
Abstract
Circadian rhythms (CRs) are 24-hour periodic oscillations governed by an endogenous circadian pacemaker located in the suprachiasmatic nucleus (SCN), which organizes the physiology and behavior of organisms. Circadian rhythm disruption (CRD) is also indicative of the aging process. In mammals, melatonin is primarily synthesized in the pineal gland and participates in a variety of multifaceted intracellular signaling networks and has been shown to synchronize CRs. Endogenous melatonin synthesis and its release tend to decrease progressively with advancing age. Older individuals experience frequent CR disruption, which hastens the process of aging. A profound understanding of the relationship between CRs and aging has the potential to improve existing treatments and facilitate development of novel chronotherapies that target age-related disorders. This review article aims to examine the circadian regulatory mechanisms in which melatonin plays a key role in signaling. We describe the basic architecture of the molecular circadian clock and its functional decline with age in detail. Furthermore, we discuss the role of melatonin in regulation of the circadian pacemaker and redox homeostasis during aging. Moreover, we also discuss the protective effect of exogenous melatonin supplementation in age-dependent CR disruption, which sheds light on this pleiotropic molecule and how it can be used as an effective chronotherapeutic medicine.
Introduction
Circadian rhythms (CRs) are rhythmic patterns that occur approximately every 24 hours in various biological processes, including sleep/wake cycles, hormone regulation, attention, cognitive function, and emotional state. 1 These inherent rhythms play a vital role in the survival of organisms as they enable synchronization of internal biological functions with external cues from the environment, particularly the alternating light and dark cycles.
CRs are found in a wide range of living organisms, spanning from microorganisms to higher mammals. 2 Within mammals, the circadian pacemaker is located in the brain, specifically in a region called the suprachiasmatic nucleus (SCN). 3 The SCN functions as the master biological clock, coordinating the peripheral circadian clocks to ensure optimal performance in response to external time cues (zeitgebers). This synchronization is crucial for maintaining a healthy and efficient timekeeping system. 4
The SCN regulates rhythmic oscillations through a neuronal pathway that influences secretion of melatonin by the pineal gland. 5 The decrease in melatonin production observed during the aging process is attributed to the circadian system's inability to accurately coordinate peripheral rhythms. This occurs because melatonin regulates the activity of the SCN through a feedback mechanism that inhibits its function. 6
The rhythmic patterns of CRs undergo significant alterations as individual age, and these changes can potentially exacerbate the progression of the aging process. 7 Aging is a natural and universal phenomenon characterized by the gradual accumulation of physical and molecular dysfunction, deterioration of tissues, and reduced organ function. These changes increase susceptibility to age-related diseases and, ultimately, result in death. 8 Research conducted on molecular clocks in animal models demonstrates that as animals age, there is a decline in both the strength and consistency of rhythmic patterns. 9,10
Circadian rhythm disruption (CRD), which refers to a disturbance in biological timing, can manifest at various organizational levels within an organism. This disruption can affect diverse aspects, ranging from the molecular rhythms of individual cells to the misalignment of behavioral cycles with environmental fluctuations. 11 CRD can lead to various physiological irregularities, such as metabolic abnormalities, 12 neurodegeneration, 13 disturbances in the sleep–wake cycle, 14 oxidative damage, 15 compromised DNA damage response, 16 and reduced activity levels. 17
Considering the current understanding of the aging mechanism, there is a proposal to investigate chronotherapeutic methods to downregulate the adverse effects of aging. This approach focuses on addressing disruptions in the
Collectively, this review article examines the interplay between CRs and aging mechanisms. Additionally, it summarizes the latest discoveries regarding utilization of exogenous melatonin as a chronotherapeutic drug against deleterious effects of CRD during aging.
Ethical Approval No: IAEC/AU/2019(1)/08.
The Circadian System
Continuous rotation of the Earth around the Sun, along with rotation on its axis, gives rise to an intricate, yet remarkably predictable, series of rhythmic alterations in the physical environment. Synchronization of rhythms to exactly 24 hours is achieved through entrainment to environmental cues known as zeitgebers, such as the daily alternation of day and night or temperature variations.
Entrainment results in establishment of a distinct phase of expression, where a particular point in a rhythmic process (such as a peak or trough) becomes associated with a specific time of day. The circadian clock governs the regulation of numerous processes, making chronobiology crucial for comprehending how neuronal function is temporally regulated. The circadian pacemaker in mammals is situated within the SCN, which receives direct input from the retina and is responsible for synchronizing the internal rhythms with external light cues. 19
The cells within the SCN operate as a closely interconnected network of neurons, and the neuropeptides, vasopressin and vasoactive intestinal peptide (VIP), play significant roles in facilitating communication between these cells. 20 The pacemaker located in the SCN also governs the synchronized phase observed in various other tissues throughout the body. Cells outside of the SCN possess an independent circadian rhythmicity. 21
Our current understanding of how the SCN controls rhythmic activity in peripheral tissues is very limited and needs to be explored. However, experiments have indicated the significance of neuronal signals, humoral signals, and intermediary pathways that are influenced by factors such as food intake and body temperature. 22,23
Each cell possesses a molecular clock that constitutes an essential component of the circadian system (Fig. 1). The molecular clock comprises a transcriptional/translational feedback loop (TTFL) that undergoes cyclical activity lasting ∼24 hours even in the absence of external stimuli. 24 Light is recognized as the most influential zeitgeber, although the feeding cycle, temperature, and physical exercise can also serve as external stimuli for resetting the circadian clock. 25

Mammalian circadian clock. Ccgs, clock-controlled genes; BMAL1, brain and muscle Arnt-like protein 1; CLOCK, circadian locomotor output cycles kaput; CRY, cyrptochrome; PER, period homolog; ROR, retinoid-related orphan receptor.
The primary transcriptional activators encompass proteins known as circadian locomotor output cycles kaput (CLOCK), which bind to brain and muscle Arnt-like protein 1 (BMAL1). These proteins form a heterodimer that recognizes E-box motifs and governs the expression of numerous genes. 26 Among the clock-controlled genes, there are genes such as period (Per1, Per2, and Per3) and cryptochrome (Cry1 and Cry2), which function as repressors. 27 During the 24-hour cycle, PER and CRY proteins are phosphorylated and subsequently return to the nucleus to hinder the transcriptional activity of the CLOCK-BMAL1 complex. This inhibition leads to suppression of their expression, forming a negative feedback loop.
Furthermore, additional pivotal regulators of the circadian clock include the nuclear receptors, REV-ERBα and REV-ERBβ, along with the retinoic acid orphan receptors (RORs) (RORα, RORβ, and RORγ), which together form another feedback loop. 28 In particular, REV-ERBs function as transcriptional inhibitors of BMAL1 expression, whereas RORs play a positive role in regulating BMAL1 expression. RORs achieve this by binding to specific sites known as retinoic acid receptor-related orphan receptor elements (ROREs) located in the promoter region of the BMAL1 gene.
Collectively, these TTFLs operate in a cyclical manner and are fundamental for establishment of biological rhythms. 29
The Circadian Clock Network During Aging
To synchronize clocks in peripheral tissues, the master circadian clock, SCN, plays several roles that are altered throughout aging. For instance, the reduced ability of the lens to transmit light in older individuals leads to diminished sensitivity in detecting and responding to light as a signal for synchronization, 30 fewer retinal ganglion cells with intrinsic photoreceptors that transmit photic information to the SCN, 31 and reduction in the functioning of the SCN itself.
A neurotransmitter VIP, an essential regulator of intercellular communication among individual SCN neurons, is expressed by fewer neurons in the old SCN in comparison with the young SCN. 32 In addition, the SCN in older people exhibits a decrease in the number of synaptic terminals and an alteration in the intercellular connectivity between individual neurons. 33 As a result, the combined decline in (a) sensitivities to light input, (b) signal transmission efficiency, and (c) connectivity within neural networks leads to a diminished ability of the aging SCN to respond effectively to light stimuli.
Since there is a functional decline in the SCN with age, it significantly affects longevity. Transplanting the SCN from a young hamster into an old hamster extends life expectancy by ∼20%. 34 During aging, SCN neuronal cells have a lower circadian amplitude in intracellular signaling, and aging affects core clock gene expression in the SCN in a gene-specific manner. 35 These factors are likely implicated in the diminishing circadian clock outputs observed with age.
Bidirectional relationship between aging and CRs
With the continuous increase in the population of older individuals, it becomes imperative to enhance our understanding of the interdependent association between aging and CRs. Aging and age-related disorders are frequently a consequence of alterations in circadian clock functioning. Furthermore, some alterations may be indicators of/and triggers for age-related neurological diseases and cancer. Nevertheless, it remains uncertain whether circadian disruptions are indicative of the aging process itself or if these disturbances potentially contribute to accelerated aging and development of age-related disorders.
Disruptions in
Modern lifestyle and a growing aged population simultaneously exposed to persistent disturbances in CRs, including shift work and irregular sleep patterns. Multiple studies indicate that CRD has the potential to hasten the aging process and contribute to the development of age-related disorders. 37 For example, numerous studies investigating chronic jet lag suggest that circadian disruptions raise the risk of cancer, and individuals engaged in shift work may have an increased susceptibility to developing cancer. 38
Consequently, circadian disturbances could result in age-related disorders. Avoiding light exposure at night and using bright light therapy are two ways people can perhaps enhance their
Effect of Melatonin on SCN
The pineal gland releases melatonin during the dark period of the light–dark cycle and it suppresses the function of the master circadian clock through a feedback inhibitory mechanism. 39 It was commonly assumed that the impact of melatonin on the clock network in the SCN and other areas was attributable to the levels of circulating melatonin in blood.
This assumption was based on the well-established recognition of the blood melatonin rhythm for about 50 years, which has been extensively observed in numerous mammalian species. 40,41 The temporal pattern of melatonin in the bloodstream usually exhibits a gradual increase after the onset of darkness, reaching its highest point during the middle of the dark period, and subsequently decreasing during the latter half of the night.
On the other hand, the rhythmicity of melatonin in cerebrospinal fluid (CSF) rises promptly after the start of darkness, remains consistently elevated throughout the night, and experiences a sharp decline at the onset of light. Moreover, it is common for the amplitude of melatonin in the CSF to be higher compared with its levels in the bloodstream. 42
We proposed that instead of relying on blood melatonin levels, the
Alternatively, it can be transported from the CSF to the SCN through tanycytes. 43 If exogenous melatonin treatment leads to significantly higher levels of melatonin in the bloodstream compared with the CSF, these elevated blood levels likely override the signal provided by the concurrently reduced CSF melatonin levels. In such cases, melatonin acts as a chronobiotic for SCN. 44,45 This reveals how peripherally administered melatonin may serve as a pharmacological agent that affects the SCN rhythmic functions.
The SCN processes a variety of inputs and delivers neuronal and endocrine signals to peripheral oscillators in all cells to control physiological and behavioral responses, ensuring that they are temporally appropriate. 46 Even though in several species different brain areas have significantly different distributions of melatonin receptors in the central nervous system, 47 most mammals possess a significant quantity of high-affinity MT1 melatonin receptors that are coupled to G-proteins. 48 These receptors play a crucial role in mediating various functions of melatonin in the inherent circadian oscillations of the central endogenous clock.
However, there is another hypothesis that melatonin may directly interact with the SCN cells' proteasome, irrespective of any receptors, to alter temporal fluctuations in molecular clock genes. 5 The predominant light–dark environment strongly affects the electrical circuitry and metabolic regulations of the SCN neurons. For example continuous exposure to artificial light particularly of short wavelengths causes phase shift in circadian amplitude.
Several investigations have documented the correlation between SCN and age-related neurodegenerative disorders. 13,49 For example, the functional decline of SCN plays a crucial part in the pathogenesis of AD and progression of sleep disturbances. 50 Wu and colleagues 51 investigated the immunohistochemistry of the SCN in young and old participants and found that the number and density of MT1-expressing neurons decreased significantly in the older group.
Additionally, researchers observed that expression of MT1 receptors was exceedingly low in the advanced neuropathological stages of AD (Braak stages V–VI). As an individual ages, the presence of a consistent circadian melatonin pattern could potentially work in conjunction with SCN neurons to prevent neuronal degeneration and sustain
Melatonin and Regulation of Clock Genes
Since melatonin is mainly produced at night, it is often considered the “hormone of darkness.” 54 The length of the night is also reflected in the time that the nocturnal peak of melatonin secretion lasts. 55 Therefore, it is believed that the diurnal rhythms of melatonin are essential for healthy circadian function. 56 The existence of melatonin receptors explains the direct action of melatonin on various organs.
The relationship between melatonin production and the
Reduced levels of BMAL1, PER1, and CRY2 were found in the adrenal cortical region of C57BL mice when compared with C3H mice. 57 Pinealectomized mice did not exhibit diurnal cycles of Per1 mRNA and their protein levels in the striatum. 58 Melatonin lowered the expression of Per1 and Clock in primary neuronal cells generated from the murine striatum, while it enhanced the expression of NPAS2 and had no influence on the level of Bmal1. However, MT1 knockout animals did not exhibit these effects. 59
In a study examining the effects of melatonin on hypertensive TGR (mRen2)27 rats, it was observed that the expression of Per2 and Bmal1 in the heart was influenced by melatonin in a phase-dependent manner, particularly during the dark phase. These findings suggest that melatonin plays a role in regulating clock genes, although the exact mechanisms involved remain unclear. 60
In a study conducted by Mattam and Jagota, 61 rats belonging to different age groups were administered melatonin (30 μg/kg) 1 hour before the onset of the dark phase [zeitgeber time (ZT)-11] for 11 days. 61 Regardless of the light–dark phase or age group, administration of melatonin resulted in elevated levels of Bmal1, Per1, Per2, Cry1, and Cry2 mRNAs. These findings indicate that melatonin directly influences the transcription of these clock genes, although the gene expression patterns vary with the age of the animals.
Melatonin acts as a proteasome inhibitor, interfering with molecular loops to enhance BMAL1 and, subsequently, transcription of the Bmal1, Per, and Cry genes. 62 Transcriptional modification in the nuclear orphan receptors (Rors and Rev-Erbα mRNAs) was used to assess the chronobiotic functions of melatonin. While melatonin did not affect Rorα, it effectively prevented Rorβ levels from dropping in the early hours.
In the first subjective night after melatonin administration, expression of Rev-Erbα was phase advanced; however, Bmal1 was merely phase shifted on the second night. 63 Therefore, the authors postulated that melatonin and the molecular clock machinery may be connected by the nuclear orphan receptor genes.
Circadian Clock, OS, and Aging
An imbalance that favors oxidants over antioxidants, which disrupts cellular redox homeostasis and control and/or causes molecular damage, is referred to as OS. 64 An important aspect of the aging process involves a rise in OS, which is considered to be among the molecular mechanisms believed to contribute to the onset and progression of various age-associated diseases. OS is triggered and regulated by a variety of factors.
Strong evidence suggests that OS is caused by mitochondrial dysfunction and contributes to age-related functional decline, in part, by oxidative damage. 65 Additionally, several signaling pathways control OS. In cells of higher living organisms, both Nrf2 and NFkB signaling mechanisms are regarded as the central switch that regulates OS. 66
Numerous studies demonstrate the link between disruption of the abovementioned mechanisms of OS and aging. 67,68 The disruption of sleep–wake patterns, resulting from an increase in OS, has been suggested to share similarities with the aging process. This phenomenon is believed to be partially attributed to the progressive accumulation of oxidative damage during aging. 69
Mice that have particular gene disruptions in the insulin/insulin-like growth factor (IGF) pathway, apoptosis, redox homeostasis, chromosome stability, controlling of the mutation rate, and telomere stability exhibit signs of accelerated aging. 70 –75 In the presence of neurological diseases, interruption in circadian clock function may make the brain more susceptible to oxidative damage and promote neurodegeneration. 76
Strong evidence suggests a relationship between the cellular balance of redox reactions and the functioning of the circadian clock. 77 The circadian system maintains reactive oxygen species (ROS) within normal physiological levels and protects living organisms against oxidative damage. 78 According to studies, BMAL1 is required for promotion of antioxidant defense to protect cells from OS or aging. 79
Additionally, BMAL1 activates the Nrf2 pathway and B cell lymphoma-2 (Bcl-2) response to maintain the antioxidant capacity within vascular smooth muscle cells. 80 These findings suggest that the role of BMAL1 is crucial in orchestrating the Nrf2-dependent antioxidant defense mechanism, which serves to counteract OS and mitigate the aging process.
OS and BMAL1/CLOCK genes
Increased OS caused by the global loss of BMAL1 accelerates aging phenotypes. 81 Additionally, there is convincing evidence that BMAL1 controls ROS in a variety of distinct tissue types. Deletion of brain BMAL1 causes OS-mediated neurodegenerative processes and astrogliosis. 76 B cell failure triggered by OS and BMAL1 deletion in the pancreas results in the diabetes phenotype. 82 However, the use of the glutathione precursor, N-acetyl-cysteine, can reverse the accelerated aging phenotype caused by BMAL1 knockout. 83
All the evidence points to the crucial role of OS in the aging phenotype caused by BMAL1 deficiency. Additionally, arrhythmic per01 flies have shown substantially higher amounts of protein carbonylation and increased lipid hydroperoxidation during aging. 84 Both in vitro and in vivo experimental settings suggest that reduced Bmal1 expression further worsens OS-induced neuronal death. 76
The most remarkable neurological abnormalities that Bmal1 −-deficient mice exhibit are reduced brain functional communication, increased OS, synaptic deterioration, extensive astrogliosis, 76 impaired neurogenesis in the hippocampus, 85 and weakened memory and learning behavior. 86 Studies have demonstrated that BMAL1 plays a direct role in regulating the balance of ROS, indicating its potential involvement in the protective mechanisms employed by cells and tissues to counteract OS. 87
Additionally, CLOCK is involved in the regulation of various forms of senescence or the aging process in organs. According to Musiek et al., 76 eliminating the clock genes (either BMAL1 or CLOCK) results in oxidative harm, significant activation of astrocytes, deterioration of axonal terminals, and disturbance in the functional connectivity observed during periods of rest. This discovery establishes a new connection between CCGs, oxidative injury in the brain, and the neurodegenerative process.
An earlier study revealed that CLOCKΔ19 mutant mice exhibit a noticeable increase in peroxidation and endoplasmic reticulum stress (ERS), which ultimately leads to a striking representation of liver aging. 88
Melatonin as a Potential Antioxidant
Maintaining cellular redox balance is crucial for various physiological functions and is dependent on the proper control of ROS production and elimination. This equilibrium plays a significant role in regulating signaling pathways as well as facilitating the activation of redox-sensitive proteins and enzymes. Melatonin can counteract both ROS and reactive nitrogen species (RNS). 89
Due to its amphipathic nature, melatonin can effectively traverse cell membranes, enabling it to protect the subcellular compartments from damage caused by free radicals. 90 Melatonin achieves redox control through its ability to directly scavenge ROS using defense mechanisms of both enzymatic and nonenzymatic antioxidants. 91 –93 With its aromatic indole ring containing a plentiful supply of electrons, melatonin acts as a potent electron donor, effectively neutralizing free radicals and demonstrating its direct antioxidant characteristics. 90,94
Melatonin exhibits indirect antioxidant properties by stimulating the activation of antioxidant enzymes such as SOD, GSH-Px, and CAT 95 and by promoting the synthesis of glutathione. 96 These actions enhance the effectiveness of other antioxidants, leading to a combined synergistic effect. 97
Melatonin elicits cellular responses through both receptor-mediated and receptor-independent pathways. 98 Melatonin possesses the ability to directly traverse cell membranes without relying on receptors, enabling it to scavenge ROS within the cytosol and mitochondria of the cell. Melatonin is concentrated within the mitochondria, where it neutralizes both ROS and RNS. 99
Furthermore, melatonin provides a defense against the peroxidation of cardiolipin and protects respiratory chain complexes and mtDNA against oxidative damage induced by free radicals. 100 The antiapoptotic action of melatonin inhibits the function of Bax, resulting in a reduction in the opening of the mitochondrial permeability transition pore (mPTP) and ultimately decreasing the permeability of the membrane. 101,102
Melatonin and Nrf2
Nrf2 is acknowledged as the primary defense mechanism against OS, occupying a pivotal position in the transcriptional control of genes responsible for the synthesis of antioxidant enzymes, antiapoptotic transcription factors, detoxifying molecules, and drug transporters. 103 Research findings have indicated that melatonin enhances upregulation of genes and proteins linked to Nrf2 signaling and the functions of the peroxisome. By eliminating dysfunctional organelles such as mitochondria, autophagy mediated by Nrf2 enhances the effectiveness of adaptive mechanisms. 104
Melatonin shields Nrf2 from degradation and initiates the activation of the Keap1-ARE (Kelch-like ECH-associated protein 1–antioxidant response element) pathway. 105,106 The interaction between melatonin and Nrf2 enhances various beneficial effects, including cardioprotection, neuroprotection, antioxidant activity, anti-inflammatory effects, and antitumor effects, as well as improvements in bone microstructure and oocyte maturation. 107,108
Mitochondrial aging and melatonin
Aging contributes significantly to the decline in respiratory chain efficiency, amplification of electron leakage, and decrease in ATP synthesis, collectively resulting in mitochondrial dysfunction. 109 As a result, the impaired function of mitochondria initiates generation of ROS, subsequently promoting additional mitochondrial injury and cell death. 110 The level of melatonin within mitochondria seems to be higher than the amount found in the bloodstream. 111 This is particularly advantageous since free radicals primarily originate from these cellular organelles.
Apart from efficiently counteracting ROS within these organelles, melatonin can decrease electron leakage from complexes within the mitochondrial respiratory chain, thereby enhancing ATP synthesis. 112 The action of reducing electron leakage also leads to a decrease in generation of free radicals, a phenomenon commonly referred to as radical avoidance. 113 Melatonin demonstrates remarkable effectiveness in repairing damage, specifically targeting complexes I and IV.
Moreover, melatonin protects against the harmful effects induced by mitochondrial Ca2+ overload and depolarization of the mitochondrial membrane caused by H2O2, both of which are associated with caspase-mediated apoptosis. 114 Based on the mechanism, melatonin-induced pro-oxidant activity can accelerate production of ROS by interacting with calmodulin. The interplay between melatonin and opening of mitochondrial transition pores or its complex III also stimulates generation of ROS. 89,115
Mitophagy and melatonin
Mitophagy plays a crucial role in preserving the redox stability of mitochondria by effectively reducing excessive production of ROS at the same time. Melatonin has exhibited promising potential in treating mitochondrial dysregulation through stimulation of mitophagy. 116 Since mitochondrial oxidative damage is believed to be both the source and a potential target of ROS, reduced mitophagy is directly associated with decreased life span (Fig. 2). 117,118
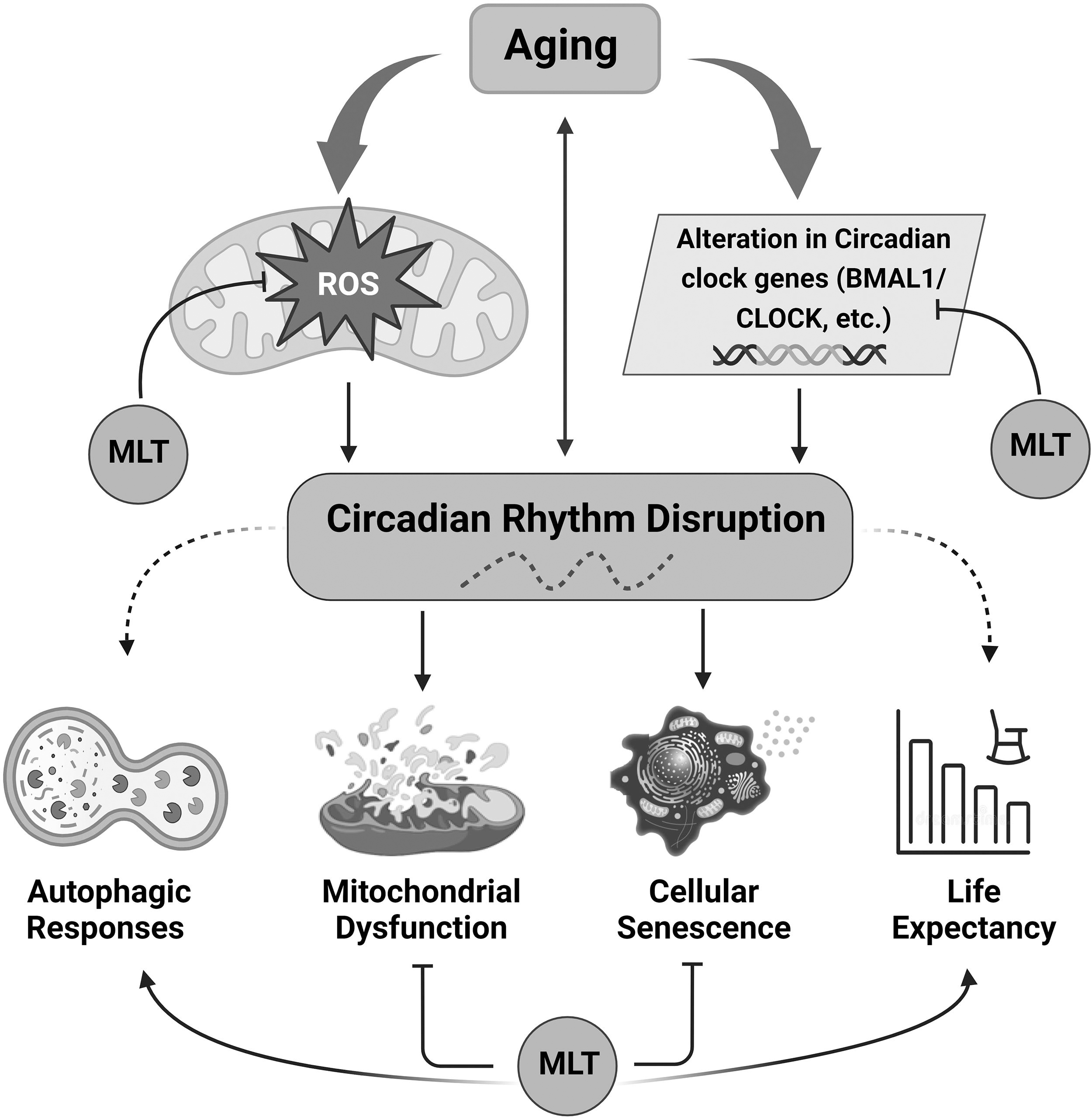
Effect of melatonin on consequences associated with CRD. An excess of ROS results in oxidative injury, which contributes to onset of diseases and reduces the life span. Melatonin directly inhibits ROS and downregulates the expression of CCGs through feedback inhibitory mechanisms. The disturbance of circadian rhythms hinders the upkeep of physiological and cellular balance, leading to cellular senescence, mitochondrial dysfunction, and an accelerated aging process. Stimulation is indicated by arrows, while reduced responses are indicated by dashed arrows. ROS, reactive oxygen species; CCG, circadian clock gene; CRD, circadian rhythm disruption; MLT, melatonin.
Melatonin decreases the leakage of electrons from mitochondria, thereby reducing the production of ROS while preventing mPTP opening in unfavorable bioenergetic conditions, thereby preserving mitochondrial membrane potential (Δψ). Additionally, in normal circumstances, melatonin stimulates uncoupler proteins to gradually decrease Δψ. 119
Emerging senotherapeutic approaches that focus on mitochondria and aim to activate melatonin-induced mitophagy have the potential to improve mitochondrial dysfunctions. Several feasible interventions, such as melatonin, have been examined for their potential to slow down aging in the senescence-accelerated mouse prone 8 (SAMP8) strain, although they have yielded conflicting results. 120 –122
Melatonin treatment can regulate molecular and morphological characteristics associated with senescence. Studies have shown that melatonin can enhance mitochondrial oxidative phosphorylation in both liver and brain tissues. 123,124 Mesenchymal stem cells (MSCs) undergoing replicative senescence often experience reduced mitophagy due to inhibition of mitofission, leading to increased mitochondrial dysfunction.
The treatment with melatonin inhibited replicative senescence by enhancing mitophagy and promoting mitochondrial activity through upregulation of the heat shock 70-kDa protein 1L (HSPA1L). 125 Although it is not possible to halt the natural aging process, mitigating the disruptive consequences of age-related changes in overall homeostasis in individuals is certainly achievable. The current utilization of melatonin as both a senotherapeutic and chemotherapeutic agent indicates that melatonin will continue to be extensively investigated in the future as one of the prominent molecules of interest.
Melatonin, aging, and CRD
It has been demonstrated that age-related alterations in melatonin synthesis are linked to CRD. Adults have a 200%–300% a rhythm-adjusted mean (MESOR) variation in the melatonin circadian pattern in their blood and saliva, but this amplitude is barely noticeable in people older than 80 years and decreases considerably after the sixth decade of life. 126 Although both MESOR and amplitude decrease, the synthesis of melatonin continues in longer lived humans.
Some authors contend that pineal production of melatonin declines with age and is not a necessary component of the aging process, while there have been several reports about the multiple beneficial impacts of melatonin on gerontological practice, and the effectiveness of melatonin in extending the life span has been shown in human beings as well as animals. 127,128 Importantly, changes in CRs of melatonin excretion are a major factor in peripheral mechanisms that interfere with a biosystem's natural temporal patterns during aging. 128
Prevention of the symptoms of age-related circadian disturbances is one of the possible mechanisms that allow the geroprotective action of melatonin. 129 Additionally, since melatonin deficit is linked to the advancement of macular degeneration, it suggests that inadequate endogenous production of melatonin escalates the signs of age-associated CRD, which have a connection to the afferent component of the circadian system. 130
Interestingly, chromaffin cells of the intestine were found to produce less endogenous melatonin during aging. 131 These findings suggest that endogenous production of melatonin in the intestine as well as pharmaceuticals derived from melatonin may be crucial for preventing oxidative damage and tissue-specific CRD, which work in combination with melatonin synthesized by the pineal gland.
Melatonin's primary chronobiotic effect is its hypothermic action, which is linked to an increase in sleep quality and contributes to prevention of CRD, particularly that connected to aging. 132 Interneuronal connectivity in the SCN deteriorates as a result of the SCN's impaired ability to compensate for temperature variations; therefore, the temperature factor plays a more substantial part in synchronization of CRs and contributes to resynchronization of neurons. 133
These CRDs are not only present in old mammals but can also be seen in middle-aged individuals. 9,134 A double-blind placebo-controlled investigation demonstrated that melatonin effectively restores age-related CRD by synchronizing the rhythmicity of body temperature, blood pressure, and heart rate. 135
Melatonin plays an important role in maintaining normal sleep patterns.
136
Currently, the disturbances in sleep observed among several aged individuals are believed to be mainly attributed to modulations in the
The circadian system is adversely affected by exposure to artificial light at night due to reduction in the release of the melatonin hormone, 94,139 which can lead to the emergence of CRD. Reduced melatonin levels accelerate aging, raise the probability of developing malignant tumors early in life and metabolic diseases, and ultimately shorten the life span. 128,130,140 Due to its ability to act as an antioxidant, prevent persistent inflammation, 141 and exhibit contrasting impacts on cell apoptosis in both healthy and cancerous states, melatonin plays a vital role in prolonging the life span and delaying the initiation of the aging process.
Melatonin seems to be inadequate for treating sleep disorders in shift workers—rather, it may lengthen their sleep cycles 142 with almost no side effects, as assessed in various clinical investigations and comparative trials throughout short periods (up to 3 months, at low doses). 143,144 Low doses of melatonin are generally safer and an improved substitute for sleeping pills that cannot be taken for an extended duration because they are not advised for long-term usage. 144
Melatonin-rich supplements could lengthen rest periods, delay disease onset, and reduce nighttime awakenings in individuals suffering from autism, particularly in children. 145,146 Recent research indicates that melatonin may be an effective and safe treatment for depression and sleeplessness in adults and children with attention-deficit/hyperactivity disorders, but longer and more comprehensive trials are required to determine the drug's long-term safety and ideal dosages. 147,148
Developing metabolic phenotype technologies can assist in identifying markers that accurately reflect how melatonin affects sleep, aging, and onset of diseases. 130,149
Conclusions
CRs are governed by an internal timekeeping machinery that regulates physiological and behavioral processes. The circadian clock is positively influenced by CLOCK and BMAL1, transcription factors that regulate the functioning of the circadian pacemaker. CRs contribute to the functioning of various physiological processes and individuals experience changes as a result of aging and associated diseases.
As organisms age, a decline in melatonin levels is commonly observed. This decrease in melatonin can result in various symptoms affecting the internal equilibrium of the organism. One of the frequently observed manifestations of this process is the deterioration of cellular clock regulation, leading to disruptions in proper functioning. Furthermore, the combination of diminished mitochondrial integrity and impaired functions, along with reduced melatonin levels, increases the vulnerability of older individuals to impaired regulation of redox processes and age-related disorders.
Age-related health abnormalities have a direct correlation with CRD. Increasing evidence from diverse organisms indicates that mutations in CCGs result in premature aging, shortened life span, and disturbances in sleep patterns. ROS are generated as by-products during mitochondrial energy metabolism, primarily within the electron transfer chain.
The circadian clock is intricately linked to the mitochondrial physiology and contributes to the regulation of ROS by coordinating generation of ROS within mitochondria and activation of antioxidant defense mechanisms. Melatonin is recognized as a “master hormonal regulator” that influences the CR, aging process, and cellular activity. Additionally, melatonin holds promise as a pharmacological agent due to its potent antioxidant properties.
The rhythmicity of melatonin in CSF also influences the molecular network within the SCN, thereby preserving optimal CRs. Furthermore, in the aging brain, mitochondria tend to generate excessive, harmful, partially reduced oxygen species. In this context, melatonin serves as a protective neuroendocrine molecule for this tissue by directly scavenging free radicals and stimulating antioxidative enzymes, thus preventing molecular damage. 99,150
The area of chronobiology research is progressively shifting toward developing innovative approaches for chronotherapy by focusing on either the molecular clock or administering medications based on specific times of the day. Enhancing biological clock functions and rectifying its disturbances through pharmacological intervention may offer a novel approach for effective management of aging and age-related diseases in a clinical setting.
To this end, different therapeutic doses of melatonin efficiently restore CRD in humans and distinct animal models of aging and its associated diseases. However, it is essential to conduct longitudinal studies in future, including repeated follow-up assessments, to determine the translational application of these findings in clinical practice.
Footnotes
Authors' Contributions
A.K.V. and S.I.R. contributed to the conceptualization and review and editing of the article. A.K.V. wrote the article. M.I.K. and F.A. contributed to writing—review.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.