
Abstract
Intestinal epithelial cellular senescence contributes to the physiological decline of intestine and induces age-associated intestinal diseases. Therefore, the intestine is a vital target to delay intestinal epithelial cellular senescence and extend healthy lifespan. Alginate oligosaccharides (AOSs) have a wide range of biological and pharmacological activities. However, there are no related reports of AOSs on intestinal epithelial cellular senescence. Our study aimed to investigate the effect of AOSs on hydrogen peroxide (H2O2)-induced senescent intestinal epithelial cells (IEC-6) and its antiaging mechanism. A senescent model was successfully constructed by H2O2 (200 μmol/L) treatment on IEC-6 for 4 hours. Different concentrations of AOSs (10, 50, 100 μg/mL) were used to intervene in H2O2-induced senescent IEC-6. The number of β-galactosidase staining-positive cells was significantly reduced by AOS intervention. The expression levels of p21 and p16, known as the senescent biomarkers, were also decreased. In addition, AOSs alleviated oxidative stress by reducing reactive oxygen species and improving antioxidative ability. To understand how AOSs rejuvenate H2O2-induced senescent IEC-6, we detected the expression level of genes in autophagy process. The results indicated that AOSs restored the expression level of Beclin 1, Atg7, and LC3 to enhance autophagy process by activating activated protein kinase (AMPK) and inhibiting mammalian target of rapamycin in H2O2-induced senescent IEC-6. Compound C, an AMPK inhibitor, abolished the effect of AOSs on activating autophagy and rejuvenating senescent IEC-6. Altogether, our study suggests that AOS is a promising drug for delaying intestinal epithelial cellular senescence.
Introduction
The elderly population is growing rapidly, which will bring tremendous pressure and severe challenges to the medical system. Studies have demonstrated that the intestinal epithelial cellular senescence is highly associated with age-associated intestinal diseases. Therefore, the intestine is a vital target for the organism to extend its healthy lifespan in the elderly population. 1
Autophagy is a lysosome-mediated catabolic mechanism that maintains intracellular environmental homeostasis by degrading damaged macromolecules and organelles within the cell. 2 Studies have shown that oxidative stress is one of the critical factors to induce senescence. Upregulating the level of cellular autophagy can reduce intracellular oxidative stress and delay cellular senescence. Autophagy maintains intestinal homeostasis, improves intestinal regenerative functions, and alleviates cytosolic stress, mainly through the mammalian target of rapamycin (mTOR) pathway. 3
Fucoidan is a natural algal polysaccharide extracted from brown algae. 4 Alginate oligosaccharides (AOSs) are produced by depolymerization of alginate and carry unique advantages such as water solubility, nontoxicity, nonimmunogenicity, and biodegradability. 5 This oligosaccharide possesses neuroprotection, antioxidation, antihyperlipidemia, anti-inflammation, antiplatelet aggregation, antitumor, antibiosis, and immunomodulation. 6 –8 To elucidate this significant interaction, we speculate whether the amelioration of hydrogen peroxide (H2O2)-induced senescence by AOSs is mediated through autophagy in intestinal epithelial cells (IEC-6).
As an important physiological energy sensor, adenosine monophosphate- activated protein kinase (AMPK) is a key regulator of cellular and organismal energy homeostasis. AMPK participates in multiple metabolic pathways, balance energy supply and requirements, and eventually regulates the growth of cells and organs. mTOR is a central regulator of proliferation, cell growth, and autophagy processes. 9 mTOR is an essential downstream signaling molecule of AMPK and plays a negative role in regulating autophagy.
Previous studies have found that AOS delays cardiomyocyte aging by activating autophagy and antioxidative stress. 10 We also found that this oligosaccharide can treat Alzheimer's disease by activating autophagy. Mannuronate oligosaccharides can inhibit Alzheimer's disease through the microbiota–gut–brain axis in neurodegeneration based on findings presented in previous reports. 11 Our previous study showed that AOSs could play an important role in age-related diseases such as aging of kidney organs, osteoporosis, and cataract through activating autophagy, inhibiting oxidative stress, and improving mitochondrial function. 12 However, there are no studies related to the role of AOSs for repairing the intestinal mucosal barrier and delaying intestinal epithelial cellular senescence.
In this study, we characterized H2O2-induced IEC-6 approached senescence along with increased markers of aging. We found AOSs rejuvenated H2O2-induced senescent IEC-6 by activating autophagy through the AMPK/mTOR signaling pathway. An AMPK inhibitor abrogated AOS functions on delaying H2O2-induced IEC-6 senescence. This study will provide a new promising treatment for age-associated intestinal diseases caused by intestinal cellular senescence, such as senile inflammatory bowel disease and senile intestinal tumors.
Materials and Methods
Cell culture
IEC-6 (ATCC, no. CRL1592) were cultured as described previously (Chang et al., 2013). Cells were incubated at 37°C with 5% CO2. Cells were treated with AOSs, whereas control cells received the same volume of dimethyl sulfoxide (DMSO) (Solarbio, D8371). The final concentration of DMSO was <0.1% in all experiments.
H2O2-induced senescence model and AOS treatments
First, IEC-6 were incubated for 24 hours in DMEM (Hyclone, Logan, UT) containing different concentrations of H2O2 (50–200 μM). Then the optimized concentration (200 μM) was selected to build a cellular senescence model according to the references and our previous data. When the confluency of cells was ∼50%, IEC-6 were administrated by H2O2 for 4 hours, then other processing for 48 hours, then IEC-6 were collected for the follow experiments.
The H2O2 group was added to 200 μM H2O2 for 4 hours to establish cellular senescence model. After 4 hours, the completely fresh medium replaced the H2O2 medium for 48 hours. The H2O2+AOS groups were added to 200 μM H2O2 for 4 hours. Then H2O2 was removed, simultaneously AOS was added for 48 hours. The H2O2 (200 μM)+reactive oxygen species (ROS) inhibitor group cells were treated with 200 μM H2O2 for 4 hours. Then H2O2 was removed, simultaneously the ROS inhibitor, N-Acetyl-
The H2O2 (200 μM)+AOS(H)+autophagy inhibitor (Wortmannin) group cells were treated with 200 μM H2O2 for 4 hours. Then H2O2 was removed, simultaneously the AOS(H)+autophagy inhibitor were added for 1 hour. After 1 hour of the AOS(H)+autophagy inhibitor administration, the AOS(H) medium replaced the AOS(H)+autophagy inhibitor medium for 47 hours. The H2O2 (200 μM)+AOS(H)+AMPK inhibitor (compound C) group cells were treated with 200 μM H2O2 for 4 hours.
Then H2O2 was removed, simultaneously the AOS(H)+AMPK inhibitor were added for 2 hours. After 2 hours of the AOS(H)+AMPK inhibitor administration, the AOS(H) medium replaced the AOS(H)+AMPK inhibitor medium for 46 hours. The control group was given synchronous replaced fresh medium. No treatment with H2O2, AOS(H),AMPK inhibitors, and only the fresh medium was changed during drug treatment in the H2O2, AOS(H), AMPK inhibitors treatment group. The efficiency of compound C to inhibit AMPK has been demonstrated in the study of Wan et al. 13 AOS was obtained from Qingdao BZ Oligo Biotech Co. Dorsomorphin (compound C) (Synonyms: compound C; BML-275) was purchased from Med Chem Express. Ltd. (Qingdao, China). Thirty percent H2O2 solution was purchased from Tianjin Guangcheng Reagent Company. All other chemicals and reagents were acquired from standard commercial suppliers if not otherwise mentioned.
Senescence-associated beta-galactosidase activity assay
Senescence-associated β-galactosidase (SA-β-Gal) is a widely used marker for detecting cellular senescence. SA-β-gal activity was measured according to the manufacturer's protocol. In brief, IEC-6 were washed three times in phosphate-buffered saline (PBS), fixed for 15 minutes at room temperature by the fixative solution, and incubated overnight at 37°C with fresh SA-β-gal stain solution at pH 6.0 (Thermo Fisher Scientific K145501). The percentage of SA-β-gal was calculated by counting the positively stained cells within a sample of 200 cells ( × 100 magnification).
Lipid peroxidation measurement
As the major secondary oxidation products of lipid peroxidation, intracellular malondialdehyde (MDA) content was detected by thiobarbituric acid colorimetric assay. MDA reacts with thiobarbituric acid to produce red production. The red production can be excited at 535 nm to produce a maximum emission wavelength of 553 nm, which was detected by the fluorescence. The protocol was conducted according to the instruction of MDA Assay kit (Jiancheng Bioengineering Institute).
Assessment of superoxide dismutase activity
Assessment of superoxide dismutase (SOD). After appropriate treatments, IEC-6 were washed thrice with PBS and centrifuged at 94 g for 5 minutes. The levels of SOD were measured using Total Superoxide Dismutase Assay Kit with WST-8 (Shanghai Beyotime Biotech). The activity of SOD content by Total Superoxide Dismutase Assay Kit with WST-8. The sample was mixed with WST-8/ enzyme solution and the reaction trigger working solution, then incubated at 37 °C for 30 min. Absorbance was measured at 450 nm using a multimode enzyme marker (BioTek, USA). SOD activity is expressed as U/mg protein.
Cellular ROS detection
Cellular ROS was detected using an ROS detection kit (S0033, Beyotime) as per the instructions. In brief, 2′,7-dichloro-dihydro fluorescein diacetate (DCFH-DA) was diluted with DMEM to a final concentration of 10 μmol/L. The cells were collected and suspended in diluted DCFH-DA at a 1 × 106 cells/mL concentration and incubated in a 37°C cell incubator for 20 minutes. Cells were washed three times with MEM to remove DCFH-DA that did not penetrate into the cells. The ROS was detected by flow cytometry.
Enzyme-linked immunosorbent assay
The 1 × 109/L cell suspension was inoculated into a 12-well plate with 2 × 105 cells per well. The cell cultures were treated according to the experimental groups and aspirated, and the levels of interleukin-6 were measured according to the instructions of enzyme-linked immunosorbent assay kits (Beijing BioRab Technology Co. Ltd. ZN2880). Parallel experiments were repeated three times.
Western blot analysis
The kit used to extract cytoplasmic proteins was provided by Kangwei Century Bioengineering Institute (Beijing, China). After quantification by the Bradford method, protein extracts (30 μg) were resuspended in Laemmli Sample Buffer (Bio-Rad). The protein extracts were separated by sodium dodecyl-sulfate polyacrylamide gel electrophoresis on a 4%–20% Ready Gel® Tris-HCl Gel (Bio-Rad Laboratories, Inc., no. 1611177) and then transferred to Immun-Blot® polyvinylidene fluoride (PVDF) membrane (Bio-Rad Laboratories, Inc., no. 1620174). Then the PVDF membranes were incubated with QuickBlock blocking buffer at room temperature for 0.5 hour.
After that, PVDF membranes were incubated with associated antibodies such as AMPK (1:1000; Santa; sc-398861), p-AMPK (1:1000; Abcam; ab133448), mTOR (1:1000; Santa; sc-517464), p-mTOR (1:1000; Santa; sc-293089), LC3-II (1:1000; Abcam; ab192890), Beclin1 (1:1000; Abcam; ab302669), p21 (1:1000; Abcam; ab109199), p16 (1:1000; Abcam; ab51243), Atg7 (1:1000; Wuhan, China, 10088-2-AP), and β-actin (1:1000, 81115-1-RR; Wuhan, China), overnight at 4°C. Then the membranes were washed with tris buffered saline with Tween-20 for 0.5 hour and incubated with horseradish peroxidase-conjugated secondary antibody in primary buffer (1: 5000; Elabscience Biotechnology, China).
The membranes were treated with enhanced chemiluminescence reagents (Millipore) for 1 minute and exposed to a chemical luminescent imager (Viber Lourmat, France). Finally, the blots were scanned and analyzed with Quantity One 1-D Analysis Software (Bio-Rad Laboratories, Inc.).
Quantitative real-time polymerase chain reaction
The Trizol total RNA extraction method was used for extracting total RNA from IEC-6. One microgram of total RNA was converted to cDNA using PrimeScript™ II 1st Strand cDNA Synthesis Kit (Takara Biomedical Technology (Beijing) Co., Ltd.; no. 6210A). The polymerase chain reaction (PCR) primer sequences were designed according to the gene sequences reported in GenBank (Table 1) and were chemically synthesized. Each sample was tested in triplicate, with the use of the quantitative SYBR Green PCR kit (VAZYME, Nanjing, China) for 40 cycles (50°C for 2 minutes, 95°C for 10 minutes, 95°C for 30 seconds, and 60°C for 30 seconds) on the ABI 7900HT fast real-time PCR System (Applied Biosystems, Foster, CA).
Primer Sets Used for Real-Time Quantitative Polymerase Chain Reaction
Statistical analysis
SPSS 17.0 statistical software was used for data analysis. The data are expressed as mean ± standard deviation (SD). The student's t-test was used for comparison between two groups. One-way analysis of variance (ANOVA) was used when more than two groups were compared, and Tukey's post hoc test was used for pairwise comparisons after one-way ANOVA. p < 0.05 was considered statistically significant.
Results
H2O2 induces cellular senescence for IEC-6
To determine AOS functions, we first generated H2O2-induced senescent IEC-6. In brief, IEC-6 were treated by 50μM, 100μM, and 200μM H2O2 for 4 hours, then we conducted the experiments after 4 hours H2O2 treatment. IEC-6 showed a dose-dependent increase of ROS and SA-β-gal signals (Fig. 1A, B). MDA, a marker of oxidative stress, was measured through thiobarbituric acid colorimetric assay and drastically increased in IEC-6 at the 200 μM H2O2 treatment group (Fig. 1C). Simultaneously, the activity of SOD, an antioxidant enzyme, was depleted by H2O2 treatment (Fig. 1D).
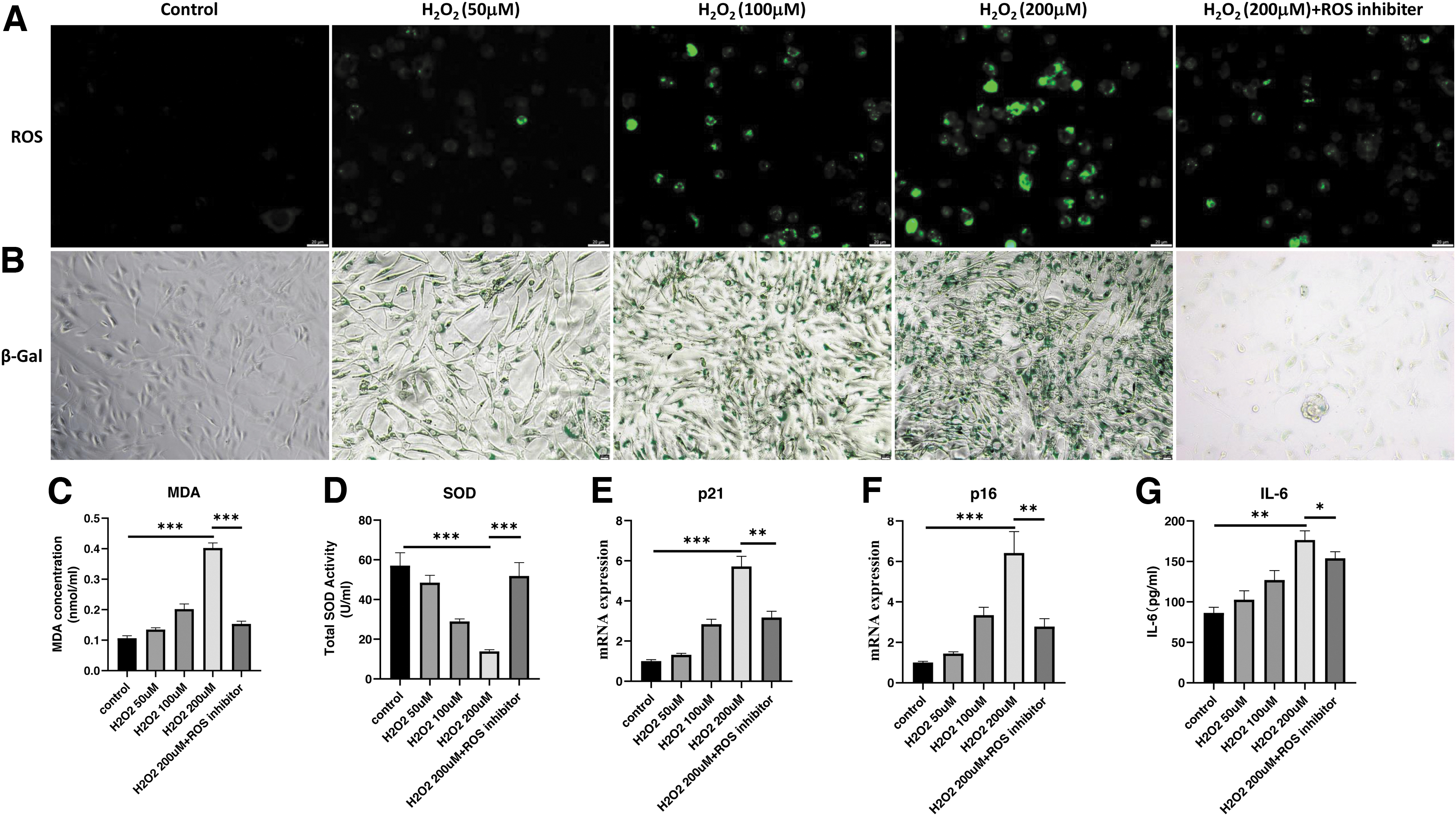
H2O2 leads to a dose-dependent induction of elevated ROS levels and senescence in IEC-6. IEC-6 were treated with 50, 100, and 200 μM H2O2 for 4 hours to induce cellular senescence, and an additional group of ROS inhibitor (2 mM NAC treatment for 6 hours) was added as a control.
Quantitative real-time PCR (RT-qPCR) experiments revealed that H2O2 treatment significantly increased the expression levels of senescent markers including p21 and p16, especially at 200 μM H2O2 concentration (Fig. 1E, F). ROS inhibitor, NAC, decreased oxidative stresses, restored SOD activity, and rescued H2O2-induced senescent phenotypes (Fig. 1). H2O2 successfully induced the senescent phenotypes for IEC-6.
AOSs reduce ROS accumulation and ameliorate markers of cellular senescence in H2O2-induced senescent IEC-6
To investigate the function of AOSs on H2O2-induced IEC-6 senescence, IEC-6 were treated with 200 μM H2O2 for 4 hours, then H2O2 was removed, simultaneously AOSs were added at AOS(L), 10 μg/mL; AOS(M), 50 ug/mL; and AOS(M), 100 μg/mL for 48 hours. The experiments covered six groups (Control, H2O2, H2O2+AOS(L), H2O2+AOS(M), H2O2+AOS(H), H2O2+ROS inhibitor). AOSs administration significantly alleviated ROS, SA-β-gal signals, and MDA levels at the H2O2+AOS(H) group compared with the H2O2 group (Fig. 2A–C).
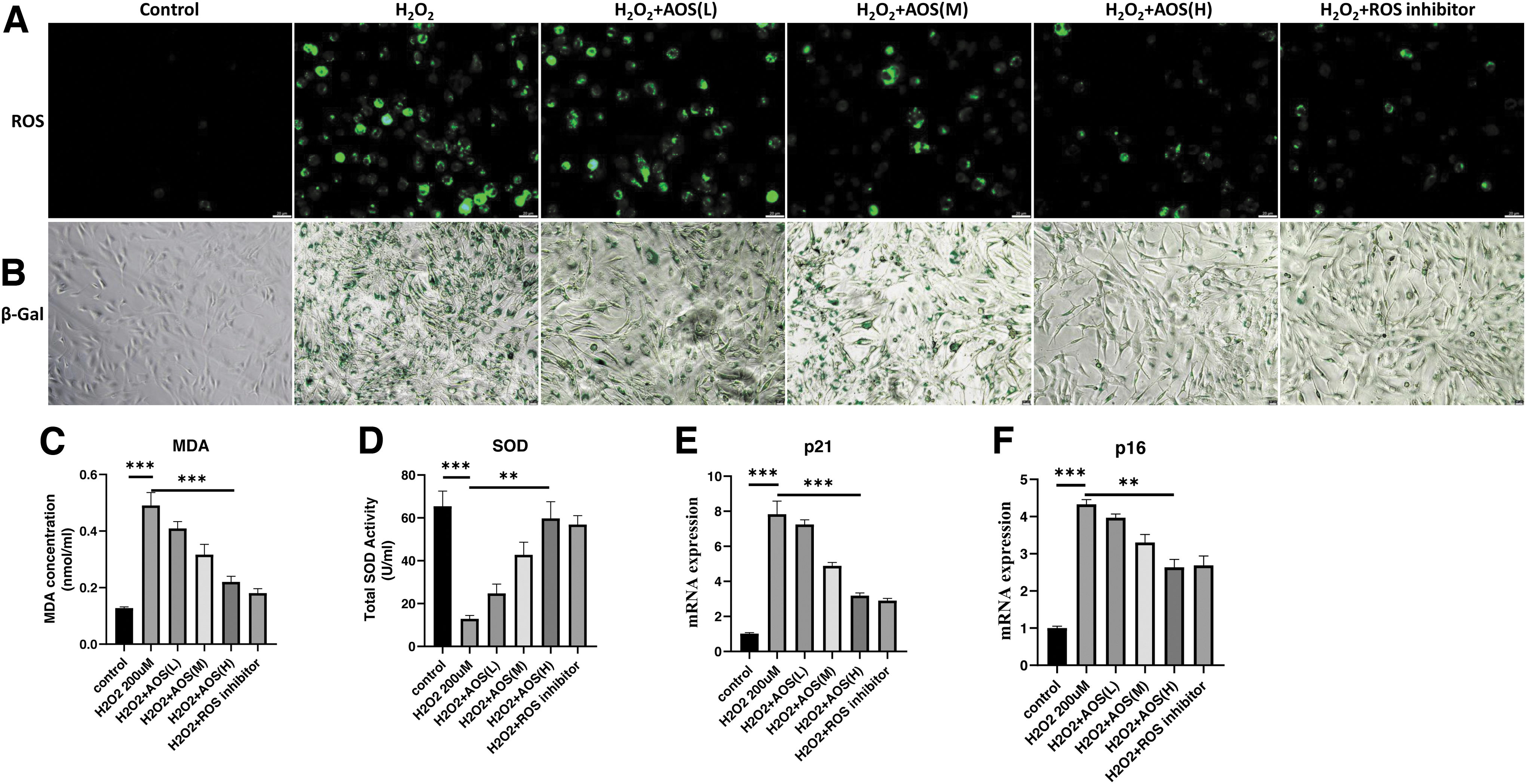
AOS concentration-dependent reduction of cellular ROS levels and senescence induced by H2O2. IEC-6 were treated with 200 μM H2O2 for 4 hours and AOSs at concentrations of AOS(L)-10 μg/mL, AOS(M)-50 μg/mL and AOS(H)-100 μg/mL were added to continue treatment for 48 hours. An additional group of ROS scavenger group (2 mM NAC treatment for 6 hours) was added as control.
In addition, AOS(H) restored SOD activity approaching the level of Control group (Fig. 2D). AOS(H) treatment also significantly reduced the expression levels of senescent markers in H2O2-induced senescent IEC-6, such as p21 and p16 (Fig. 2E, F). The rejuvenating effect of AOSs for H2O2-induced senescent IEC-6 was dose dependent (Fig. 2). The data revealed that AOS(H) was the optimal concentration to rejuvenate H2O2-induced senescent IEC-6. Taken together, these results showed that the senescent cellular model of IEC-6 was established by H2O2 induction. We found AOS administration rescued the H2O2-induced senescence of IEC-6, and its antiaging effect was dose dependent.
AOSs rescue autophagy in H2O2-induced senescent IEC-6
Autophagy is an essential catabolic process in eukaryotic cells, which maintains cell survival, development, and cellular homeostasis. To assess the effect of AOSs on autophagy in H2O2-induced senescent IEC-6, we first induced autophagy by rapamycin and measured autophagy markers, Beclin1, Atg7, and LC3 by RT-qPCR. As shown in Fig. 3A, the control group showed that rapamycin strongly enhanced cellular autophagy. H2O2 treatment abolished autophagy in IEC-6.
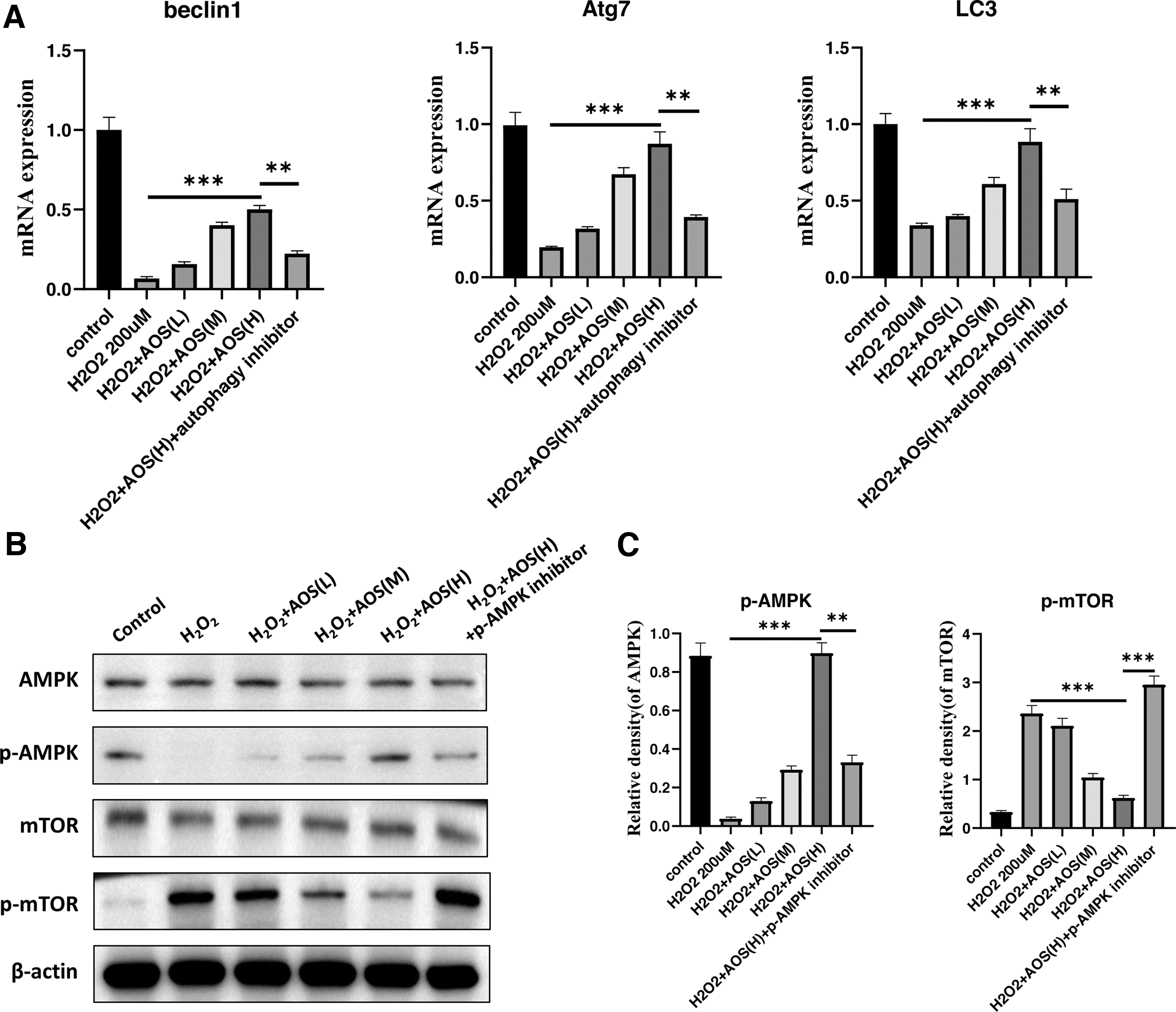
AOS concentration-dependent enhancement of autophagy levels in IEC-6 cells and inhibition of cellular senescence induced by H2O2 through autophagy. IEC-6 were treated with 200 μM H2O2 for 4 hours after induction of autophagy by rapamycin (1 μM treatment for 24 hours), and continued treatment with 10, 50, and 100 μg/mL concentrations of AOSs for 48 hours. An additional autophagy inhibitor group (500 nM Wortmannin treatment for 1 hour) was added as a control.
We examined Beclin1, Atg7, and LC3 in AOS-treated IEC-6 and observed an obvious restoration of autophagy, especially in the AOS(H) group. Autophagy inhibitor abrogated the AOS effect on activating autophagy (Fig. 3A). Here we demonstrated that AOS rescued the expression of autophagy protein Beclin1, Atg7, and LC3 in H2O2-induced senescent IEC-6 to enhance the autophagy process.
Previous studies reported that the AMPK/mTOR signaling pathway, as one of the key pathways in the regulation of autophagy, executed an important role. 14,15 Therefore, we asked whether AOSs promoted autophagy by the AMPK/mTOR pathway in H2O2-induced senescent IEC-6. To test this idea, we first examined p-AMPK/AMPK and p-mTOR/mTOR by Western Blot. The results showed that H2O2 treatment inhibited AMPK phosphorylation and enhanced mTOR phosphorylation compared with the control group.
AOSs promoted the phosphorylation and activity of AMPK, whereas decreasing mTOR activation by inhibited mTOR phosphorylation in H2O2-induced senescent IEC-6 (Fig. 3B, C). Moreover, the pharmacological inhibitor of AMPK, compound C, abolished effect of AOSs on AMPK and mTOR.
AOSs activate autophagy through regulating AMPK/mTOR signaling
AMPK activity and mTOR inhibition after AOSs treatment may induce autophagy activity. We examined the mRNA and protein levels of autophagy markers. The expression levels of Beclin1, Atg7, and LC3 were rescued by AOSs treatment in H2O2-induced senescent IEC-6. AMPK inhibitor abolished AOSs effect on autophagy markers (Fig. 4A–C). In addition, AMPK inhibitor increased senescent markers, such as p16 and p21, to remove AOSs antiaging effect in H2O2-induced senescent IEC-6 (Fig. 4 D, E). Collectively, AOSs rejuvenated H2O2-induced senescent IEC-6 by activating autophagy through the AMPK/mTOR signaling pathway.
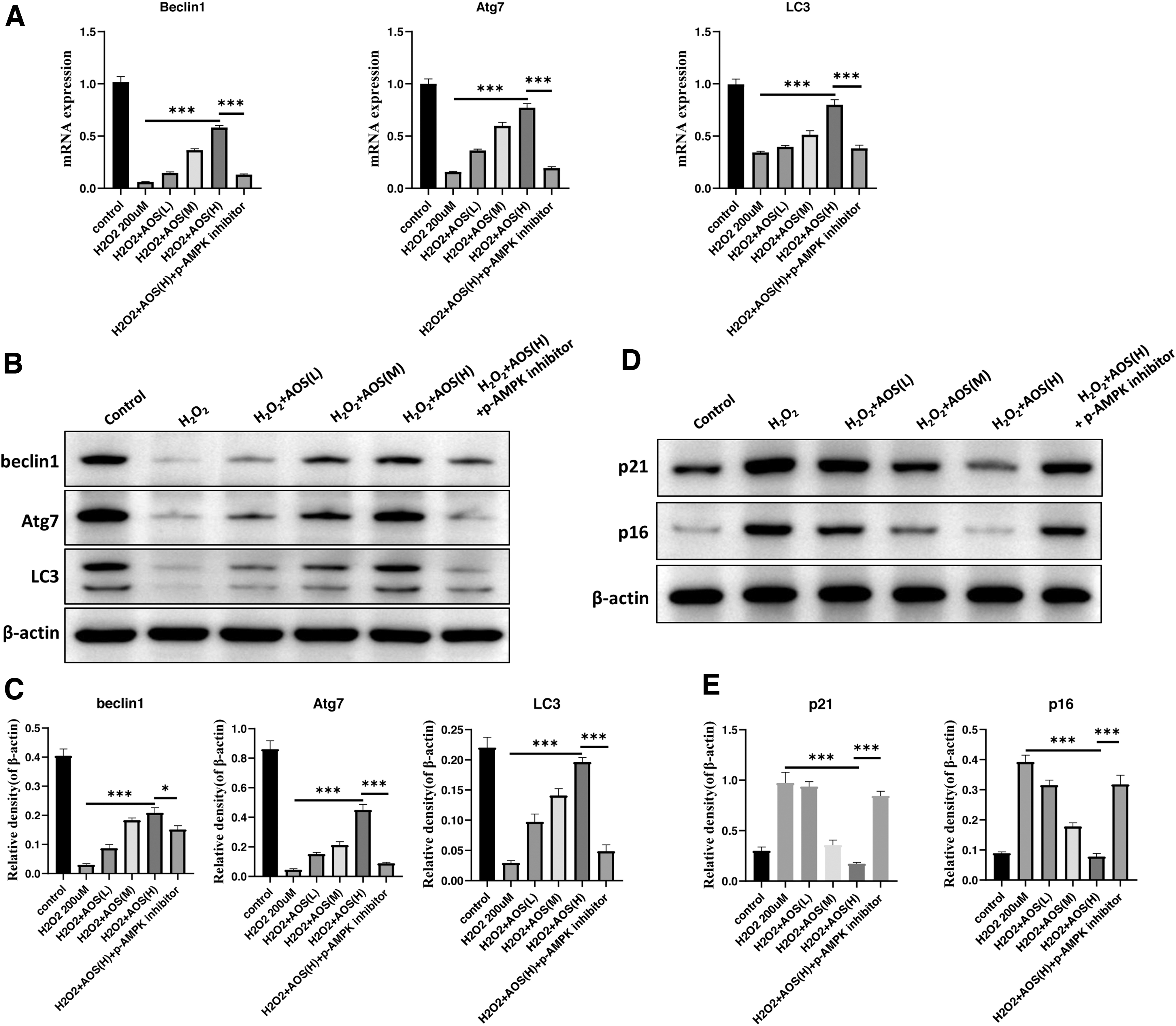
AOS enhances the level of autophagy in IEC-6 through a concentration gradient of AMPK/mTOR signaling pathway, thereby inhibiting cellular senescence induced by H2O2. IEC-6 were treated with 200 μM H2O2 for 4 hours after induction of autophagy by rapamycin (1 μM treatment for 24 hours), and 10, 50, and 100 μg/mL concentrations of AOSs were added to continue treatment for 48 hours. An additional group of p-AMPK inhibitor group (0.5 μM compound C treatment for 1 hour) was added as control.
Discussion
Aging is a chronic multiorgan-related systemic alteration in the organism. The intestinal system manifests degenerative changes presenting morphological alteration, decreasing secretion and absorption, and reducing motility. 16 Through the release of proinflammatory factors and the production of oxidative stress, the intestinal epithelial cells undergo apoptosis or pyroptosis, leading to the decline of intestinal immune function and the aging of the human organism. 17 Therefore, it is vital to investigate a novel antiaging intervention to improve intestinal function and delay aging-induced digestive diseases in the elderly population.
Alginate is a linear copolymer of polysaccharides extracted from brown seaweed. AOS, derived from alginate hydrolysis, has received increasing attention due to its low molecular weight and viscosity. It also has been widely used in several fields because of its nontoxicity, biodegradability, and less to induce immune responses. 11 In recent years, antioxidant function of AOSs has received particular attention for age-related diseases. It is used to treat neurodegenerative diseases, delay cardiac and renal aging, and improve osteoporosis cataracts in mice. 18 However, there are no studies that focus on the role of AOSs in intestinal epithelial cellular senescence. 19
Oxidative damage is considered to be one of the significant causes of cellular senescence. Excessive production of endogenous ROS impaired mitochondrial homeostasis and induced cellular senescence along with aging. 20,21 A single layer of epithelial cells confers the first line of defense of the intestine. 22 In addition to their role in nutrient absorption and metabolism, intestinal epithelial cells establish a mucosal barrier that protects the body from exogenous factors, including pathogenic bacteria. In this study, we found AOSs not only reduced ROS stress, but also mitigated the senescent phenotype, such as SA-β-gal signals and aging markers shown in Fig. 2.
Feng et al. found that AOSs significantly promoted the expression of Beclin 1, a signature protein of autophagy in cardiomyocytes, in a dose-dependent manner by inhibiting mTOR phosphorylation. 23 Beclin 1, Atg7, and LC3 can be used as markers to detect autophagy levels. 24 Autophagy activity declined with aging in humans, resulting in a considerable accumulation of harmful substances in the cytoplasm, promoting the development of aging-related diseases. Studies have shown that increasing the level of autophagy can delay aging and have a particularly protective effect on aging-related diseases. 25 In this study, we observed a clear reduction of Beclin 1, Atg7, and LC3 in H2O2-induced senescent IEC-6 (Fig. 3). AOSs restored the expression of Beclin 1, Atg7, and LC3 (Fig. 3).
Multiple lines of evidence suggest a critical link between intestinal function and autophagy. Autophagy function and its regulatory mechanisms are involved in intestinal homeostasis and repair ability, supporting intestinal barrier integrity in response to cellular stress through tight junctions. 26 AMPK plays an essential role in regulating intracellular energy balance and maintaining cellular homeostasis. AMPK is activated and transformed into p-AMPK by activating autophagy and inhibiting the activation of mTOR. 27
Therefore, we examined the effect of AOSs on the AMPK/mTOR pathway to clarify the potential mechanisms that AOS regulates autophagy in H2O2-induced senescent IEC-6. We found that H2O2 treatment inhibited intracellular AMPK phosphorylation and enhanced mTOR phosphorylation. Whereas AOSs restored AMPK phosphorylation and reduced mTOR phosphorylation in H2O2-induced senescent IEC-6 (Fig. 3B, C). Therefore, we hypothesized that AOSs enhanced autophagy activity in H2O2-induced senescent IEC-6 through the AMPK/mTOR signaling pathway.
Next, we conducted AMPK inhibitor experiment and found AMPK inhibitor abolished AOSs effect on enhancing autophagy activity (Fig. 4A–C). In addition, AMPK inhibitor accelerated senescent phenotype and abrogated AOSs antiaging function (Fig. 4D, E).
This study elucidated that AOSs rejuvenated intestinal epithelial cellular senescence by activating autophagy in intestinal epithelial cells through regulating the AMPK/mTOR signaling pathway. Consistent with the mentioned findings, the AMPK inhibitor experiment attenuated the antiaging effect of AOSs. Taken together, our results tentatively confirm that AOS delays intestinal epithelial cellular senescence by regulating the AMPK/mTOR signaling pathway to activate the level of autophagy in intestinal epithelial cells.
The current understanding of AOS-mediated autophagy to delay aging is still in its primary stage. Until now, our study has some limitations. First, we examined the effect of AOSs on senescent IEC-6 only at the protein level. Second, further studies are needed to investigate the specific autophagic pathways triggered by AOSs. More experimental data are needed to support the application of AOSs in the treatment of digestive diseases associated with intestinal epithelial cellular senescence. Furthermore, whether AOSs can delay intestinal epithelial cellular senescence by improving the intestinal mucosal barrier through modulation of intestinal flora needs to be investigated.
In conclusion, AOS exhibits a wide range of promising biological activities with multiple pharmacological effects and may be an effective therapeutic medicine for delaying intestinal epithelial cellular senescence, preventing intestinal epithelial cellular senescence-related inflammatory bowel disease, inhibiting intestinal tumors and other common digestive diseases in the elderly population. However, in our study, we only explored the mechanism of the autophagy-related pathway to delay intestinal epithelial cellular senescence, and other mechanisms of AOSs to protect intestinal mucosal barrier and delay the intestinal epithelial cellular senescence need to be further determined. The results of this study provide a new tool for treatment of aging-related intestinal diseases.
This study suggests that AOS is a promising drug for rejuvenating intestinal epithelial cellular senescence. Its potential molecular mechanism is related to enhancing autophagy by the AMPK/mTOR signaling pathway. The results of this study provide a new tool for treatment of aging-related intestinal diseases.
Conclusions
This study suggests that AOS is a promising drug for rejuvenating intestinal epithelial cellular senescence. Its potential molecular mechanism is related to enhancing autophagy by the AMPK/mTOR signaling pathway. The results of this study provide a new tool for treatment of aging-related intestinal diseases.
Footnotes
Acknowledgments
The authors thank all of the doctors and graduate students who were involved in the study.
Authors' Contributions
X.Z. contributed to conceptualization (equal), data curation (lead), formal analysis (lead), investigation (lead), methodology (lead), project administration (equal), software (lead), validation (lead), visualization (lead), and writing—original draft (lead). S.Z. was involved in investigation (supporting), formal analysis (equal), methodology (equal), project administration (equal), resources (supporting), and software (supporting). S.H. was in charge of investigation (supporting), formal analysis (equal), methodology (supporting), project administration (equal), and resources (supporting).
P.W. took charge of investigation (supporting), formal analysis (supporting), methodology (supporting), project administration (supporting), and resources (supporting). J.M. was in charge of investigation (supporting), methodology (supporting), project administration (supporting), and resources (supporting). C.G. was in charge of software (supporting), methodology (supporting), writing—review and editing (supporting), validation (supporting), and visualization (supporting). X.Z. was in charge of funding acquisition (supporting), resources (supporting), and writing—review and editing (supporting).
Q.Z. was in charge of funding acquisition (supporting), resources (supporting), and writing—review and editing (supporting). Y.Y. was involved in conceptualization (lead), funding acquisition (lead), project administration (lead), resources (lead), supervision (lead), and writing—review and editing (lead).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by funding from the National Natural Science Foundation of China (81771498), the Project of Traditional Chinese Medicine Science and Technology Plan in Zhejiang Province (2020ZQ032), the Project of Traditional Chinese Medicine Science and Technology Plan in Zhejiang Province (2021ZQ052), the Traditional Chinese Medicine (Integrated Chinese and Western Medicine) Key Discipline Construction Project of Zhejiang Province (no. 2017-XK-A31), National Natural Science Foundation of China (2020NSFC82002013), and Zhejiang Provincial Natural Science Foundation (2019LQ20H020009).
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.