
Abstract
Spilanthes acmella Murr., a well-known Thai traditional medicine, has been used for treatment of toothache, rheumatism, and fever. Diverse pharmacological activities of S. acmella Murr. have been reported. In this study, antioxidative and neuroprotective effects of S. acmella Murr. extracts as well as bioactive scopoletin, vanillic acid, and trans-ferulic acid found in the aerial parts of this plant species have been described. Protective effect of S. acmella Murr. extracts and bioactive compounds on dexamethasone-induced neuronal cell death was investigated. Different plant crude ethyl acetate (EtOAc) and methanol (MeOH) extracts including pure compounds of S. acmella Murr. were evaluated in human neuroblastoma SH-SY5Y cells. Cytotoxic effects were performed by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Mechanisms involved in the antioxidant effects of S. acmella Murr. regarding the activation of antioxidant marker proteins such as superoxide dismutase 2 (SOD2) and sirtuin 3 (SIRT3) were determined using 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) assay, Western blot analysis, and immunocytochemistry. Dexamethasone significantly caused the decrease of SH-SY5Y cell viability. Conversely, the increases in reactive oxygen species (ROS), autophagy, and apoptosis were observed in dexamethasone-treated cells. S. acmella Murr. MeOH and EtOAc extracts, as well as the bioactive compounds, reversed the toxic effect of dexamethasone by increasing the cell viability, SIRT3 protein expression but reducing the ROS, autophagy, and apoptosis. This study demonstrated that S. acmella Murr. may exert its protective effects against ROS through SOD2 and SIRT3 signaling pathways in dexamethasone-induced neurotoxicity. S. acmella Murr. may be a candidate therapy for neuroprotection.
Introduction
Depression is a devastating disorder that gives a negative impact on structural and functional plasticities of the brain development. Previous studies indicated that patients with depression show increased activity of the hypothalamus pituitary adrenal axis, and excessively high level of glucocorticoid. 1 Glucocorticoid acts on glucocorticoid receptor and mineralocorticoid receptor. Under physiological condition, the glucocorticoid is released into the blood by the adrenal gland, and exerts anti-inflammatory and immunosuppressive properties. Stress and elevation of glucocorticoid have been shown to enhance neurodegeneration. 2
Dexamethasone is a synthetic glucocorticoid, which blocks the migration of the human neuroblastoma SK-N-SH 3 and also induces cell cycle arrest, inhibits normal cell activity, and causes nervous system dysfunction resulting in the activation of neuronal cell death in the cerebellum, cortex, hippocampus, hypothalamus, and amygdala. 4,5 The neuronal cell death is usually characterized by cell morphological and biochemical changes, including cell volume diminution, mitochondrial transmembrane potential collapse, phosphatidylserine evagination, chromatin condensation, DNA fragmentation, apoptotic body formation, and apoptotic cell death. 6,7 Recent evidence demonstrates that high dose of dexamethasone induced neurotoxicity via impairment of mitochondrial dynamics in SH-SY5Y cells. 8
Spilanthes acmella Murr., known in Thai as Phak-Krad-Hauwaen, is a herbaceous plant of the Compositae family that is widely grown in the tropic and subtropic areas. S. acmella Murr. has long been used as an ingredient in local dishes, and a traditional herb for treatment of severe toothache, stomatitis, rheumatism, malaria, and fever. 9 A variety of biological effects have been reported such as antioxidant, anti-inflammatory, antipyretic, antidiabetic, anticancer, and vasorelaxant activities. 10,11 Previous investigations demonstrated that aerial parts of S. acmella Murr. exerted hepatoprotective activity by reversing the elevation in the level of alkaline phosphatase, aspartate transaminase, and alanine aminotransferase. 12 In addition, aerial parts of S. acmella Murr. ethyl acetate (EtOAc) and methanol (MeOH) extracts showed strong and potent antioxidant activity. 10 It has been reported to decrease inflammation and oxidative stress in central nervous system. 13 However, ethnopharmacological studies regarding antioxidant effects of S. acmella Murr. remain unclear.
Phytochemical studies revealed that the genus contains alkaloids, tannins, flavonoids, alkylamides, spilanthol, undeca-2E-ene-8,10-diynoic acid isobutylamide, 14 α- and β-amyrin esters, germacrene-D, β-sitosterol, 15 myrecene, stigmasterol, 16 myricyl alcohol, phenolics, coumarin, triterpenoids, limonene, (Z)-β-ocimene, 17 β-caryophyllene, 18 3-acetylaleuritolic acid, vanillic acid, β-sitostenone, scopoletin, trans-ferulic acid, and trans-isoferulic acid. 10 S. acmella Murr. extracts have been reported for health promoting effects and well-being with minimal undesirable side effects. 19 Modern pharmacological research showed that the extracts of S. acmella Murr. could protect neuronal cell death against pesticide-induced neurotoxicity. 20 In addition, scopoletin, vanillic acid, and trans-ferulic acid containing compound of S. acmella Murr. were reported to display neuroprotective effects. 21 –23
The signaling pathway that regulates cellular stress, superoxide dismutase 2 (SOD2), and sirtuin 3 (SIRT3) has been implicated in neuronal death. SIRT3 is a nicotinamide adenine dinucleotide (NAD+)-dependent deacetylase, which maintains mitochondrial homeostasis 24 through Oxidative phosphorylation (OXPHOS) regulation and mitochondrial oxidative stress. 25 It is localized in mitochondrial inner membrane and matrix, and in nucleus of neurons. 26 SOD2 is a superoxide dismutase 2, which is upregulated in neuroprotection. 27 Furthermore, as previously reported SIRT3 directly binds and deacetylates SOD2, and promotes antioxidant activity. 28 The SIRT 3 can also protect motor neurons against the cell death in familial amyotrophic lateral sclerosis (ALS) 29 and neurodegeneration in Alzheimer's disease. 30
Considering the antioxidant properties of S. acmella Murr. and their possible therapeutic usages, the present investigation focused on exploring the antioxidant activity of S. acmella Murr. extracts and bioactive constituents on stress experimental model in human neuroblastoma SH-SY5Y cells and the underlying mechanisms. This study reports for the first time regarding the possible therapeutic applications of the natural compounds involved in degenerative disease, and new strategies for prevention of chronic stress.
Methods
Chemicals and reagents
Human dopaminergic neuroblastoma (SH-SY5Y) cell line was obtained from American Type Culture Collection (Manassas, VA). Nunclon™ culture flasks and Corning culture plates were obtained from Corning Incorporated (Acton, MA). The chemicals were purchased from the following sources: Dulbecco's modified Eagle's medium (DMEM)/Nutrient Mixture F-12 (Ham), fetal bovine serum, penicillin and streptomycin from Gibco BRL (Gaithersburg, MD), cellular apoptosis and autophagy kits from Millipore (Billerica, MA); antibodies used in Western blotting from Cell Signaling Technology, Inc. (Beverly, MA); enhanced chemiluminescence (ECL) plus Western blotting reagent from Amersham Biosciences (Piscataway, NJ); and MitoTracker and Alexa488-conjugated antirabbit from Invitrogen™ (Carlsbad, CA). Scopoletin (95%) and vanillic acid (98.5%) were purchased from ACROS organics. Trans-ferulic acid (99%) and fluoxetine were purchased from Sigma-Aldrich (St. Louise, MO). All other chemicals were of analytical grade and obtained from Sigma-Aldrich.
Plant extract preparation
EtOAc and MeOH extracts of the dried aerial parts of S. acmella Murr. were prepared as previously described 11 using EtOAc and MeOH to give the corresponding plant extracts. S. acmella Murr. has been identified (BKF 112361) by The Forest Herbarium, Royal Forestry Department.
Chemical profiles of the plant extracts (EtOAc and MeOH) include 1H-NMR (Supplementary Data), as well as HPLC was performed as described by Wongsawatkul et al. 11 1H-NMR spectra were recorded on a Bruker AVANCE 300 NMR spectrometer operating at 300 MHz using CDCl3 and CD3OD as solvents.
Cell culture and treatment
SH-SY5Y cells were maintained and cultured as previously described. 20 Once cells reached a confluency of 80%, the medium was discarded and washed with phosphate buffered saline (PBS). Cells were detached using 0.25% trypsin-EDTA solution and harvested by centrifugation at 1000 rpm for 5 minutes. The SH-SY5Y cells were resuspended in 1 mL medium and counted using trypan blue (0.4% w/v in PBS). Confluent cells were seeded onto 96- or 6 well plates at a density of 1 × 105 cells/mL.
SH-SY5Y cells were treated with different concentrations of dexamethasone (0.001, 0.01, 0.1, and 1 μM). To investigate the neuroprotective effect, S. acmella Murr. extracts or bioactive compounds were added to the medium for 24 hours before incubation with 1 μM of dexamethasone for 24 hours. Control-untreated cells were incubated with the culture medium. SH-SY5Y cells were treated with the S. acmella Murr. extracts or bioactive compounds according to the following protocols:
Pretreatment: SH-SY5Y cells were treated with S. acmella Murr. extracts (1 μg/mL) or bioactive compounds (1 μM). After 24 hours, the cells were then treated with 1 μM dexamethasone for an additional 24 hours.
Post-treatment: SH-SY5Y cells were treated with 1 μM dexamethasone for 24 hours, followed by a treatment with S. acmella Murr. extracts (1 μg/mL) or bioactive compounds (1 μM) for an additional 24 hours.
Determination of cell viability
Cell viability was evaluated using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay, which is based on the conversion of MTT from yellow to dark blue formazan crystals by mitochondrial dehydrogenase as previously described. 31 SH-SY5Y cells were cultured in 96 well plates at a density of 1.0 × 105 cells/mL. After each specific treatment, cells were treated with MTT and incubated in a 5% CO2 incubator at 37°C for 3–4 hours. The solution was discarded, then the extraction buffer (0.04 N HCl in isopropanol) was added. The absorbance of each reaction product was measured with a microtiter plate reader at a wavelength of 570 nm. The results are expressed as the percentage (%) of MTT reduction, with the assumption that the absorbance of the control cells is 100%.
Detection of cell apoptosis
Cell apoptosis was determined using fluorescein-5-isothiocyanate (FITC)-Annexin V apoptosis detection kit I. Cells were trypsinized at 37°C; 1 × 105 cells were labeled with the fluorochrome (5 μL Annexin V and 5 μL propidium iodide), and the cells were gently vortexed, then incubated with the reagent for 15 minutes at room temperature in the dark. At the end of incubation, the samples were analyzed by a fluorescence-activated cell sorter within 1 hour.
Detection of autophagy
Autophagy was measured using EMD Millipore's Muse™ Autophagy LC3-antibody–based kit. After the incubation, cells were washed with PBS and stained with autophagy reagent A for 4–6 hours at 37°C. Then, the cells were trypsinized and incubated with anti-LC3 at 4°C for 30 minutes in the dark. SH-SY5Y cells were harvested and determined using Muse™ Cell Analyzer.
Detection of cell reactive oxygen species
Intracellular production of reactive oxygen species (ROS) was monitored spectrofluorometrically using 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) as a fluorescence probe. After treatment, cells were incubated with 10 μM DCFH-DA for 30 minutes at 37°C in the dark. The fluorescence intensity was measured by the microplate fluorescence reader (Bio-Tek Instrumentation) at an excitation wavelength of 485 nm and an emission wavelength of 538 nm. Hydrogen peroxide (H2O2) 400 μM treated SH-SY5Y cells were used as a positive control. The level of ROS was expressed as a percentage value in relation to the control.
Proteomic analysis
SH-SY5Y control and dexamethasone-treated cells were precipitated to obtain protein by using 2-D Clean-up kit (GE Healthcare). Then, the protein pellets were dissolved in lysis buffer (30 mM Tris, 2 M thiourea, 7 M urea, 4% Zwitterionic non-denaturing detergent (CHAPS) [wt/vol] at pH 8.5). Two-dimensional gel electrophoresis was carried out using precast-immobilized pH gradient strips (18 cm, pH 3–10) on an Ettan IPGphor III system (GE Healthcare, Uppsala, Sweden) followed by 12.5% polyacrylamide gel electrophoresis. A sample containing 200 μg of total protein was mixed with 360 μL of rehydration buffer containing 1% pH 3-10 immobilized pH gradient (IPG) buffer and 1.5% destreak reagent (GE Healthcare). Samples were run through the steps of strip rehydration, and isoelectric focusing was carried out at 20°C for a total of 27,000 volt-hour.
After isoelectric focusing, the IPG strips were immediately used for the second dimension. The IPG strips were incubated in equilibration buffer (50 mM Tris-HCl [pH 8.8], 6 M urea, 30% [vol/vol] glycerol, and 2% [wt/vol] sodium dodecyl sulfate [SDS]) containing 2% (wt/vol) dithiothreitol (DTT) for 15 minutes with gentle agitation, followed by incubation in the same equilibration buffer supplemented with 2.5% (wt/vol) iodoacetamide for the same period as above at room temperature. The second dimension was carried out in 12.5% polyacrylamide gel at a constant power of 10 W per gel using an Ettan DALTsix (GE Healthcare) for 4 hours. Gels were stained overnight using colloidal Coomassie staining. After staining, gel images were acquired using Image scanner III (GE Healthcare).
Differential analysis was performed by ImageMaster 2D Platinum version 7.0 (GE Healthcare) software tool. Protein spots of interest were manually excised from preparative gels and digested using in-gel trypsin digestion. Peptides were reconstituted in 15 μL of 0.1% formic acid and identified by LC-MS (Dionex Ultimate 3000; Thermo Scientific, Waltham, MA) in combination with an electrospray ionization–ion trap mass spectrometry (Model Amazon SL; Bruker, Bremen, Germany). Mass spectral data from 300 to 1500 m/z were collected in the positive ionization mode. To identify proteins, all ms/ms spectra derived from each spot were searched against protein sequences from SwissProt databases using the MASCOT search program (
Detection of related proteins
Immunoblotting was performed following the standard procedures. Aliquots (10 μg of protein) of cell lysates were centrifuged at 14,000 rpm for 15 minutes at 4°C and electrophoresed in a 10%–12% SDS-acrylamide gel. Proteins were transferred electrophoretically to polyvinylidene difluoride (PVDF) membranes, which were blocked by 5% (w/v) nonfat milk in Tris-based immunoblot wash buffer (TBST) for 1 hour and further incubated with antibodies against dynamin-related protein 1 (DRP1), SIRT3, SOD2, or β-actin overnight at 4°C before incubation with horseradish peroxidase-conjugated secondary antibody. Signal detection was performed using an ECL plus Western blotting solution. The protein levels were quantified by densitometry using ImageJ software, and expressed relative to a housekeeping protein, such as β-actin.
Morphological assessment by inverted microscope
The morphology of treated cells and untreated cells was observed by inverted phase-contrast microscope (Olympus CKM41, Japan) at 20 × magnification, and the images were captured using digital camera (Nikon DS126311, Tokyo, Japan). Multiple independent images were taken for each experiment.
Immunocytochemistry
SH-SY5Y cells were allowed to settle in 24 well plates and coated with poly-L-lysine. The cells were then treated with or without 1 μM scopoletin for 24 hours followed by unchanging the medium, and further incubated with 1 μM dexamethasone for 24 hours. The cells were stained with MitoTracker Red and fixed with 4% paraformaldehyde. Then, the cells were permeabilized with 0.5% Triton X-100 and blocked with 5% bovine serum albumin (BSA). Cells were incubated with SIRT3 antibody in PBST overnight, then stained with FITC-conjugated antirabbit IgG for 2 hours at room temperature. The coverslips were mounted with ProLong® Gold Antifade (Molecular Probe®) and covered with glass slides. The images were captured with a confocal laser-scanning microscope (Fluoview FV10i, Olympus, Japan).
Statistical analysis
Statistical analysis was performed using GraphPad Prism 5.0 (GraphPad Software, Inc., San Diego, CA). All data represent at least four independent experiments, and are expressed as mean ± standard error of the mean; statistical comparisons were made using one-way analysis of variance (ANOVA). A p-value of <0.05 was considered statistically significant.
Results
Effect of dexamethasone on cell viability, apoptosis, and autophagy in SH-SY5Y cells
Initially, to determine the effect of dexamethasone on cell viability, SH-SY5Y cells were incubated at various concentrations (0.001, 0.01, 0.1, and 1 μM) of dexamethasone for 24 hours. The cellular viability was evaluated by MTT reduction assay, and was expressed as percentage of the control cells (100% cell viability). The cell viability was decreased by dexamethasone in a dose-dependent manner, with the half-cell viability loss at 1 μM, 44.65% ± 6.84% (p < 0.01) (Fig. 1A). Furthermore, Annexin-V/PI double staining was used to detect apoptosis by dexamethasone. Flow cytometric analysis showed that the presence of 1 μM dexamethasone significantly increased by 27.84% ± 2.42% (p < 0.001) for the early apoptosis, and 20.83% ± 1.17% (p < 0.001) for the late apoptosis as compared with the control (0 μM dexamethasone) (Fig. 1B, C).
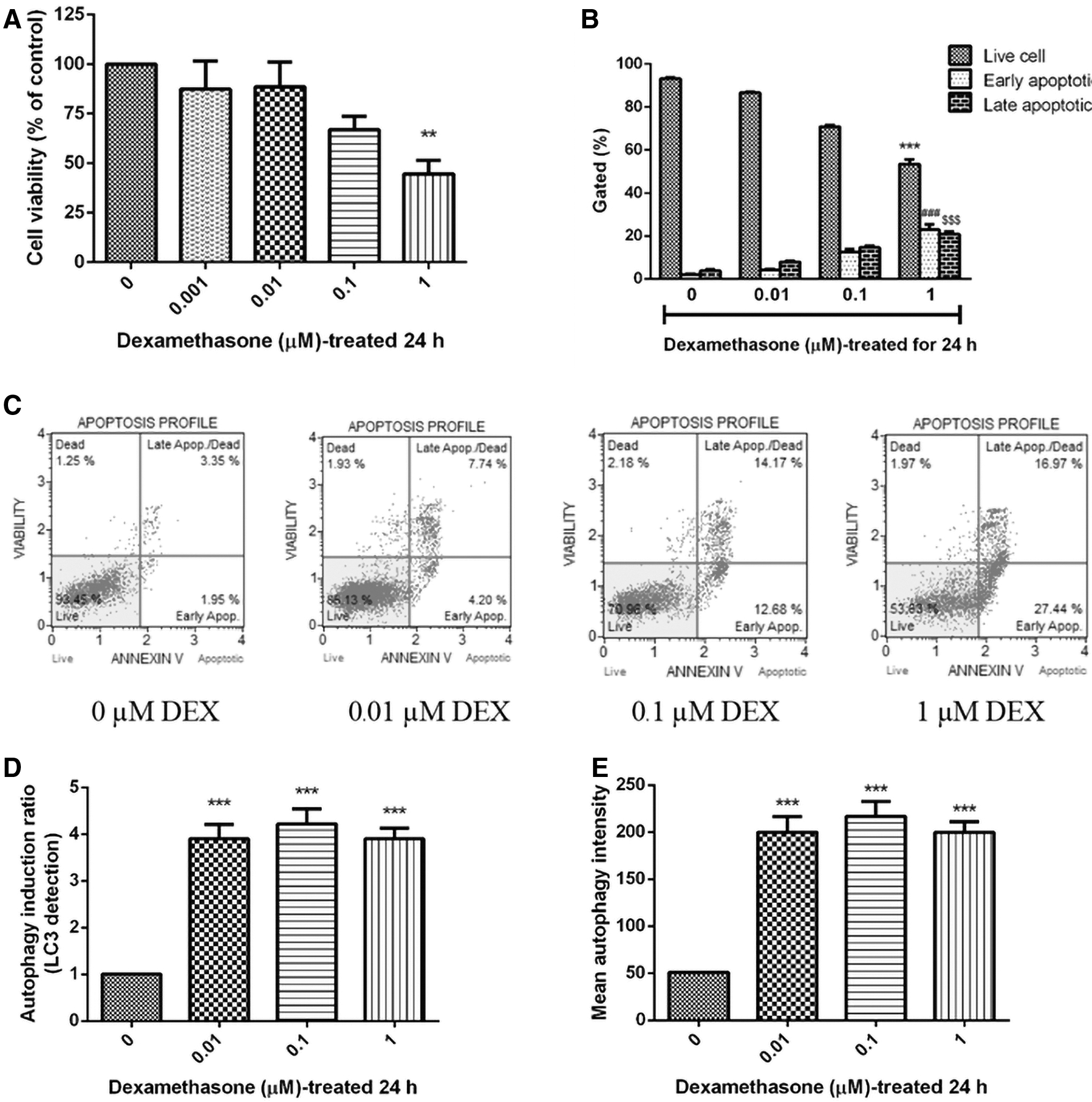
Effect of dexamethasone on cell viability apoptosis and autophagy, after treatments with 0, 0.01, 0.1, and 1 μM in SH-SY5Y cells.
The results indicated that dexamethasone caused cell apoptosis in a dose-dependent manner in SH-SY5Y cells. Therefore, 1 μM of dexamethasone was selected as the final concentration in the following experiments; while autophagy plays an active role in the dynamic cellular response to oxidative stress. The conversion of LC3 from the soluble form (LC3-II) to the autophagosome-associated form (LC3-II) implies an increase of autophagosomes within the cells. 32 The effect of dexamethasone on autophagy-related markers was determined by flow cytometry using anti-LC3 antibody. As shown in Figure 1D and E, dexamethasone (0.01, 0.1, and 1 μM) could increase the autophagy induction of SH-SY5Y cells to 3.90 ± 0.32, 4.22 ± 0.32, and 3.90 ± 0.23, respectively. These results suggested that dexamethasone induced the increase of autophagy induction in SH-SY5Y cells.
Proteomic profiling of proteins associated with dexamethasone-treated cells
Proteomics has emerged as an important tool to protein profiles. High-throughput techniques such as liquid chromatography-tandem mass spectrometry (LC-MS/MS) are able to provide effective profile of the proteins present in the dexamethasone-treated cells. LC-MS/MS detected a total of 920 proteins in the control (0 μM dexamethasone) and 789 proteins in the presence of 1 μM dexamethasone (Fig. 2A, B). Protein spots were searched against protein sequences from SwissProt databases using the MASCOT search program. Finally, Up- or downregulations of protein in dexamethasone-treated cells are summarized in Table 1.
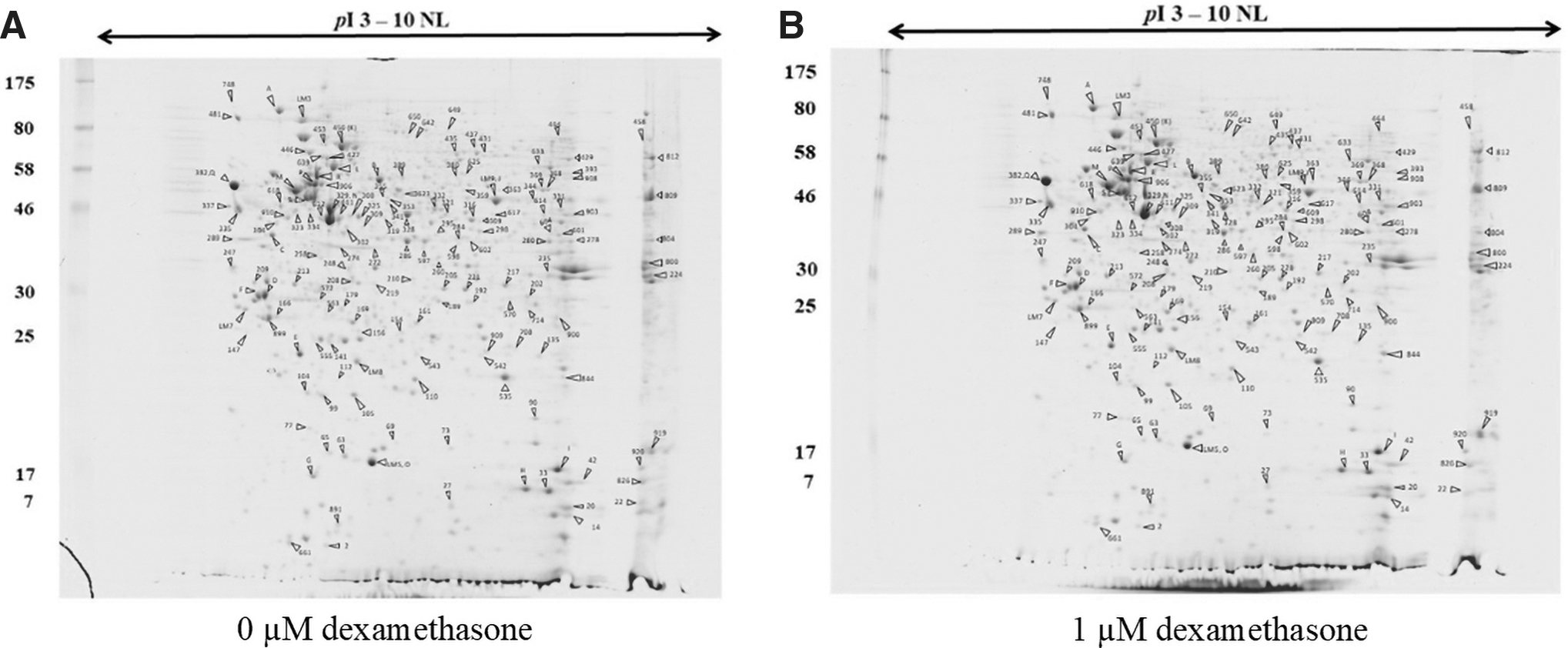
2D-PAGE image or the master maps representing protein profiles of SH-SY5Y cells,
The List of Differentially Abundant of Protein Treated with Dexamethasone in SH-SY5Y Cells
Each protein change ratio has been represented as a means of four individual values and standard deviations. The protein spots showing a statistically significant differential expression (p < 0.01) were selected.
PRR, postreplication repair.
In dexamethasone-treated cells, the five major groups of proteins with 26 protein spots were highlighted, including cytoskeleton organization such as actin cytoplasmic 1 (spot 329), actin (spot 489), and vimentin (spot 906); metabolic process such as L-lactate dehydrogenase A chain (spot 272), glucosidase 2 subunit beta (spot 481), and transaldolase (spot 597); protein biosynthesis such as RNA-binding protein 3 (spot 42), 26S proteasome non-ATPase regulatory subunit 14 (spot 210), elongation factor 1-alpha 1 (spot 809), and calumenin (spot 995); regulation of transcription proteins such as sorcin (spot 77), SETSIP (spot 289), zinc finger protein 143 (spot 325), and heterogeneous nuclear ribonucleoprotein D-like (spot 601); stress response such as superoxide dismutase [Mn], mitochondrial (spot 535) and cold-inducible RNA-binding protein (spot 919). However, after dexamethasone-treated cells, a strong signal was evident in biosynthesis and transcription regulation proteins.
In addition to identifying proteins upregulated by expression of
Dexamethasone-induced reduction in SOD2 and SIRT3 expressions in SH-SY5Y cells
Evidence of dexamethasone-induced mitochondrial ROS has been implicated in mitochondrial dynamics (unpublished results). The excessive level of ROS could change the antioxidant protein levels such as SOD2. Therefore, the effect of dexamethasone-induced ROS production on SOD2 protein expression was investigated. Results showed that SOD2 protein expressions were significantly diminished in dexamethasone (0.01, 0.1, and 1 μM) treatments, and were observed as 77.99% ± 17.80%, 47.05% ± 14.04% (p < 0.01), and 36.79% ± 7.14% (p < 0.001), respectively, when compared with the control (100%) (Fig. 3A).
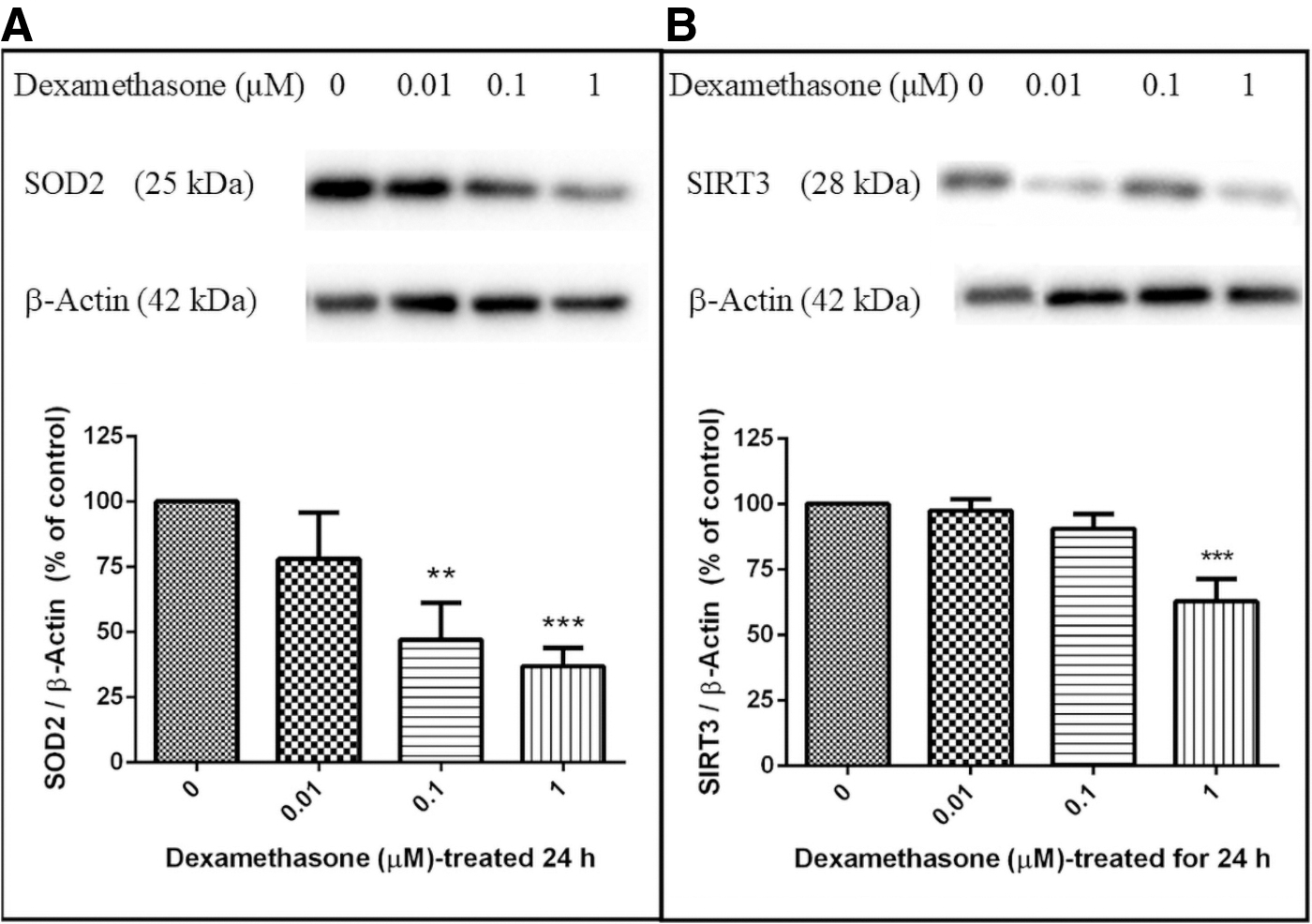
Dexamethasone-induced reduction of SOD2 and SIRT3 expressions in SH-SY5Y cells. SH-SY5Y cells were treated with 0, 0.01, 0.1, and 1 μM dexamethasone and collected after 24 hours treatment. Cell lysates were immunoblotted for SOD2
Furthermore, SOD2 is mainly regulated by the deacetylation of specific conserved lysines in a reaction catalyzed by the mitochondrial sirtuin, SIRT3. 33 As shown in Figure 3B, the level of SIRT3 was significantly decreased during 24 hours treatment with 1 μM of dexamethasone (62.78% ± 22.49% (p < 0.001)). These results suggested that suppression of SOD2 and SIRT3 signaling was involved in the dexamethasone-induced neurotoxicity in SH-SY5Y cells.
Neuroprotective effect of pre- and post-treatments with S. acmella Murr. extracts on dexamethasone-induced neuronal cell viability
Protective effect of pre- and post-treatments with S. acmella Murr. (MeOH and EtOAc) extracts on dexamethasone-induced neuronal cell death was investigated. SH-SY5Y cells were pretreated with 1 μg/mL of S. acmella Murr. extracts for 24 hours, followed by the addition of 1 μM dexamethasone for an additional 24 hours. The viability of SH-SY5Y cells treated with dexamethasone was shown to be 44.65% ± 6.34% (p < 0.001, Fig. 4A, B). However, pretreatment with S. acmella Murr. (MeOH and EtOAc extracts) could significantly increase the cell viability to 73.00% ± 2.44% (p < 0.01) and 68.37% ± 13.12% (p < 0.01), respectively, compared with the dexamethasone-treated group (Fig. 4A, B). Interestingly, MeOH and EtOAc extracts of S. acmella Murr. were found to be ineffective in post-treatments of the SH-SY5Y cells exposed to 1 μM of dexamethasone (Fig. 4A, B).
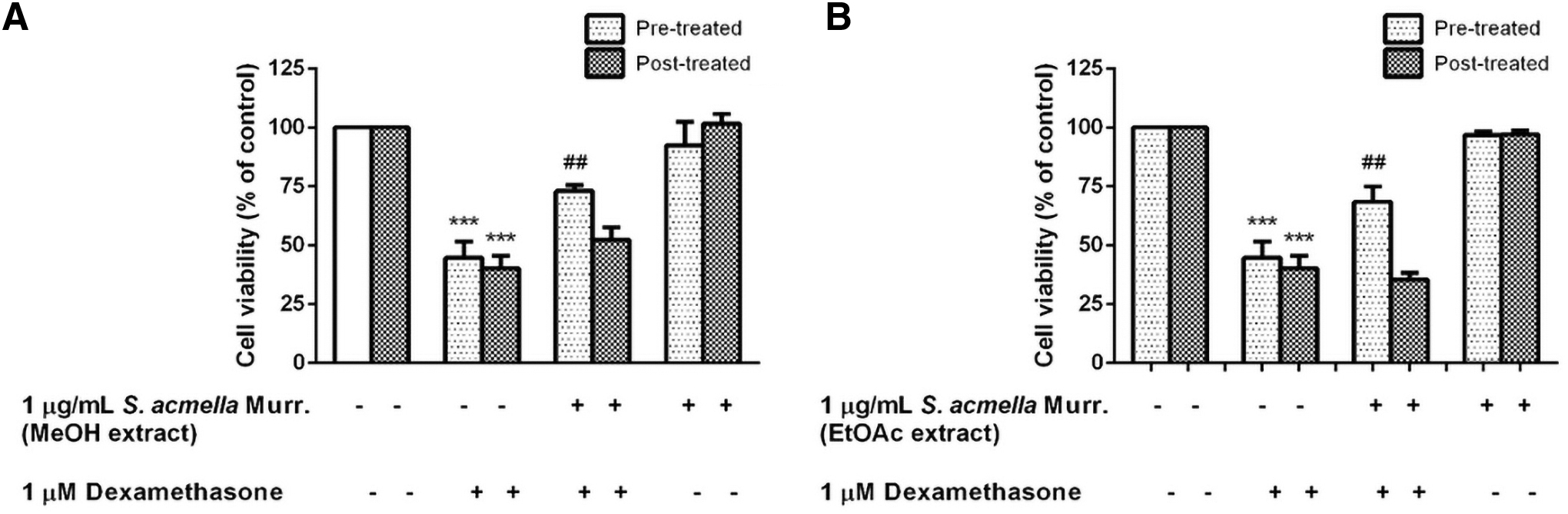
Neuroprotective effect of pre- and post-treatments with Spilanthes acmella Murr. extracts on dexamethasone-reduced cell viability by MTT assay.
Effect of scopoletin, vanillic acid, and trans-ferulic acid on antioxidant enzyme
To understand the antioxidant function of scopoletin, vanillic acid, and trans-ferulic acid (Fig. 5) in SH-SY5Y cells, different treatments were assessed via the Western blot analysis. SOD2 is the enzyme that plays a key role in regulating the ROS homeostasis. In this study, SH-SY5Y cells were exposed to 1 μM of scopoletin, vanillic acid, and trans-ferulic acid. The SOD2 protein levels were found to increase by 229.33% ± 42.19% (p < 0.05), 222.63% ± 39.64% (p < 0.05), and 187.89% ± 34.89% (p < 0.05), respectively, compared with the control (100%) (Fig. 5A). In addition, treatment of the cells with fluoxetine (a positive control) significantly increased the level of SOD2 expression to 109.87% ± 22.55% (p < 0.05) compared with the control-untreated cells.
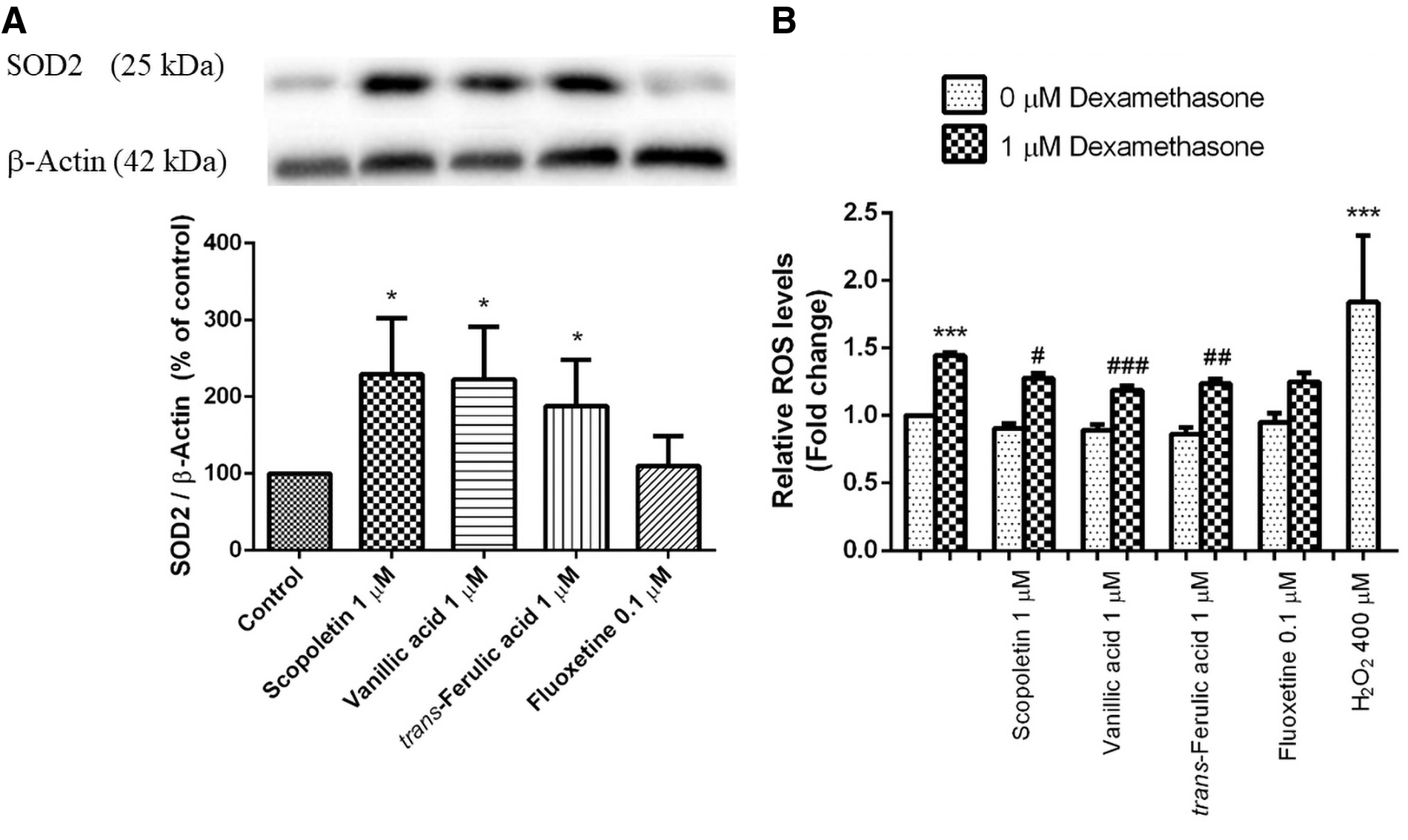
Effect of scopoletin, vanillic acid, and trans-ferulic acid on antioxidant enzyme. SH-SY5Y cells were treated with 1 μM scopoletin, vanillic acid, and trans-ferulic acid, and collected after 24 hours treatment.
The antioxidant property of these bioactive compounds might play a critical role in the free radical scavenging activity. As shown in Figure 5B, cell treatment with 1 μM dexamethasone significantly increased the intracellular ROS to 1.45 ± 0.04 (p < 0.001) fold change of the control cells, but the increase in intracellular ROS was significantly decreased to 1.28 ± 0.73 (p < 0.05), 1.19 ± 0.06 (p < 0.001), and 1.24 ± 0.08 (p < 0.01) fold change by pretreatment of the cells with 1 μM of scopoletin, vanillic acid, and trans-ferulic acid, respectively, compared with the positive control, 400 μM H2O2. These results showed that ROS induced by dexamethasone could be decreased by scopoletin, vanillic acid, and trans-ferulic acid.
Effect of scopoletin, vanillic acid, and trans-ferulic acid attenuated dexamethasone-induced neuronal cell death
As shown in Figure 6, treatment of the cells with 1 μM dexamethasone significantly decreased cell viability to 74.21% ± 3.69% (p < 0.001) compared with the control. However, pretreatment with 1, 10, and 100 μM scopoletin significantly increased cell viability to 101.30% ± 5.99% (p < 0.01), 100.70% ± 6.11% (p < 0.01), and 97.37% ± 6.30% (p < 0.05), respectively, compared with the control values (Fig. 6A). Pretreatment with 1, 10, and 100 μM vanillic acid significantly increased cell viability to 94.13% ± 3.56% (p < 0.01), 95.07% ± 4.68% (p < 0.001), and 96.54% ± 4.59% (p < 0.001), respectively, compared with the control values (Fig. 6B).
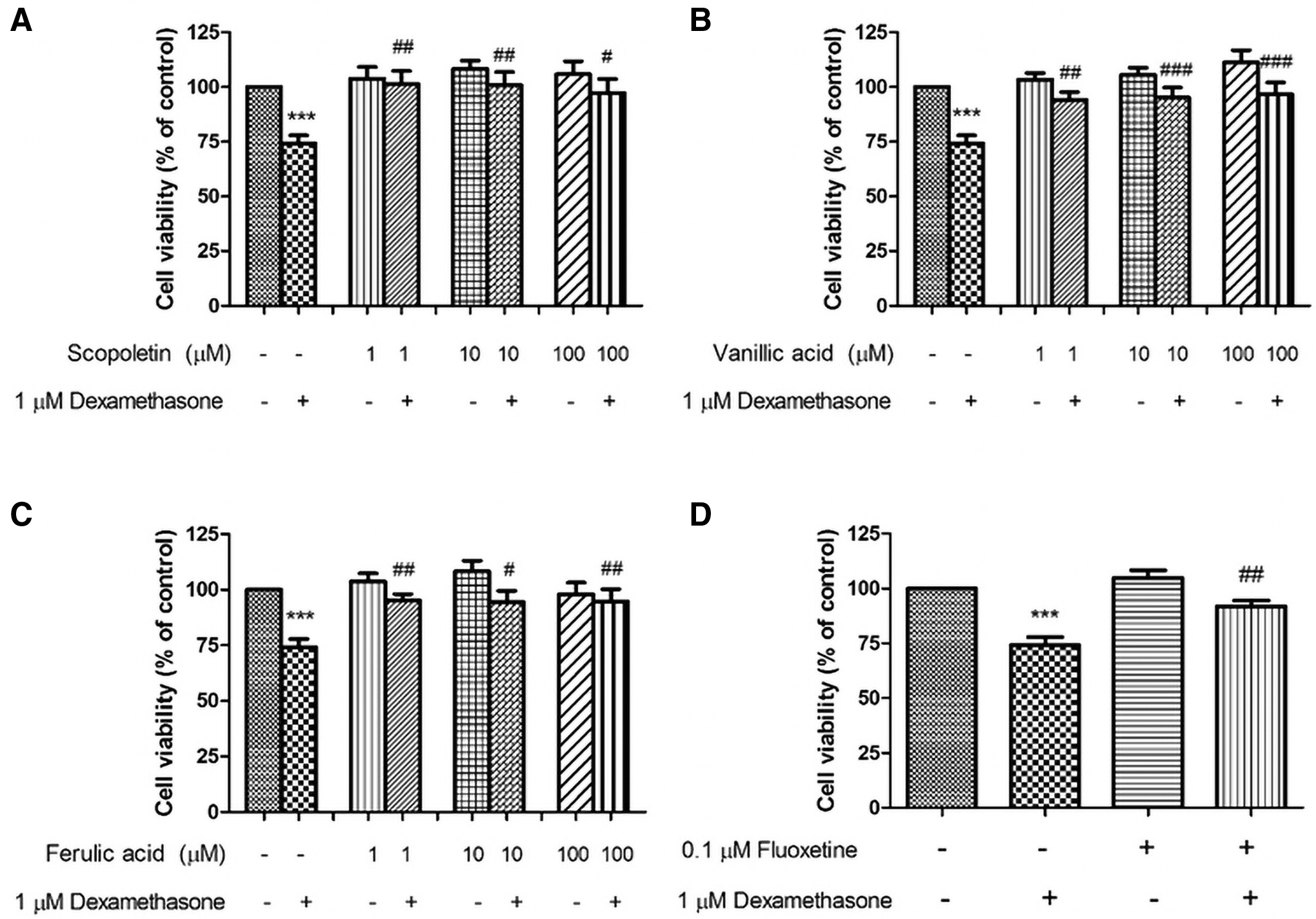
Scopoletin, vanillic acid, and trans-ferulic acid attenuated dexamethasone-induced neuronal cell death. SH-SY5Y cells were pretreated with scopoletin
Similarly, pretreatment with 1, 10, and 100 μM trans-ferulic acid significantly increased cell viability to 95.23% ± 2.82% (p < 0.01), 94.43% ± 5.14% (p < 0.05), and 94.71% ± 5.53% (p < 0.01), respectively (Fig. 6C). Finally, the cell pretreatment with 0.1 μM fluoxetine significantly increased the cell viability to 91.89% ± 2.64% (p < 0.01) when compared with dexamethasone-treated cell (74.21% ± 3.69%, p < 0.001; Fig. 6D). The results showed that scopoletin exhibited higher cell viability. Moreover, treatment with three of the bioactive compounds had no significant effects on the cell viability in untreated control cells.
Effect of scopoletin, vanillic acid, and trans-ferulic acid on dexamethasone-induced SIRT3 reduction in SH-SY5Y cells
To investigate an enhancement of SIRT3 expression by bioactive compounds was investigated on dexamethasone-induced neuronal cell death. The level of SIRT3 protein in the cells was determined by Western blot analysis (Fig. 7). Treatment with 1 μM dexamethasone significantly decreased SIRT3 expression to 75.85% ± 10.16% of the control (p < 0.01). In contrast, pretreatment of the cells with 1 μM scopoletin, vanillic acid, and trans-ferulic acid significantly increased SIRT3 expressions to 104.30% ± 8.53% (p < 0.01), 95.81% ± 18.19% (p < 0.05), and 97.65% ± 10.05% (p < 0.05) of the control, respectively (Fig. 7B); however, fluoxetine (0.1 μM) pretreatment significantly upregulated the SIRT3 expression to 90.39% ± 7.93% (p < 0.05) of the control.

Effect of scopoletin, vanillic acid, and trans-ferulic acid on dexamethasone-induced SIRT3 reduction in SH-SY5Y cell. SH-SY5Y cells were pretreated with scopoletin, vanillic acid, trans-ferulic acid, and fluoxetine prior to the incubation with 1 μM dexamethasone for another 24 hours. Results are expressed as the mean ± SEM, and were analyzed using ANOVA for four independent experiments. **p < 0.01 compared with the control-untreated cells, and # p < 0.05 and ## p < 0.01 compared with the dexamethasone-treated cells.
Meanwhile, the Western blotting markedly showed the increase of SIRT3 expression in scopoletin-pretreated SH-SY5Y cells (Fig. 7A). The protective effect of the bioactive constituents in order of potency was scopoletin > trans-ferulic acid > vanillic acid. These results were also confirmed by immunofluorescence studies of SIRT3 in SH-SY5Y cells. Treatment of the cells with dexamethasone (1 μM) resulted in the decrease of SIRT3 expression (in green) (Fig. 8D), which was significantly elevated by the scopoletin pretreatment (Fig. 8G). However, the red color indicated active mitochondrial sites, upon treatment with dexamethasone the decrease in mitochondrial site signals was observed (Fig. 8E) when compared with the control cells (Fig. 8B). The results indicated that scopoletin may play an important role in the regulation of SIRT3 expression after the oxidative stress caused by dexamethasone treatment.

Effect of scopoletin on SIRT3 expression in SH-SY5Y cells. SH-SY5Y cells were treated with 1 μM scopoletin for 24 hours and exposed to 1 μM dexamethasone. The green color indicates SIRT3 immunostaining using FITC-conjugated anti-IgG, and the red color indicates mitochondrial sites using MitoTracker Red CMXRos. Scale bar = 10 μm. FITC, fluorescein-5-isothiocyanate.
Scopoletin suppresses dexamethasone-induced autophagy through SIRT3 expression in SH-SY5Y cells
Previously the activation of autophagy was reported to associate with neuronal cell death. 7 In this study, protective effect of scopoletin on dexamethasone-induced toxicity in SH-SY5Y cells was examined using flow cytometry. After treatment of the cells with dexamethasone, the induction of autophagy was shown to increase as 2.27 ± 0.63 (p < 0.001) compared with the control-untreated cells (Fig. 9). Pretreatment of the cells with scopoletin, acting as antioxidant and SOD2 activator, resulted in the decrease of autophagy (p < 0.01) compared with the treated cells. In addition, the autophagy induction was not affected in the scopoletin-treated cells. These findings suggested that scopoletin may reduce autophagy through the activation of SOD2-SIRT3 signaling pathways.
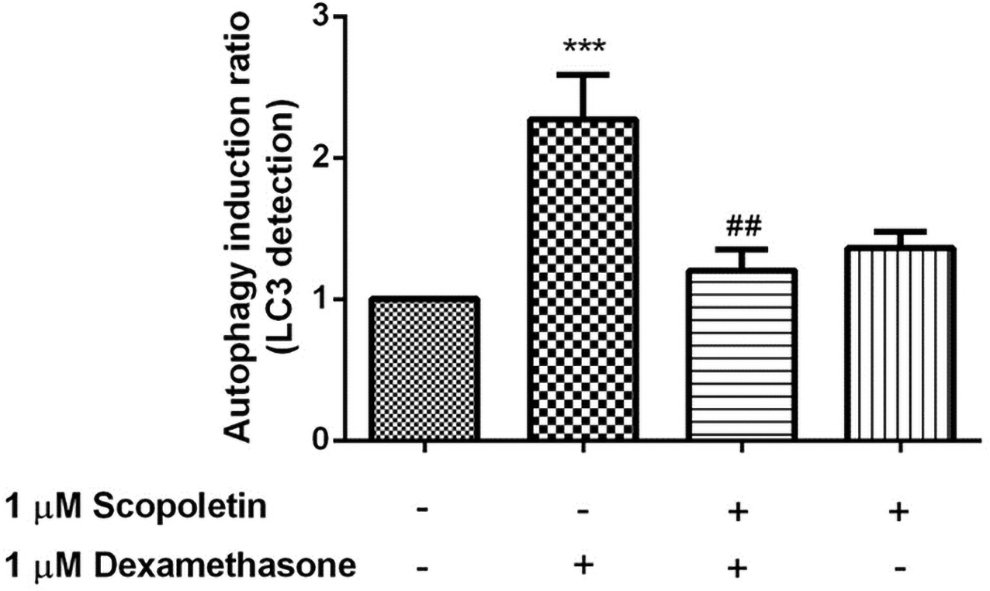
Protective effect of scopoletin on dexamethasone-induced autophagy in SH-SY5Y cells. SH-SY5Y cells were pretreated with 1 μM scopoletin before incubation with 1 μM dexamethasone for another 24 hours, and the LC3 level was measured using flow cytometry. Data are expressed as mean ± SEM, and were analyzed using ANOVA for at least four independent experiments. ***p < 0.001 compared with the control-untreated cells, and ## p < 0.01 compared with the dexamethasone-treated cells.
Discussion
Dexamethasone has many medical advantages for treatment of cancer, inflammatory and autoimmune diseases. Under the excess of dexamethasone conditions, dexamethasone induced apoptosis in osteoblast, 34 alveolar macrophages, 35 glioblastoma neurospheres, 36 and dopaminergic cells. 37 In recent years, a growing number of studies have indicated that dexamethasone exposure can cause cognitive impairment with low-functional programming of the hippocampal CREB/BDNF/TrkB signaling pathway. 38,39
Autophagy has been demonstrated to be a process that maintains the cellular homeostasis. However, the excessive activation of autophagy could induce programmed cell death. 40 The present study revealed that administration of a high dose of dexamethasone (1 μM) can induce apoptosis, autophagy and reduce neuronal cell survival. Such findings have demonstrated that dexamethasone induced neuronal cell death through SIRT3 and SOD2 reductions. These results are consistent with the previous study of chondrocytes. 41 In fact, SIRT3 has been reported to regulate ROS generation, whereas SOD2 plays role in catalyzing the dismutation of superoxide radicals into oxygen and H2O2.
Furthermore, S. acmella Murr. extracts prevented the dexamethasone-induced neuronal cell death when administered as pretreatment (as preventive) and post-treatment (as therapeutic). Pretreatment of the neuronal cells with S. acmella Murr. extracts resulted in an enrichment of antioxidants, and the increase of cell survival against the dexamethasone-induced neurotoxicity. It has been reported that S. acmella Murr. extract maintained calcium homeostasis in pesticide-induced neurotoxicity, 20 improved mitochondrial dysfunction in chronic restraint stress, 42 inhibited nuclear factor-κB and mitogen-activated protein kinase signaling pathways in lipopolysaccharide (LPS)-stimulated macrophages. 43
Previous studies showed that scopoletin, vanillic acid, and trans-ferulic acid (bioactive compounds isolated from S. acmella Murr. extract) exerted the preventive potential for the oxidative damage. 44 –46 The bioactive compounds displayed the increase of SOD2 expression, which might be considered as a free radical scavenger. This study showed that scopoletin, vanillic acid, and trans-ferulic acid could inhibit dexamethasone-induced neurotoxicity (Fig. 6). This is consistent with the ability of such bioactive compounds to reduce ROS generation and neuronal cell damage. The mechanism of ROS reduction involved the induction of SOD2 (Fig. 5) and SIRT3 protein levels (Fig. 7). Apparently, scopoletin, vanillic acid, and trans-ferulic acid exerted better effect in reducing the oxidative damage compared with the fluoxetine (positive control) in in vitro studies.
It has been first demonstrated that S. acmella Murr. extracts and their bioactive compounds act as neuroprotectants in a model of depression through SIRT3/SOD2 signaling pathways. SIRT3 is primarily active in neuronal mitochondria, and directly contributes to neuroprotection in Alzheimer's, 30 Parkinson's, 47 and Huntington's 48 diseases as well as ALS. 49 Expression of SIRT3 is associated with mitochondrial function, including mitochondrial biogenesis, mitochondrial dynamic, 50 and mitochondrial respiration. 30
Importantly, SIRT3 regulates ROS production through the activation of the antioxidant enzymes such as SOD2, isocitrate dehydrogenase 2 (IDH2), glutathione (GSH): glutathione disulfide (GSSG) ratio, and peroxiredoxin (Prx3). 50 SIRT3 interacts with FoxO3a and enhances FoxO3a DNA binding to the SOD promoter. 51 The recent experimental study demonstrated that toxicity of dexamethasone inhibits SIRT3 and SOD2 expressions in SH-SY5Y cells. These results are in agreement with the fact that SIRT3 and SOD2 reductions exhibited excessive ROS generation and finally cell death. This is significantly alleviated when the cells were pretreated with the bioactive compounds.
Summing up, this study demonstrates that the bioactive constituents of S. acmella Murr. extracts significantly increased the neuronal SH-SY5Y cell viability through the modulation of SIRT3/SOD2-mediated pathways. In this regard, scopoletin, vanillic acid, and trans-ferulic acid potentiated the antioxidant effect on dexamethasone-induced neuronal cell death. In this light, S. acmella Murr. extracts could be developed as a new strategy for the treatment of neurotoxicity.
Footnotes
Authors' Contributions
W.S. designed, analyzed the data, and prepared a draft of the article. C.S. collected the samples. S.S. collected S. acmella Murr. and prepared the plant species extracts. C.I.-N.-A. contributed to proteomic analysis. S.P. performed proteomic analysis and the chemical profiles, and studied the article.
Acknowledgments
This research project was supported by the Mahidol University.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.