
Abstract
DNA damage represents one of the cell intrinsic causes of stem cell aging, which leads to differentiation-induced removal of damaged stem cells in skin and blood. Dietary restriction (DR) retards aging across various species, including several strains of laboratory mice. Whether, DR has the potential to ameliorate DNA damage-driven stem cell exhaustion remains incompletely understood. In this study, we show that DR strongly extends the time to hair graying in response to γ-irradiation (ionizing radiation [IR])-induced DNA damage of C57BL/6 J mice. The study shows that DR prolongs resting phase of hair follicles. DR-mediated prolongation of hair follicle stem cell (HFSC) quiescence blocks hair growth and prevents the depletion of HFSCs and ckit+ melanoblasts in response to IR. However, prolongation of HFSC quiescence also correlates with a suppression of DNA repair and cannot prevent melanoblast loss and hair graying in the long run, when hair cycling is reinitiated even after extended periods of time. Altogether, these results support a model indicating that nutrient deprivation can delay but not heal DNA damage-driven extinction of melanoblasts by stalling HFSCs in a prolonged state of quiescence coupled with inhibition of DNA repair. Disconnecting these two types of responses to DR could have the potential to delay stem cell aging.
Introduction
Hair graying is one of the most significant signs of aging. Hair is pigmented by melanin produced by melanocytes, which are generated by melanoblasts that are renewed by melanocyte stem cells (MeSCs) during proliferative stage of hair follicles (HFs). The hair bulge contains the hair follicle stem cells (HFSCs), which control hair growth and serve as the niche for MeSCs. 1,2 When HFSCs receive proliferative signals from the hair germ in late phase of resting stage, the hair bulge turns from quiescent state into proliferative state, which triggers MeSCs to proliferate. 2 In general, HFSCs and MeSCs act in concert to regenerate pigmented hair with each hair cycle. 2,3 Therefore HFSCs, MeSCs, and hair cycle represent an in vivo model to study molecular pathways that control stem cell quiescence, proliferation, and tissue maintenance as well as the possible influence of these pathways on stem cell aging—a process characterized by the accumulation of molecular damages and functional decline of stem cells.
Calorie restriction is a dietary regiment based on reduced calorie intake without malnutrition. 4 In experimental settings, calorie restriction is most often performed by 10%–30% reduction in food intake, which is known as dietary restriction (DR). DR is a nonpharmacological way to delay aging and extend life span across different species.
A body of studies have shown that DR delays and lowers onset of age-associated pathologies in mammalians, including suppression of cancer development, type 2 diabetes, hypertension, heart disease, memory loss, and hearing impairments. 4 –9 However, cellular consequences of DR at the level of adult stem cells, which sit in the central core of tissue aging remain largely unknown. It was shown that DR augments self-renewal activity of intestinal stem cells by upregulating mammalian target of rapamycin complex1 (mTORC1) signaling in Paneth cells that form a niche for intestinal stem cells. 10 DR also improved endogenous muscle repair and benefit muscle regeneration in transplantation assay through enhancing muscle stem cell activity. 11 Recently, it was shown that DR promotes extensive skin and fur remodeling associated with expansion of the HFSC pool. 12
Previously, we showed that DR significantly delays hematopoietic stem cell (HSC) aging by increasing HSC quiescence in homeostasis state and also strongly reduces their proliferation in response to stress. 13 It remains to be investigated whether DR also plays a role in regulating cell cycle of HFSCs and how this may affect the abrogation of stem cell function in response to molecular damages that promote stem cell and tissue aging, such as DNA damage.
The accumulation of DNA damage is one of the major causes of aging. 14,15 Premature hair graying can be induced by ionizing radiation (IR), which causes DNA double-strand breaks. 16 On a molecular level, it was shown that IR leads to loss of MeSCs by Atm-dependent induction of stem cell differentiation and loss of self-renewal. 16 Similarly, a Batf-dependent differentiation-inducing DNA damage checkpoint was identified in lymphoid-biased HSCs. 17 It is currently unknown whether DR would have the potential to prevent stem cell aging by abrogating the induction of DNA damage checkpoints that lead to a depletion of stem cell pools, such as the DNA damage-dependent induction of stem cell differentiation.
In this study, we investigated the effect of DR on hair graying induced by IR in C57BL/6 J mice. The study provides the first experimental evidence that adult-onset DR (30% of ad libitum [AL] diet) significantly delays hair graying after IR by promoting a significant prolongation of the resting phase of HFs for up to 3 months after initiation of DR. The study shows that DR mediates a prolonged induction of HFSC quiescence, which was sufficient to prevent damage-induced differentiation and loss of activated ckit+ MeSCs (melanoblasts).
However, DR also leads to profound inhibition in the expression of DNA repair gene in HFSCs and when hair cycle is reintroduced by refeeding (RF) or spontaneously after prolonged periods of DR, MeSCs get quickly depleted and hair graying proceeded like in freshly irradiated, AL-fed mice. These results indicate that DR has the potential to delay DNA damage-induced MeSC depletion by inducing a prolonged maintenance of HFSC quiescence. However, DR also suppresses the expression of key DNA repair pathways in HFSCs and cannot prevent DNA damage-induced stem cell depletion when cell cycle activity resumes even after prolonged periods of time.
Materials and Methods
Mice and dietary intervention
C57BL/6J mice were obtained from Hunan SJA Laboratory Animal Co. Ltd. (Hunan, China) and maintained in the animal facilities of Nanchang Royo Biotech under pathogen-free conditions on a 12-hour light/12-hour dark cycle. Mice were housed individually and received either of the following regimen: AL, fed with unlimited amount of food; DR diet, fed daily with an amount of food corresponding to 70% of the amount of food consumed by body weight- and gender-matched mice in the AL group. The provided food amount was constant over the whole DR period. All mouse experiments were approved by the Nanchang Royo Biotech Co. Ltd. (RYEI20170420-1). The number, ages, and sexes of mice used in each experiment are outlined in Table 1.
The Number, Ages, and Sexes of Mice Used in Each Experiment
Radiation
A commercial medical electronic linear accelerator (Elekta VersaHD) was used for the radiation experiments. The position of irradiated samples was set at source to surface distance 100 cm from the source of the machine. Around 20 × 20 cm2 was set for the radiation field size. The beam used was 6 MV X-ray with dose rate of 600 MU/min. The daily dose output was checked using a commercial farmer ion chamber PTW 30013, which was calibrated by secondary standard dosimetry laboratory. The prescribed doses used in this research were 6 and 12 Gy, respectively.
Histology staining
Three-micrometer paraffin sections of the skin were used for H&E staining. For immunofluorescence staining of ckit, 5-μm-thick frozen sections of skin were used. Sections were fixed with 4% PFA for 10 minutes at room temperature, and then washed with phosphate-buffered saline (PBS) followed by 1 hour blocking with 5% horse serum/1% BSA/0.3% Triton X-100 at room temperature. Then the sections were washed again and afterward incubated with primary antibody (Kit, BioLegend) diluted at a ratio of 1:100 at 4°C overnight in a humid chamber. The slides were then washed three times with PBS the next day, and incubated with secondary antibodies diluted at a ratio of 1:500 at room temperature for 1 hour. After washing with PBS, slides were mounted with mounting medium with 4′,6-diamidino-2-phenylindole (DAPI) (Vector) and covered with coverslips.
For proliferating cell nuclear antigen (PCNA) and γ-H2AX immunohistochemistry staining, 5-μm paraffin sections of skin samples were used. The sections were deparaffinized and hydrated, and then placed in an antigen retrieval solution (0.01 M citrate buffer, pH 6.0) for 15 minutes in a microwave oven at 100°C at 600 W and quenched and incubated with freshly prepared 3% H2O2 for inhibition of endogenous peroxidase.
Then the sections were washed and afterward incubated with primary antibody at 4°C overnight in a humid chamber. The slides were then washed three times with PBS the next day, and incubated with goat anti-mouse biotinylated IgG at room temperature for 1 hour. Afterward, the slides were washed and stained with diaminobenzidine and were counterstained with hematoxylin, dehydrated, and mounted. For negative control in the immunohistochemistry procedures performed, mouse 10% normal serum replaced the primary antibodies. Antibodies used were the following: PCNA (ab29; Abcam) 1:1000 diluted; γ-H2AX (Ser139 Merck) 1:500 diluted; Goat anti-mouse biotinylated IgG (ZSGB-BIO, PV-6000) 1:1000 diluted.
Tunel staining
Five-micrometer paraffin sections of the skin were used for tunel staining. The sections were deparaffinized and hydrated, and then incubated at room temperature with 20 μg/mL proteinase K for 20 minutes. The sections were washed and subsequently incubated with 3% hydrogen peroxide in absolute methanol for 5 minutes at room temperature to inhibit endogenous peroxidase. Then the sections were washed and afterward incubated in blocking buffer (10% BSA in PBS) for 10 minutes. After being rinsed in PBS, the sections were incubated in TdT equilibration buffer for 5 minutes. Slides were incubated in the TUNEL mixture for 60 minutes at 37°C afterward and washed in PBS. Then slides were mounted with mounting medium with DAPI (Vector) and covered with coverslips.
Statistics
GraphPad Prism 6.0 software was used to calculate P-values. The unpaired two-tailed Student's t-test was used for two-group datasets and one-way analysis of variance (ANOVA) or two-way ANOVA was used for multigroups datasets.
Results
DR delays IR-induced hair graying
Hair graying is one of the most conspicuous phenotypes of aging, which has been linked to DNA damage 14 –16 and telomere dysfunction. 18 Accordingly, premature hair graying can be induced by IR, which causes DNA double-strand breaks. 16,19 DR is one the most well-known regiments to delay aging processes and to increase organism lifespan. To test the effect of DR on hair graying, 7-week-old mice were irradiated with 6 Gy γ-ray and then exposed to DR or an AL diet. Visible gray hair appeared on dorsal skin of AL-fed mice around 3 weeks after irradiation. Approximately 3 months after IR, hair of AL mice had nearly, completely turned gray. In striking contrast, hair of DR mice remained black during the whole observation period of 3 months (Fig. 1A, B).
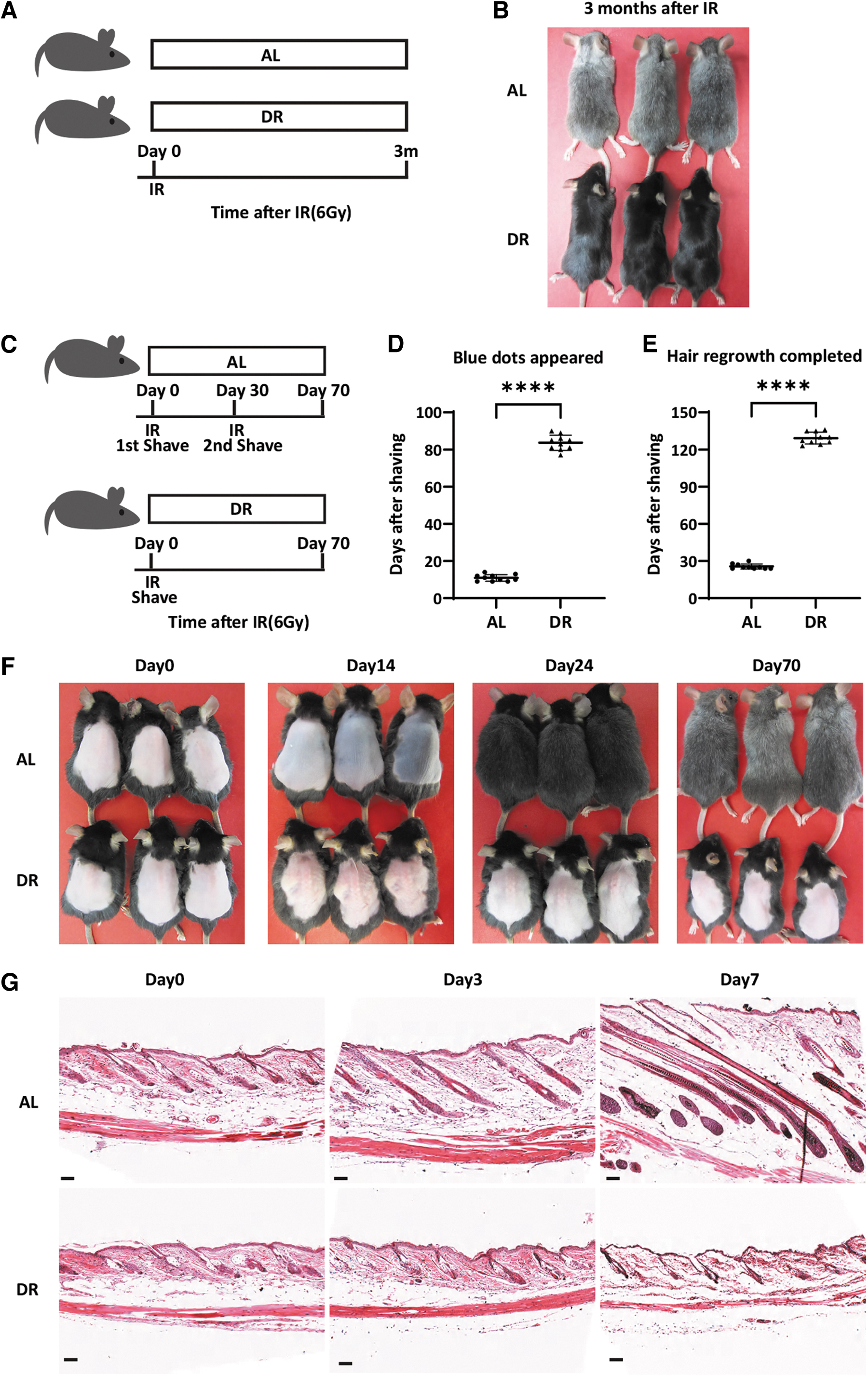
DR delays hair graying after gamma ray irradiation associating with prolonged telogen phase of HFs.
DR significantly prolongs resting phase of HFs after γ-irradiation
HFSCs and MeSCs act in concert to regenerate pigmented hair with each hair cycle. 2 The onset of hair graying is preceded by the appearance of ectopically pigmented melanocytes with a dendritic morphology in mid-anagen phase after IR, which is due to DNA damage-induced differentiation of MeSCs in the niche 16,20 with quiescent MeSCs being most sensitive to this mode of DNA damage-driven stem cell depletion. 21 DNA damage-induced MeSC depletion inevitably results in depigmentation of newly grown hair. 16 The preceding stem cell activation at early anagen or stem cell division is necessary for ectopic pigmentation of MeSCs, 16 thus representing a hallmark phenotype of DNA damage-induced differentiation, which is easily accessible and thus suitable to analyze the potential of interventions to delay DNA damage-driven stem cell and tissue aging.
DR has been shown to delay early aging of HSCs by reducing Igf-signaling and cell cycle activity. 13 To analyze whether reductions in cell cycle activity would associate with the protective effects of DR on IR-induced hair graying, we analyzed hair cycle in DR versus AL-fed mice.
To this end, dorsal hair of 7-week-old mice was removed before irradiation (Fig. 1C–F). At this stage, dorsal skin HFs were all in resting phase (telogen), as evidenced by the homogeneous pink color of the skin (Fig. 1F). Directly after irradiation, mice were exposed to DR or AL diet. In mice of the AL group, blue dots appeared on the dorsal skin 10.90 ± 1.792 days (n = 10 mice) after irradiation, which is a sign of HFs entering the anagen phase of the hair cycle, followed by gradual increases in the blue coloring of the skin of AL-fed mice on day 14 after DR (Fig. 1D, F). Twenty-four to 30 days after irradiation, the fur was fully regrown in AL-fed mice and hair graying became visible at this time point (Fig. 1E, F). At this time point, the dorsal hair of AL mice was removed again, and the dorsal skin appeared pink suggesting the HFs returned to telogen phase again.
However, the hair regrew similar to the first round of shaving and at day-70 after initiation of the experiment with IR the fur had completely regrown for a second time and at this point was almost completely gray (Fig. 1C, D). Remarkably, dorsal skin of DR mice remained pink and hair did not regrow while AL mice had completed two hair cycles (Fig. 1F). Blue dots started to appear on the dorsal skin of DR mice 83.70 ± 4.084 days (n = 10 mice) after irradiation, which was significantly delayed compared with AL mice (p < 0.0001, Unpaired two-tailed, Student's t-test). DR mice completed first round of hair regrowth 129.20 ± 4.780 days (n = 10 mice) after irradiation, which was around four times as long as the AL mice (p < 0.0001, Unpaired two-tailed, Student's t-test). These results indicated that DR significantly delayed initiation of hair cycle after irradiation.
To substantiate this finding, we analyzed the morphology of HFs on skin sections of DR versus AL-fed mice within 7 days after IR and hair shaving. In accordance with the skin pigmentation, HFs of AL mice entered anagen phase shortly after shaving, while HFs of DR mice remained in the resting phase (Fig. 1G). Altogether, these data indicated that HFs of DR-exposed mice are kept in the resting phase, whereas AL fed mice finished two rounds of hair cycles and regrowth in the same observation period (Fig. 1C–F). These results suggested that the delay in hair growth may be related to the delay in hair graying in mice that were exposed to DR compared with AL diet (Fig. 1A, B).
DR significantly prolongs resting phase of HFs under nonirradiated condition
To analyze whether DR-dependent suppression of hair cycle activity was a specific effect in irradiated mice or a general IR-independent response to DR, cohorts of nonirradiated mice were exposed to AL diet or DR and simultaneously the dorsal hair was removed to analyze hair cycle activity. Nine to 14 days after depilation, dorsal skin of AL mice turned blue indicating anagen development, while DR mice had pink back as a sign of remaining in telogen (Fig. 2A–D). Twenty-five to 32 days after depilation, hair of AL mice had grown back completely, while dorsal skin of DR mice remained pink at this time point (Fig. 2D). When the hair of AL mice grew back completely (day 30 after the first depilation), the dorsal hair was depilated for a second round. Sixteen to 22 days after the second depilation (46 to 52 days after the first depilation), dorsal skin of AL mice turned fully blue, indicating that HFs had reentered anagen (Fig. 2D). Sixty-four to 72 days after the first depilation (34 to 42 days after the second depilation) the hair of AL mice had fully grown back.
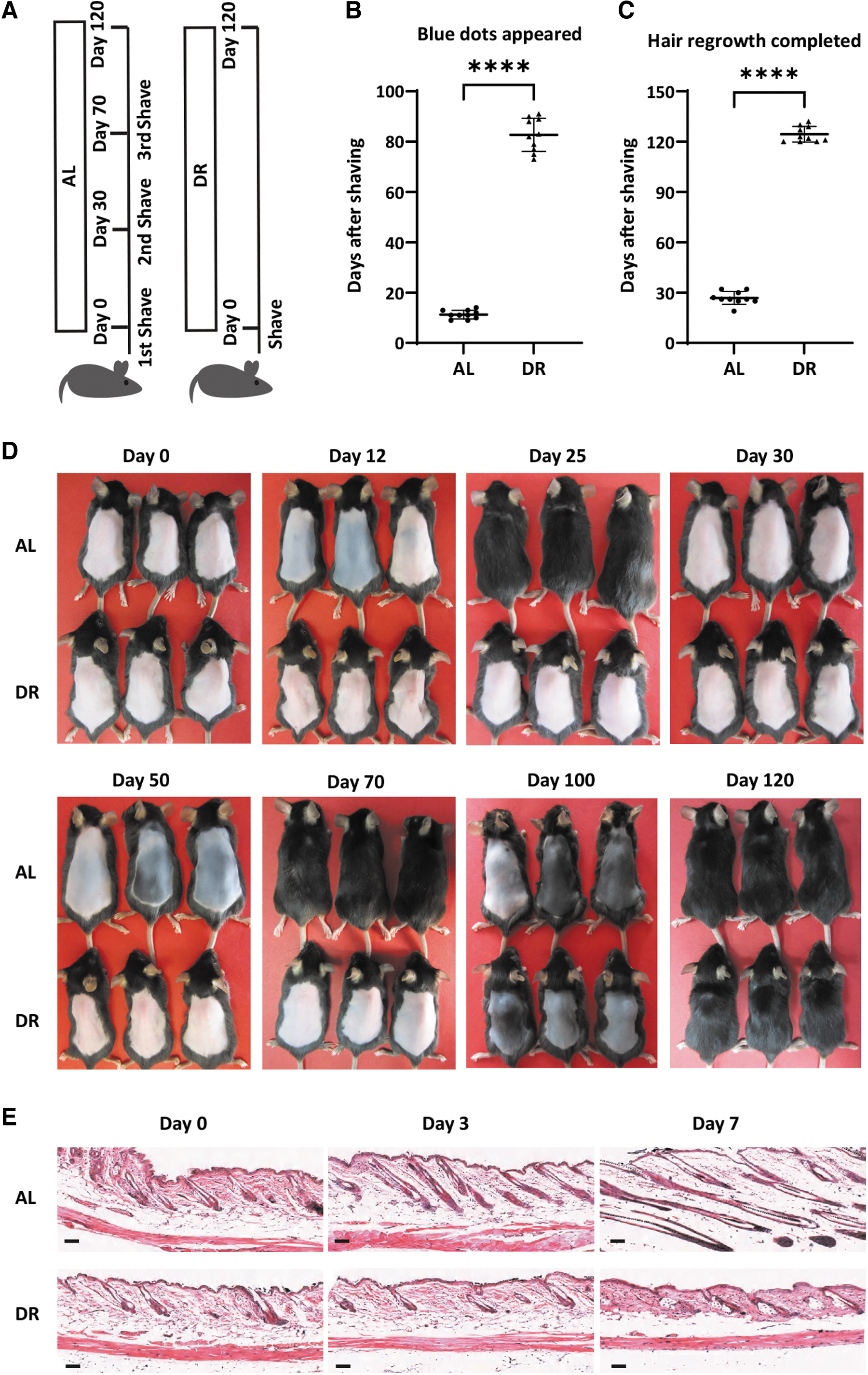
DR prolongs the telogen phase of HFs of nonirradiated mice.
The dorsal hair of AL mice was removed again, and their back skin returned pink at this time point, suggesting AL mice have finished two rounds of hair cycles after the first depilation and returned to telogen phase again. Of note, dorsal skin of DR mice remained pink indicating that HFs remained in telogen throughout this period (Fig. 2D). After the first depilation, dorsal skin of DR mice started to show blue dots at day 73 to 91 (Fig. 2B, D, 82.70 ± 6.584 days in DR mice vs. 11.30 ± 1.703 days in AL mice in the first hair cycle, n = 10 mice per group, p ≤ 0.0001, Unpaired two-tailed, Student's t-test) and had hair fully grown back at day 120–132 (Fig. 2C, D, 124.40 ± 4.648 days in DR mice vs. 26.80 ± 3.853 days in AL mice in the first hair cycle, n = 10 mice per group, p ≤ 0.0001, Unpaired two-tailed, Student's t-test). These findings indicate that DR leads to a very strong prolongation of the resting phase of HFs after a depilation-induced stimulus. In line with the gross skin illumination, skin sections also showed a clear induction of proliferative stages of HFs of AL mice at day-7 after the first depilation while HFs of DR mice were kept in telogen phase (Fig. 2A–C).
Future work should explore this phenotype in greater detail, for example by analyzing proliferation, cell death, and differentiation of HFSCs and MeSCs using mouse models such as K5tta-TetO-H2BGFP mice or Dct-lacZ transgenic mice.
DR prevents IR-induced depletion of cKit+, activated MeSC (melanoblasts)
Hair pigments are produced by melanocytes, which are generated by MeSCs during anagen. 1,16 Ionizing irradiation induces MeSC differentiation resulting in the loss of MeSCs by conversion of MeSCs into differentiated, pigmented melanocytes in mid-anagen phase. 16,20
Since DR prevents HFs from entering anagen phase, we further studied its effects on the maintenance of MeSCs by analyzing the number of ckit+ MeSCs (melanoblasts) at 1 month after IR and removal of the dorsal hair of the mice. At this time point, AL mice had run through one round of hair cycle and returned to telogen phase (see above), while HFs of DR mice remained in telogen phase without entering a round of hair cycling. Of note, the percentage of HFs containing ckit+ melanoblasts (80.83% ± 10.680% in DR mice vs. 51.67% ± 9.832% in AL mice, n = 6 mice per group, p = 0.0006, Unpaired two-tailed, Student's t-test) and the number of ckit+ melanoblasts per ckit+ HF (2.376 ± 0.2402 in DR mice vs. 1.336 ± 0.0906 in AL mice, n = 6 mice per group, p < 0.0001, Unpaired two-tailed, Student's t-test) were significantly higher in irradiated, DR-exposed mice compared with irradiated, AL-fed mice (Fig. 3A–D).
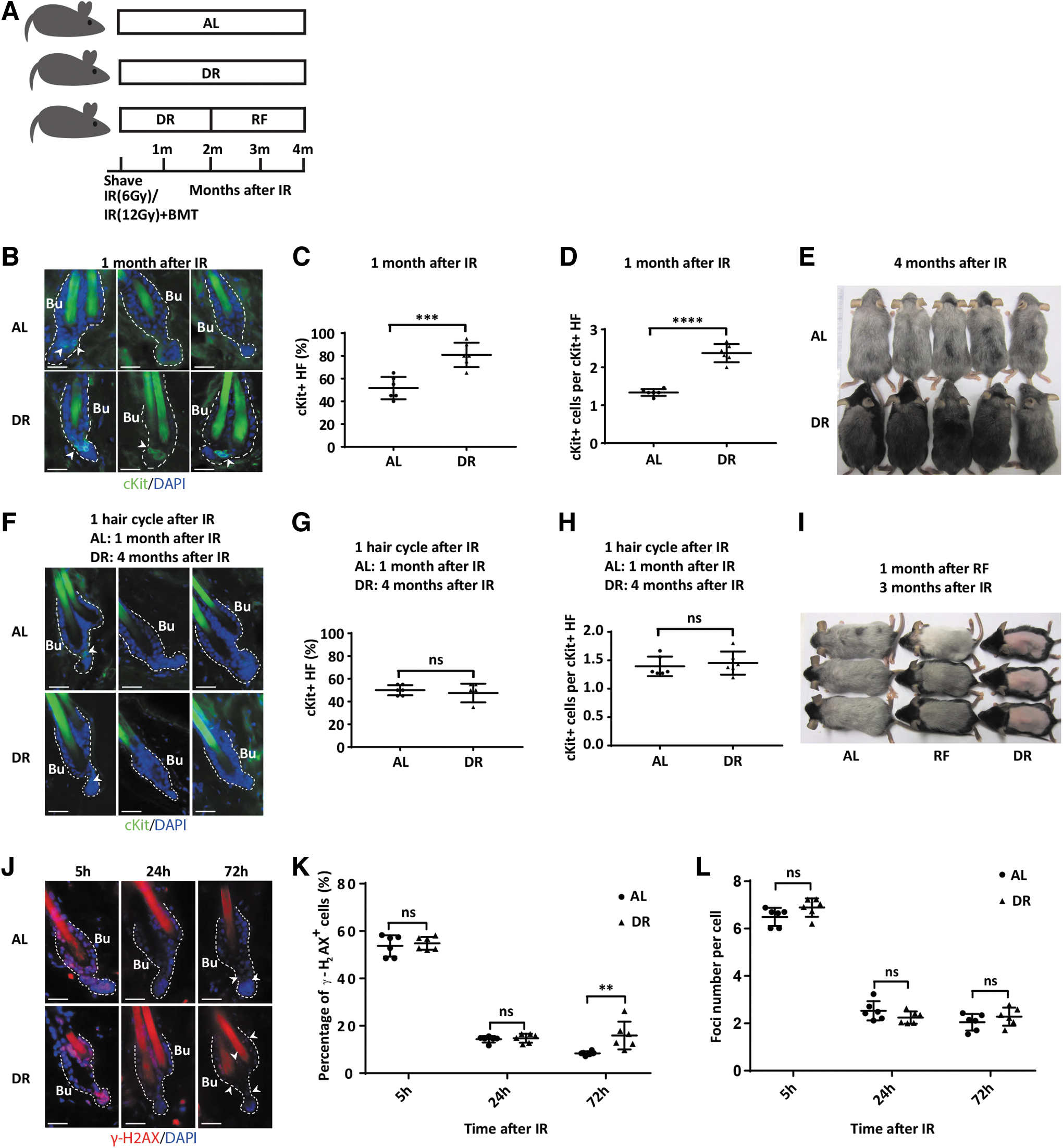
DR prevents a DNA damage induced depletion of activated, ckit+ MeSCs (melanoblasts) but fails to preserve melanoblast maintenance after resumption of cell cycle activity even after prolonged periods of DR-induced quiescence.
To test whether the prolonged quiescence of stem cells and HFs was indeed responsible for the protective effects of DR on hair graying, the back of irradiated, AL versus DR-exposed mice was shaved before the treatment procedure and the animals were followed until hair would finally regrow in the DR group. In the DR group, dorsal hair fully grew back only 4 months after IR compared with 1 month after IR in AL-fed mice (see above). Notably, the newly grown hair of DR mice, 4 months after IR followed by DR, was also gray, indicating that DR delayed but did not prevent the occurrence of DNA damage-induced graying (Fig. 3E). To analyze whether the delayed onset of graying in irradiated, DR-exposed mice would also associate with a depletion of ckit+ melanoblasts, skin was collected for histological analysis when both AL mice and DR mice had finished one round of hair regrowth after IR, which was 1 month for AL-fed mice and 4 months for DR-exposed mice after IR.
Notably, there was no difference in the percentage of HFs containing ckit+ cells (47.50% ± 8.216% in DR mice vs. 50.00% ± 4.472% in AL mice, n = 6 mice per group, p = 0.5275, Unpaired two-tailed, Student's t-test) and the number of ckit+ melanoblasts per ckit+ HF (1.452 ± 0.2038 in DR mice vs. 1.395 ± 0.1718 in AL mice, n = 6 mice per group, p = 0.6087, Unpaired two-tailed, Student's t-test) between AL mice and DR mice when comparing these two time points (Fig. 3G, H). These data suggested that DR can only delay but not fully prevent the loss of ckit+ melanoblasts in irradiated mice after resumption of cell cycle. While hair graying and loss of MeSCs was significantly delayed in DR- versus AL-exposed mice (Figs. 1B, D, 3A–D), the loss of ckit+ melanoblasts also occurred in DR mice after resumption of cell cycle activity, and after one round of anagen, the loss of ckit+ melanoblasts was similar in DR- versus AL-fed mice (Fig. 3F–H).
To further substantiate the connection between hair cycle activity in anagen and the failure of DR to prevent IR-induced hair graying despite delaying it, an RF experiment was conducted. To this end, the dorsal hair of 7-week-old mice was removed and mice were irradiated followed by exposure to AL diet or DR. As described above, 3 months after irradiation, the hair of AL-fed mice was completely regrown and nearly completely gray, whereas the shaved, dorsal skin of DR mice remained pink and hairless. However, when DR-exposed mice were refed with an AL diet from 2 to 3 months after IR, the hair also completely grew back at the 3-month time point, and the newly grown hair was also nearly completely gray as in mice that were on AL diet for 3 months (Fig. 3I).
Altogether, these results support the conclusion that DR preserved maintenance of ckit+ MeSCs (melanoblasts) and delayed the onset of hair graying in irradiated mice by maintaining HFs in resting telogen. However, DR treatment could not prevent the late onset of hair graying and loss of melanoblasts upon the delayed activation of hair cycle anagen in continuously DR-exposed mice or in mice that were refed with an AL diet after DR exposure.
To further investigate the effect of DR on DNA damage and DNA repair in HFs, expression of γ-H2AX was analyzed kinetically after IR in AL and DR mice. At early time points (5 and 24 hours) after IR, percentage of γ-H2AX+ cells and foci number per γ-H2AX+ cell were similar between AL and DR mice (Percentage: 5 hours, 54.80% ± 2.718% in DR mice vs. 53.76% ± 4.493% in AL mice, p = 0.9025; 24 hours, 14.76% ± 1.877% in DR mice vs. 14.33% ± 1.336% in AL mice, p = 0.9921; Foci number: 5 hours, 6.887 ± 0.3871 in DR mice vs. 6.492 ± 0.3828 in AL mice, p = 0.3747; 24 hours, 2.243 ± 0.2601 in DR mice vs. 2.529 ± 0.4046 in AL mice, p = 0.6711; n = 6 mice per group per time point, two-way ANOVA) (Fig. 3J–L).
Intriguingly, DR mice had significantly more γ-H2AX+ cells remained at later time point (72 hours) after IR (15.910 ± 5.8490 in DR mice vs. 8.387 ± 0.8441 in AL mice; n = 6 mice per group, p = 0.0031, two-way ANOVA, although foci number per cell remained similar between the two groups (2.283 ± 0.3822 in DR mice vs. 2.047 ± 0.3511 in AL mice; n = 6 mice per group, p = 0.3672, two-way ANOVA) (Fig. 3J–L), suggesting that DR delayed DNA repair.
The possible underlying mechanism could be that AL allows while DR blocks induction of proliferation (Fig. 1C), which is important for DNA repair process. These data indicated that HFs harbor DNA damage upon IR both under AL and DR conditions. However, DR delays the adverse effects of DNA damage on MeSC depletion but delays DNA repair in the affected cells.
DR inhibits proliferation while not affecting apoptosis of HF cells
To further test inhibition of proliferation of HF cells, we performed IF staining of PCNA on skin sections from both IR and NIR mice exposed to DR or AL. In addition to a clearly prolonging of HFs, the skin samples from AL mice showed a clear induction of expression of PCNA while it was hardly detected with a constant short shape of HFs in samples from DR mice both under IR and NIR conditions (Fig. 4A–D). In line with that, statistical analysis shows that the number of PCNA-positive cells per bulge in DR mice was significantly lower than AL mice on day 3 and 7 after depilation both under IR and NIR conditions (IR: day 3, 1.300 ± 0.2000 in DR mice vs. 8.867 ± 1.877 in AL mice, p = 0.0023; day 7, 0.7333 ± 0.1528 in DR mice vs. 13.3700 ± 2.5150 in AL mice; NIR: day 3, 1.867 ± 0.6110 in DR mice vs. 9.733 ± 1.106 in AL mice, p = 0.0004; day 7, 0.6667 ± 0.4163 in DR mice vs. 16.70 ± 1.2770 in AL mice; n = 3 mice per group per time point, p < 0.0001, Unpaired two-tailed, Student's t-test). We also performed TUNEL staining to determine apoptosis rate in bulge cells.
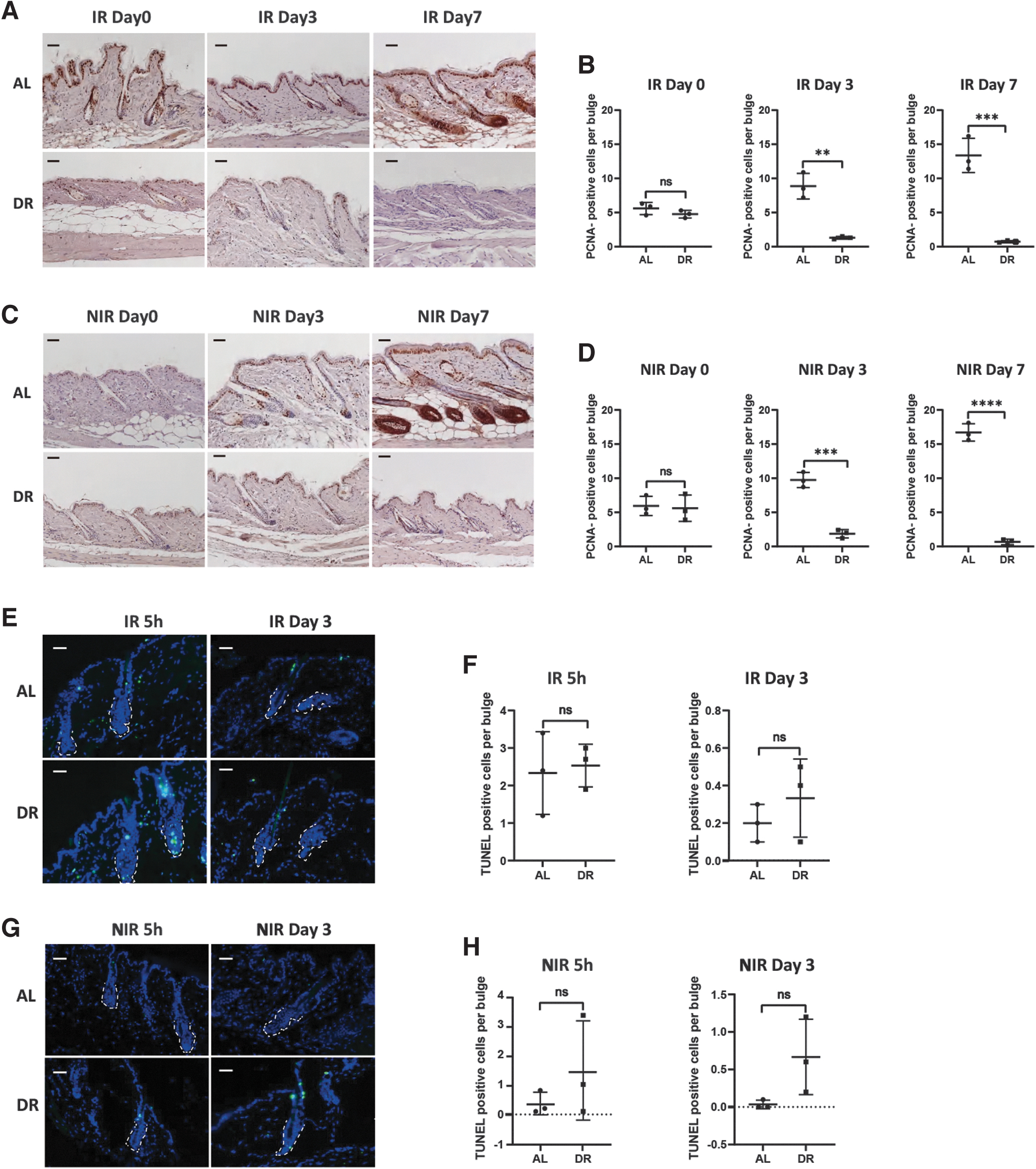
DR inhibited proliferation with neutral effect in HF cells.
The results show that the number of TUNEL-positive cells was not significantly different between AL and DR mice both under IR and NIR conditions (Fig. 4E–H). These results indicate that DR strongly suppresses proliferation while having no significant effect on apoptosis in the hair bulge cells as compared with AL mice.
Discussion
This study provides the first experimental evidence that DR can prolong the maintenance of activated MeSCs (melanoblasts) in mice that were exposed to irradiation-induced DNA damage. The preservation of melanoblasts associates with DR-induced prolongation of the resting phase of the hair cycle and quiescence of HFSCs and MeSCs. 22
The current results are different from observed increases in hair growth after plugging of mice that were treated with DR for 6 months compared with AL-fed mice. 12 Multiple parameters could explain the different outcomes, including (1) the use of a different mouse strain (Swiss mice in the study of Forni et al. compared with C57Bl-6J in our study); (2) the analysis of a different time point: our study shows that DR keeps HF in quiescence over 3 months after initiation of DR in telogen of 7-week-old mice. In contrast, the study of Forni et al. initiated DR in 8-week-old mice. It was not tested in which stage of the hair cycle the mice were at that age. Also, the Forni et al. study only investigated hair growth after 6 months of DR without investigating difference in hair cycling at earlier time points. Our study showed that after 4 months after initiation of DR, mice exhibit an induction of only one hair cycle, whereas AL-fed mice have already undergone three hair cycles at that time point. Based on these results it is well possible that Forni et al. investigated a time at which DR mice reentered the hair cycle after a prolonged arrest.
Interestingly, however, Forni et al. also noted a reduced replenishment of hair shafts in their study, speculating that DR may also reduce hair cycle activity, which is now proven by our study.
The current study presented a well characterization of the effects of DR in the hair graying and hair cycle at the phenotypic and histological level. However, despite the very interesting finding and possible important implication in understanding aging, the current study lacks mechanistic depth at a molecular level in understanding the phenotype. A gene expression profiling on sorted MeSCs and HFSCs and detailed analysis of behavior of these stem cells in response to DR after IR could help further mechanistically understand the role of DR, which was not performed in the current study due to a lack of mouse models, such as Dct-EGFP or Tyr-CreER mice. Therefore, the study does not delve beyond a descriptive study. How DR mechanistically regulates the hair cycle, HFSCs and MeSCs, especially the DNA repair in MeSCs, was not studied, and we can only speculate that the DNA repair was improved by DR given the observed phenotype.
The current study indicates that the prolonged preservation of dormancy of MeSCs in response to DR does not rescue the depletion of MeSC in response to RF after hair cycle reentry, even after prolonged periods of arrest time. Since most of the genes involved in DNA repair are also cell cycle dependent, therefore the delayed DNA repair that were detected by γH2AX staining in the current study could be a result from the suppression of proliferation by DR. It is tempting to speculate that the long-term failure of DR to rescue MeSC's maintenance (after resumption of hair cycle activity) could be related to the downregulation of DNA damage repair pathways in DR-exposed mice versus AL-fed mice. It would be an important field of future studies to delineate, which repair pathways are functionally affected by DR in MeSC. If it would be possible to reactivate these pathways in DR-exposed mice, such an intervention may have the potential to achieve a long-term, DR-mediated rescue in the maintenance of MeSCs and HFSCs in DNA damage-exposed skin.
Disentangling DR-mediated signaling in quiescence maintenance versus suppression of DNA repair in stem cells could help the development of interventions aiming to increase survival of stem cell populations in the context of DNA damage by selective targeting quiescence maintenance without dismantling DNA repair. Such approaches could be beneficial to maintain stem cell function during aging and may also be useful to protect somatic tissue stem cells from DNA damaging agents that still remain the most frequent and effective therapies for many types of cancer. The model system of DNA damage-induced MeSC differentiation, which results in hair graying, 16,21 could represent a valuable in vivo model to delineate fundamental processes underlying these two effector pathways (DNA repair and stem cell quiescence) and how these are regulated by dietary interventions, such as DR.
Footnotes
Authors' Contributions
R.Q., X.Q., and M.S. performed and analyzed majority of all experiments. M.S., Y.W., and J.W. helped with mouse treatment and photographing. H.W. and D.T. helped with H&E staining and photographing. R.Q. and S.T. conceived and designed the experiments. The article was written by X.Q., M.S., and S.T. and commented on by all other authors.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the National Natural Science Fund of China (NSFC-82160029), and the Jiangxi Provincial Nature Science Foundation (20204BCJ22027 and 20204BCJ22030).