
Abstract
The current understanding of skin aging is that senescent fibroblasts accumulate within the dermis and subcutaneous fat to cause abnormal tissue remodeling and extracellular matrix dysfunction, triggering a senescence-associated secretory phenotype (SASP). A novel therapeutic approach to prevent skin aging is to specifically eliminate senescent dermal fibroblasts; this requires the identification of specific protein markers for senescent cells. Apolipoprotein D (ApoD) is involved in lipid metabolism and antioxidant responses and is abundantly expressed in tissues affected by age-related diseases such as Alzheimer's disease and atherosclerosis. However, its behavior and role in skin aging remain unclear. In this study, we examined whether ApoD functions as a marker of aging using human dermal fibroblast aging models. In cellular senescence models induced through replicative aging and ionizing radiation exposure, ApoD expression was upregulated at the gene and protein levels and correlated with senescence-associated β-galactosidase activity and the decreased uptake of the proliferation marker bromodeoxyuridine, which was concomitant with the upregulation of SASP genes. Furthermore, ApoD-positive cells were found to be more abundant in the aging human dermis using fluorescence flow cytometry. These results suggest that ApoD is a potential clinical marker for identifying aging dermal fibroblasts.
Introduction
Skin aging is caused by both endogenous factors stemming from inevitable physiological processes and exogenous factors such as air pollution, smoking, nutritional deficiencies, and sun exposure. As a result, the skin acquires characteristics such as coarse wrinkles, loss of elasticity, laxity, and a rough appearance. 1,2 These changes are representative of the process of cellular aging, in which the proliferative capacity of cells such as keratinocytes, fibroblasts, and melanocytes is reduced. 3
The emerging hypothesis of skin aging postulates that the senescence of fibroblasts primarily causes skin deterioration by promoting irreversible proliferative arrest and the release of senescence-associated secretory phenotype (SASP) factors. The SASP involves chronic inflammation through chemokines and proinflammatory factors and the inhibition of proliferation through an impaired release of essential growth factors. Particularly in skin, SASP factors promote extracellular matrix degradation by triggering the activation of proteolytic enzymes including matrix-degrading metalloproteinases. 4,5 Fibroblast senescence is primarily a persistent and permanent condition that results in skin aging, general aging, and aging-related diseases. 6 –8 In skin samples from human donors of various ages, senescent fibroblasts have been observed predominantly within the dermis. 9,10 Therefore, given that fibroblasts trapped in connective tissues stabilize all skin parenchyma, cellular senescence in these cells plays a central role in skin aging.
Targeted elimination of senescent fibroblasts or inhibition of SASP factors could arrest the progression of aging and may even rejuvenate the skin. SASP factors are associated with the disease-enhancing effects of senescent cells that accumulate in aging tissues, and hence, they are used to identify the occurrence of cellular senescence. However, SASP is not restricted specifically to senescent cells, and is therefore a poor biological marker for this type of cell. For this reason, discovering unique markers that function as senescence-associated autoantigens would enable targeted clearance of senescent cells, which is a potential therapeutic strategy to slow aging-related processes. In fact, systemic rejuvenation, improvement of aging-related diseases, and prolongation of life span have been reported following the removal of senescent cells from the body. 11 –13
This can be achieved by identifying potential markers and therapeutic targets among the diverse cell surface proteins, based on the fact that senescent cells differ significantly from proliferating cells in their protein expression patterns. So far, DPP4, 14 PLAUR, 15 and ACKR3, which we discovered, 16 have been identified as membrane proteins specifically expressed in senescent cells. In addition, senescent cell-specific proteins include p16ink4a and SFRP4. 11,17 These targetable senescence markers may be important in developing interventions aimed at eliminating senescent cells.
Apolipoprotein D (ApoD) is a glycosylated protein of ∼25 kDa, a member of the lipocalin superfamily, and widely expressed in mammalian tissues. 18 ApoD is involved in lipid transport, food intake, inflammation, antioxidant response and development, and various types of cancer, as well as being a multifunctional protein interacting with multiple ligands. 19 ApoD levels have been reported to be significantly elevated with age and in neuropathologies such as Alzheimer's disease and Parkinson's disease. 20,21 Furthermore, an increased ApoD deposition is detectable in atherosclerotic lesions in patients with established cardiovascular diseases and in mice with early atherosclerosis. 22 Thus, ApoD has been considered an important player in lipid metabolism and aging, but its relevance to skin aging has not been discussed. ApoD mRNA transcripts are expressed primarily by fibroblasts, especially by those located near blood vessels, 23,24 but the dermal behavior of ApoD in fibroblasts remains unknown.
To fill this knowledge gap, we investigated the behavior of this protein in a human dermal fibroblast model of senescence. The methods for the induction of this model (repeated passaging and use of ionizing radiation) were based on previous studies. 4,25 We hypothesized that ApoD is specifically expressed in association with an SASP in the dermal fibroblasts of aging epidermal or dermal cell models. We also investigated the possibility of ApoD being a robust dermal fibroblast surface antigen marker for human aging.
Materials and Methods
Cell culture
Normal human dermal fibroblasts (NHDFs), a pooled cell population containing cells from multiple donors, were obtained from PromoCell GmbH (Heidelberg, Germany). NHDFs were grown and cultured in low-glucose Dulbecco's modified Eagle's medium (Wako Pure Chemical Industries, Osaka, Japan) supplemented with 1% penicillin/streptomycin (Thermo Fisher Scientific, Waltham, MA) and 10% fetal bovine serum (Thermo Fisher Scientific). Replicative senescence was defined as a cell population doubling level (PDL) greater than 50 and an absence of growth for more than 2 weeks. Ionizing-radiation-induced senescence was induced in the same manner as previously reported. 16 Cells were irradiated with 10 Gy of X-ray radiation using the AB-160 X-Ray Irradiation System (AcroBio, Tokyo, Japan) and analyzed 10 days later. Control (proliferating) cells were mock irradiated by transporting them from the incubator to the irradiator but not irradiating them. Their growth outside the irradiator was maintained for the same period of time as the irradiated cells.
Intracellular senescence-associated beta-galactosidase (SA-β-gal) activity was assessed by staining the cells using the Senescence β-Galactosidase Staining Kit from Cell Signaling (Danvers, MA).
Evaluation of proliferative capacity through measuring bromodeoxyuridine uptake
Bromodeoxyuridine (BrdU) uptake was assessed using the Frontier BrdU Cell Proliferation Assay (Exalpha Biologicals, Inc., Shirley, MA) according to the manufacturer's protocol. SpectraMax Paradig (Molecular Devices, San Jose, CA) was used to analyze sample absorbance at wavelengths of 450/550 nm.
Immunocytochemistry
Cells were placed on circular cover glasses, allowed to stand for 15 minutes to adhere, and then fixed in acetone for 5 minutes at room temperature (15°C–25°C). After washing the cells three times with chilled phosphate-buffered saline (PBS), the cells were blocked with PBS containing 3% fetal bovine serum at room temperature for 1 hour. The cells were then incubated overnight at 4°C with the anti-ApoD antibody (ab108191; Abcam, Cambridge, United Kingdom) and anti-interleukin (IL)-6 antibody (ab6672; Abcam) diluted 1:200 in PBS. To confirm the specificity of the immunostaining, fetal bovine serum diluted to the same concentration (1:200) was used in place of the primary antibody as the negative control.
After washing the cells three times with PBS, slides were incubated with the Alexa Fluor 488-labeled goat anti-rabbit antibody (Thermo Fisher Scientific) and the Alexa Fluor 555-labeled donkey anti-goat antibody (Thermo Fisher Scientific) diluted 1:2000 in PBS at room temperature for 1 hour. After incubation, the cells were washed three times with PBS and counterstained with ProLong Gold Anti-fade Mountant (Thermo Fisher Scientific) containing 4′,6-diamidino-2-phenylindole to visualize the nuclei.
Immunohistochemistry
Paraffin-embedded whole skin sample slides of 3- and 88-year-old human trunks were purchased from OriGene Technologies, Inc. (Rockville, MD) with ethical approval. Samples were collected after informed consent was obtained from anonymous donor patients.
Slides were washed and deparaffinized with xylene three times for 5 minutes at room temperature, then soaked twice in 100% ethanol (3 minutes per soaking step), then soaked in 95%, 70%, and 50% ethanol consecutively (3 minutes each), and finally rehydrated at room temperature. Next, the slides were immersed in 0.01 M citrate buffer (pH 6.0) at 95°C for 1 hour for antigen activation before being incubated with 2% fetal bovine serum in PBS for 60 minutes at room temperature to block nonspecific binding sites. The slides were then incubated overnight at 4°C with the same anti-ApoD antibody (1:200 dilution in PBS) used for immunocytochemistry. After being washed three times with PBS, the slides were incubated with the biotinylated rabbit anti-goat antibody (Vector Laboratories, Burlingame, CA) (1:500 dilution in PBS) for 1 hour at room temperature.
The signal was amplified by using the avidin–biotinylated peroxidase complex method with 3,3′-diaminobenzidine solution. Briefly, the samples were incubated with the VECTASTAIN ABC Kit (Vector Laboratories, Newark, CA) and 20 mg/dL 3,3′-diaminobenzidine solution (FUJIFILM Wako Pure Chemicals, Co., Osaka, Japan) for 1 hour at room temperature.
The sections were rinsed with tap water for 5 minutes and then nuclear-counterstained with Gill's hematoxylin solution (Merck Millipore, Billerica, MA) for 6 seconds. Finally, sections were rinsed with tap water for 5 minutes, dehydrated with ethanol twice (each at 95% and 100% concentration for 5 minutes), rinsed three times with xylene, and finally sealed with Mount Quick Sealant (Takara Bio, Shiga, Japan).
For fluorescent staining, after blocking, the slides were incubated with the anti-ApoD antibody (1:200 dilution in PBS) and anti-p16ink4a antibody (1:200 dilution) at 4°C overnight. After washing three times with PBS, slides were incubated with the Alexa Fluor 488-conjugated goat anti-rabbit antibody (Thermo Fisher Scientific) and the Alexa Fluor 555-conjugated donkey anti-goat antibody (Thermo Fisher Scientific) diluted 1:2000 in PBS and incubated for 1 hour at room temperature. After incubation, slides were washed three times with PBS and counterstained with ProLong Gold Anti-fade Mountant (Thermo Fisher Scientific).
All slides were observed using an integrated stereomicroscope (BZ-X800; KEYENCE, Osaka, Japan).
RNA extraction and reverse transcription
Total RNA was extracted from cells using an RNeasy Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. Total RNA was mixed with random primers, reverse transcriptase, and deoxynucleotide mixture (Takara Bio). The mixture was annealed in a T100™ thermal cycler (Bio-Rad Laboratories, Inc., Hercules, CA) at 25°C for 5 minutes and synthesized at 55°C for 10 minutes. The reverse transcriptase was heat-inactivated at 80°C for 10 minutes to obtain cDNA.
Reverse transcription quantitative real-time polymerase chain reaction
Real-time quantitative polymerase chain reaction (RT-qPCR) was performed on an Applied Biosystems 7500 Fast Real-Time PCR System (Thermo Fisher Scientific) for 40 cycles and the fluorescence of each sample was measured at the end of each cycle. The PCR was performed in two major steps: (1) 95°C for 3 seconds (denaturation) and (2) 60°C for 30 seconds (annealing and extension). For the subsequent melting curve analysis, the temperature was increased from 60°C to 95°C and fluorescence was measured continuously. Gene expression was analyzed for ApoD (assay ID: 4331182 Hs00155794_m1), IL-6 (Hs00985639_m1), IL-8 (Hs00174103_m1), p16INK4a (Hs00923894_m1), MMP3 (Hs00968305_m1), and MMP9 (Hs00957562_m1) using gene-specific primers (Thermo Fisher Scientific) and a PCR master mix (4352042; Applied Biosystems, Foster City, CA). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (Hs02786624_g1) was used as the control gene for normalization. Gene expression levels of proliferating cells were used as baseline and relative expression levels were determined using the 2-ΔΔCT method.
Western blotting
Total protein was extracted from cells using RIPA Lysis Buffer (Santa Cruz Biotechnology, Santa Cruz, CA, USA) according to the manufacturer's protocol. Each sample (40 μg) was electrophoresed on a 10% polyacrylamide gel (Mini-PROTEAN TGX Precast Gels; Bio-Rad Laboratories, Inc.) and transferred onto polyvinylidene chloride membranes (Bio-Rad Laboratories, Inc.) using the Trans-Blot Turbo Transfer System (Bio-Rad Laboratories, Inc.).
After blocking with 3% nonfat milk dissolved in Tris-buffered saline with Tween 20 for 1 hour at room temperature, the cells were incubated with antibodies, namely, anti-ApoD (ab108191; Abcam), anti-p21 (ab220206; 1:100; Abcam), anti-CDKN2A/p16INK4a (EPR1473; Thermo Fisher Scientific; 1:200), anti-SIRT1 (ab32441; 1:200; Abcam), and anti-GAPDH (1:2000; Santa Cruz Biotechnology). Dilutions were made in blocking solution, and the cells were incubated overnight with the antibodies at 4°C. On the following day, the samples were incubated with secondary antibodies, specifically, donkey anti-goat IgG H&L [horseradish peroxidase (HRP)] (ab6885; Abcam), goat anti-rabbit IgG H&L (HRP) (ab205718; Abcam), and goat anti-mouse IgG H&L (HRP) (ab205719; Abcam) at a 1:1000 dilution for 2 hours at 37°C. After washing, immunoreactive protein bands were visualized using an electrochemiluminescence detection kit (Pierce Biotechnology, Rockford, IL).
Images of the bands were captured using a chemiluminescence imager (ImageQuant LAS4000mini; GE Healthcare, Chicago, IL). Image analysis was performed on ImageJ software (version 1.53p; National Institutes of Health, Maryland). Each experiment was repeated thrice.
Flow cytometry
Proliferating and senescent human skin fibroblasts were counted using TC20 Cell Counter (Bio-Rad Laboratories, Inc.) and washed with fluorescence activated cell sorting (FACS) buffer (0.5% bovine serum albumin in PBS). After washing, human TruStain FcX (BioLegend, San Diego, CA) was added to the cells to block Fc receptors, and then the cells were incubated with the anti-ApoD antibody (ab108191; Abcam) at 4°C for 10 minutes. Next, the cells were incubated with the Alexa Fluor 488-conjugated rabbit anti-goat antibody (Thermo Fisher Scientific) for 15 minutes at 4°C in the dark. Then, 7-aminoactinomycin D (7AAD) (Immunostep, S.L., Salamanca, Spain) was added for incubation at 4°C for 15 minutes to label the photoreceptor cells. Analysis was performed on FlowJo software (version 10.2). Briefly, the background autofluorescence of the negative population was measured using unstained controls. 7AAD-viable cells were then gated, and the number of Alexa Fluor 488-positive cells among them was counted.
Statistical analysis
Statistical analysis was performed using the software programs GraphPad Prism (version 5.0; San Diego, CA) and SPSS 22.0 (Chicago, IL). The Mann–Whitney U test was used to examine statistical significance. One-way analysis of variance and Tukey's post hoc test were used to compare differences among three or more groups. Values where p < 0.05 were considered statistically significant.
Institutional review board statement
Human skin samples complied with the ethical guidelines for life science and medical research involving human subjects.
Results
ApoD is specifically expressed in senescent human dermal fibroblasts
In cells that have undergone long-term culture (PDL >50), the replicative-senescence (ReS) and radiation-induced-senescence (RaS) cell groups exhibited flattened and expanded cytoplasm and an increased SA-β-gal activity (Fig. 1A). Furthermore, analysis of BrdU uptake showed that proliferative activity was significantly lower in senescent cells than in proliferating cells (young vs. RaS, p = 0.000082; young vs. ReS, p = 0.000077; Fig. 1B).
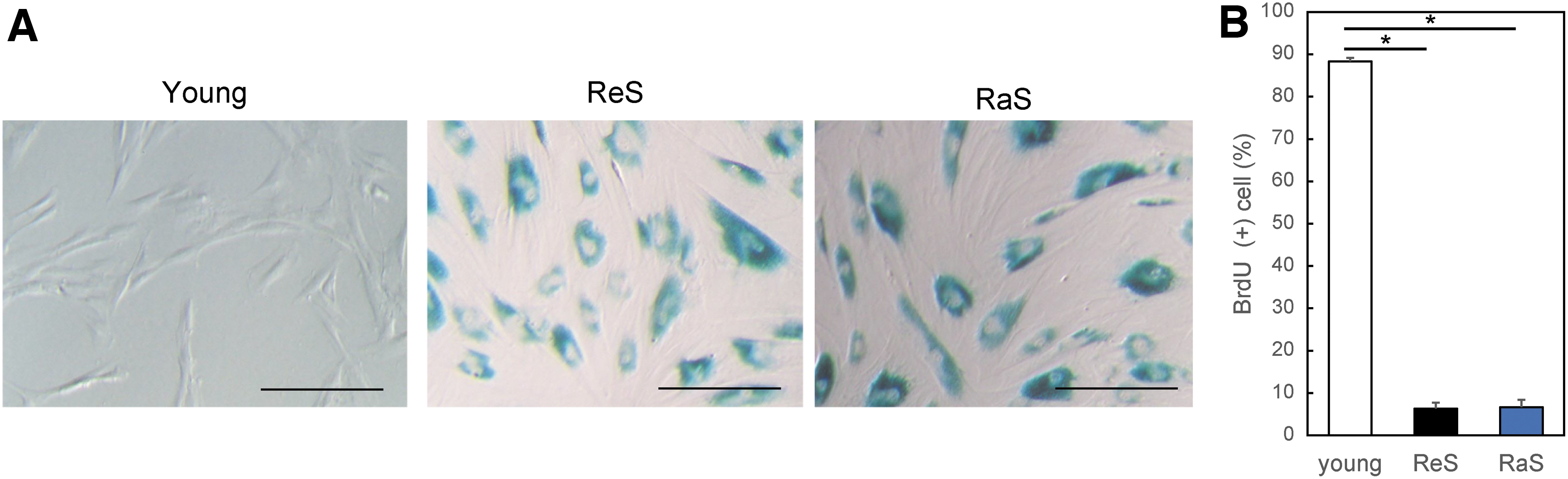
Induction of senescent cells by replication and irradiation.
In addition, coimmunostaining for p16INK4a, one of the markers of senescent cells, alongside ApoD showed that neither p16INK4a nor ApoD was expressed in the proliferating young cells, whereas these proteins were expressed in the two aging models (Fig. 2A). Protein-level analysis following Western blotting showed that the protein expression levels of p16INK4a and ApoD were higher in senescent cells than in control young cells (Fig. 2B). In addition, the expression of SIRT1, a marker of young cells, was decreased in senescent cells.
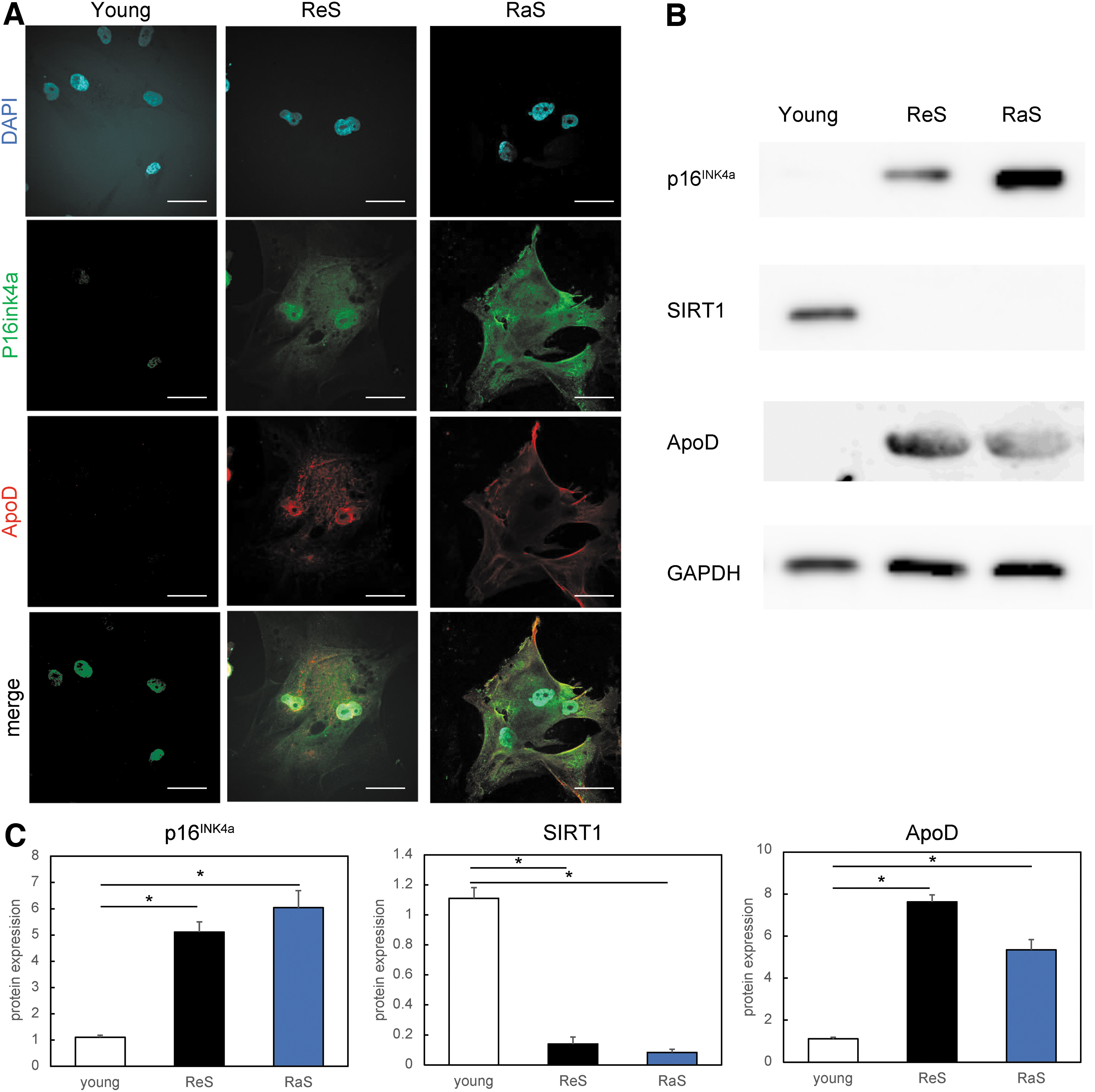
ApoD is upregulated in human senescent dermal fibroblasts.
ApoD is upregulated concomitantly with SASP factors
RT-qPCR results showed that in all senescent cell models, the ApoD gene was upregulated when compared with its gene expression level in proliferating (young) cells (Fig. 3). This was concomitant with increased gene expression levels of SASP factors such as IL6, IL8, p16INK4a, MMP3, and MMP9. Thus, we speculate that ApoD may be a marker for fibroblasts exhibiting an SASP.

Expression level analysis of senescence-associated secretory phenotype factor genes and ApoD gene. Experiments were performed independently in triplicate. *p < 0.05.
ApoD is upregulated in the aging human skin dermis
We next investigated the expression of ApoD in human skin as a marker of aging by immunostaining human skin samples from 3- and 88-year-old patients. Within the dermis of young human skin, ApoD-positive cells were infrequently observed, whereas multiple ApoD-positive cells were observed in aged skin (Fig. 4A). Furthermore, costaining for ApoD and p16ink4a, an existing senescent cell marker, revealed a large number of cells double positive for these molecules in the aging dermis (Fig. 4B). This result suggests that ApoD may also be an in vivo marker of dermal senescence.
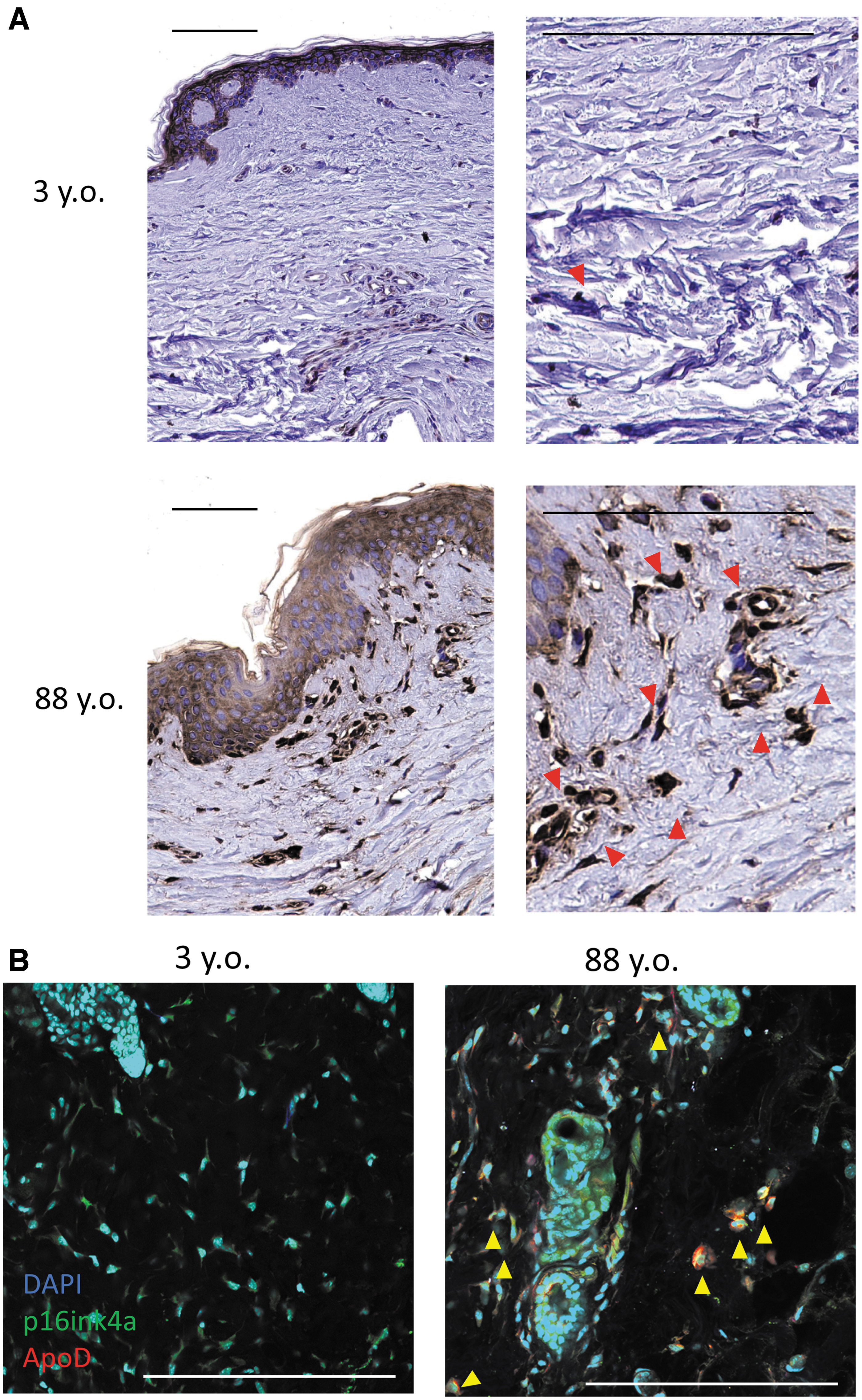
Expression of ApoD in the human dermis.
Selective targeting of senescent cells using anti-ApoD antibody
Using FACS, we analyzed whether ApoD can selectively label senescent cells in the cell population. In dermal fibroblasts, 56.22% of cells in the ReS (PDL50) population and 58.5% of cells in the RaS population were ApoD-positive, but very few were positive for ApoD in the proliferating cells (PDL10), indicating significant differences between the two groups (ReS vs. young; p = 0.0000022, RaS vs. young; p = 0.0000017) (Fig. 5A, B). Hence, ApoD may be a potential candidate marker for specific labeling of aging dermal cells in vitro.
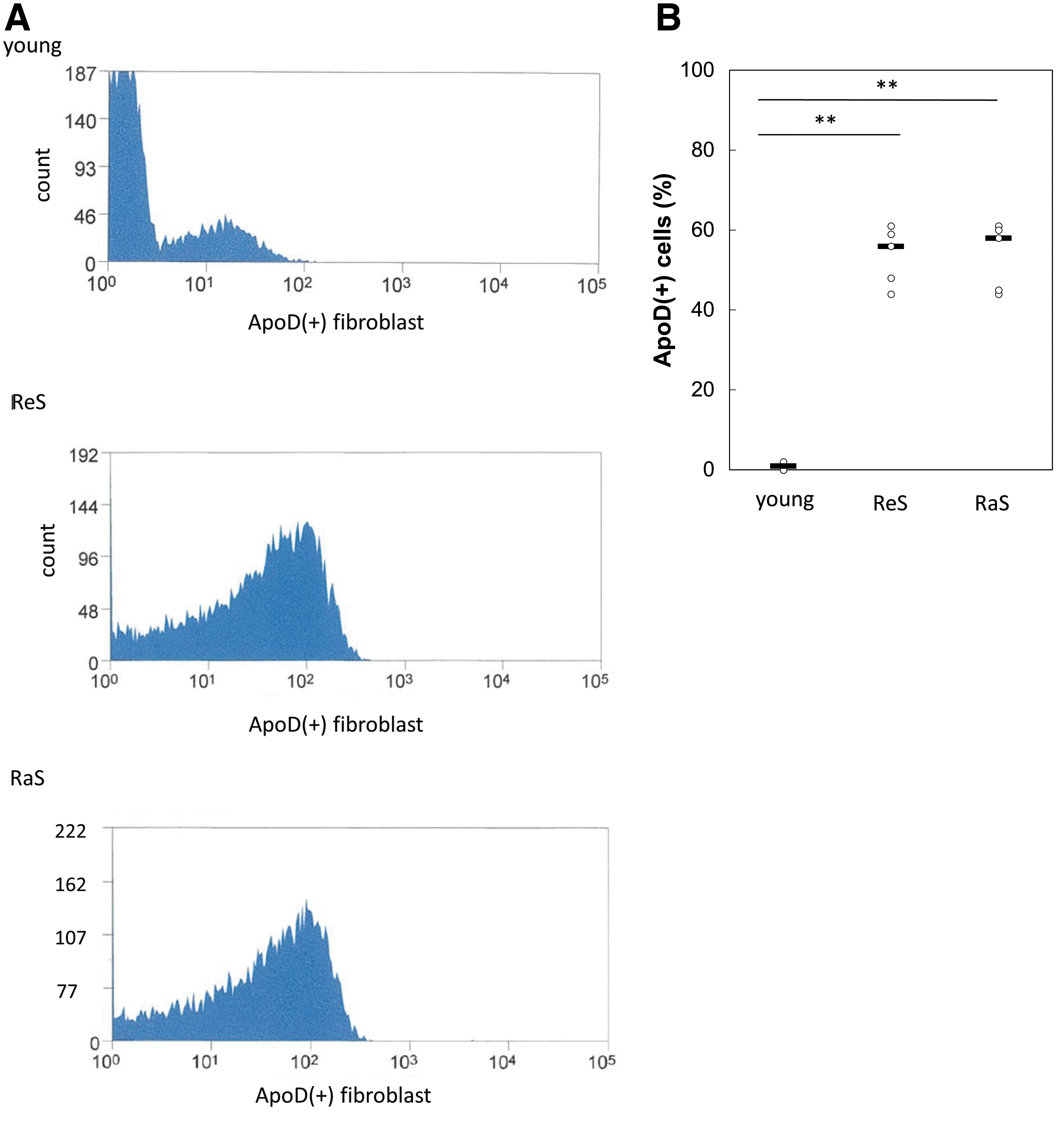
Sorting of senescent cells using an anti-ApoD antibody.
Discussion
Senescent skin contains ∼15% to 60% of senescent fibroblasts 26,27 ; these senescent cells accumulate primarily in the dermis and subcutaneous adipose tissue depots. 28 An emerging hypothesis regarding skin aging is that the secretion of SASP factors by senescent fibroblasts in the dermis causes abnormal tissue remodeling and extracellular matrix dysfunction that results in an altered collagen composition, elastin destruction, inflammatory cell infiltration, fibrosis, and adipose tissue atrophy. 29,30 Senescent cells have emerged as a novel target in antiaging therapies to counteract these processes.
ApoD, which we have now identified as a candidate marker for senescent dermal fibroblasts, has previously been reported to be expressed following induction by stress conditions such as oxidative stress, inflammatory stress, or UV treatment. 31 The nuclear factors poly(ADP-ribose) polymerase 1 and apurinic/apyrimidinic endodeoxyribonuclease 1, which are induced in pathological conditions such as oxidative stress and neurodegeneration, are upregulated in growth-arrested cells, bind to the ApoD promoter, and are capable of inducing ApoD expression. 32 –34 These factors potentially modulate ApoD in the context of senescence, allowing ApoD to function as a marker for dermal fibroblasts whose proliferation has been arrested by artificial DNA-damaging processes, such as continuous replication or irradiation.
However, whether the upregulation of ApoD observed in senescent cells causes senescence or is a consequence of senescence-associated processes remains open to debate. Previous reports have clearly demonstrated a highly conserved neuroprotective and antioxidant function for ApoD during aging. For example, a study has shown that the overexpression of the ApoD orthologue in Drosophila, GLaz, enhanced oxidative stress tolerance and prolonged life span. 35 Overexpression of human ApoD also leads to a decrease in lipid peroxidation associated with aging, accompanied by an increased stress tolerance and life span. 36 Transgenic mice overexpressing human ApoD in the brain underwent oxidative stress, exhibiting a reduced rate of lipid peroxidation and increased survival; conversely, a loss of ApoD function results in increased lipid peroxide levels in mouse brains, alongside an increased sensitivity to oxidative stress. 37 Indeed, a meta-analysis of aging-related gene expression profiles across 27 data sets, including those from mice, rats, and humans, found that ApoD is one of the most important common signatures of aging. 38 This study also hypothesizes that the age-associated upregulation of ApoD may be a response to the aging process rather than a detrimental mechanism associated with the physiological decline characteristic of aging.
In summary, the upregulation of ApoD is evidently a part of an evolutionarily conserved stress response pathway in aging organisms. Based on its reported antioxidant and anti-inflammatory effects, ApoD appears to protect against age-related oxidative stress, which accumulates through elevated reactive oxygen species levels. In other words, ApoD should be used primarily as a marker for senescent cells, not as a target for senescence inhibition.
To date, therapies that selectively destroy senescent cells include the senescent-cell-depleting drugs ABT263 and ABT737, 13,39 as well as dasatinib and quercetin 40 ; each has significant side effects. Strategies that target antigens that are specifically expressed in senescent cells include the induction of antibody-dependent cytotoxicity with dipeptidyl peptidase 4 labeling 14 and T cell therapy with urokinase-type plasminogen activator receptor 15 ; however, these methods have not been applied to human skin. Taking these strategies into account, ApoD as a targeted marker of aging could complement interventions aimed at eliminating senescent cells. Importantly, senescent cell populations are heterogeneous and express different markers in different tissues and cell types, 41 but the present study has shown that ApoD was upregulated in dermal fibroblasts, contemporaneous with the expression of several SASP factors. Such a universal marker has not been reported previously and may be an important antigen in the development of skin rejuvenation therapies that target senescent cells.
The mechanism by which the modulation of ApoD gene expression affects the senescence phenotype remains unclear in this study, and siRNA-mediated knockdown of ApoD did not affect the SASP in senescent human dermal fibroblasts. To overcome the limitations of the current study, future research should examine the effects of ApoD on skin maintenance in vivo using transgenic mice in which ApoD expression in the skin dermis is knocked out. Our current data also preclude us from drawing conclusions regarding the markers for aging in other types of cells that make up the skin, such as macrophages, or fibroblasts in other tissues. Regarding keratinocytes, previous studies have reported that ApoD increases with aging. 42 If ApoD is found to be useful as a marker for aging in other cells, it may contribute to the development of antiaging therapies for other organs.
Conclusions
In conclusion, we discovered that ApoD was strongly upregulated in human dermal senescent cells. Furthermore, senescent cells were selectively targeted by anti-ApoD antibodies in this study, providing a strong basis for the development of therapies to eliminate senescent cells in the skin.
Footnotes
Acknowledgments
The authors would like to thank all the laboratory members for their technical assistance with the experiments.
Data Sharing Statement
The data presented in this study are available upon request from the corresponding author.
Authors' Contributions
Conceptualization: K.T. and K.K. Investigation: K.T. Animal work: K.T. and K.K. Data analysis and interpretation: K.T. Writing—original draft: K.T. Supervision: T.A. and K.K. All authors have read and approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by Japan Society for the Promotion of Science KAKENHI, grant number JP 22K19589.