aged rats suggests an elevated incidence of other pathological disorders. 49 Serum AGE is an important substitute biomarker for oxidative stress. 50 A high level of blood glucose is a key factor that promotes aging due to the generation of AGEs. 51 Our study shows an elevated level of AGEs in senescent rats, which corroborates earlier research. 20 There was a substantial decrease in the amount of AGEs after treatment with glucosamine in old rats, a possible explanation could be that the glucosamine amine group competes with the sugar-reducing proteins for nonenzymatic reactions, thus inhibiting the development of AGEs. 52 Growing data suggest that ROS not only cause oxidative stress but rather may also function as signaling molecules that promote health by preventing/delaying several chronic diseases. The effects are manifested as an increase in lifespan. 53 Although high levels of ROS are usually expected to cause cellular harm and to facilitate aging, low levels of ROS have been shown to enhance systemic defense mechanisms by triggering an adaptive response. This phenomenon has been referred to as mitochondrial hormesis or mitohormesis. 54
A nutrient-restricted diet induces moderate stress. Following the concept of mitohormesis, GlcN promotes the increased formation of mitochondrial ROS. Higher ROS, as seen in GlcN-treated rats, suggest stress in the cells under conditions of low glucose supply. Concomitantly with higher ROS, certain antioxidant biomarkers display elevated status, suggesting a potential antiaging activity of GlcN, substantiating its CRM function. Our findings thus show that GlcN acts through an elevated ROS-induced mitohormetic cycle to protect the cells from age-dependent oxidative shifts. Hence, our study supports a growing concept that mildly elevated ROS protect cells and tissues through secondary response pathways.
Footnotes
Ethical Statement
All the protocols and procedures were followed as mandated by the ethical committee of the University of Allahabad, Allahabad, India.
Acknowledgment
Financial funding from Department of Biotechnology, Govt. of India under the RRFSP is gratefully acknowledged.
Authors' Contributions
S.I.R. conceived and designed research. R.K. conducted experiments, analyzed data, and wrote the article, K.S. conducted experiments and wrote the article. All authors read and approved the article and all data were generated in-house and that no paper mill was used.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by a research grant to S.I.R. from the SERB, Department of Science and Technology, Govt. of India (EMR/2016/006470), the Department of Biochemistry is supported by an FIST grant of DST-SERB, and UGC-SAP Grant.