
Abstract
Recently, natural herbs have gained increasing attention owing to their comparatively low toxicity levels and the abundance of historical medical documentation regarding their use. Nevertheless, owing to a lack of knowledge regarding these herbs and their compounds, attempts to find those that could be beneficial for treating diseases have often been ad hoc; thus, there is now a growing demand for an in silico method to identify beneficial herbs. In this study, we present a computational approach for identifying natural herbs specifically effective in treating cognitive decline in Alzheimer's disease (AD) sufferers, which analyzes the similarities between herbal compounds and known drugs targeting AD-related proteins. Our in silico method suggests that Corydalis ternata can improve cognitive decline in AD sufferers. Behavioral tests with an AD mouse model for the confirmation of the in silico prediction reveals that C. ternata significantly alleviated the cognitive decline (memory and motor functions) caused by neurodegeneration. Further pathology analyses reveal that C. ternata decreases the level of Aβ plaques, reduces neuroinflammation, and promotes autophagy flux, and thus C. ternata can be clinically effective for preventing mild cognitive impairment during the early stages of AD. These findings highlight the potential utility of our in silico method and the potential clinical application of the identified natural herb in treating and preventing AD.
Introduction
The process of drug discovery and development generally takes approximately 9 years to bring a new drug into the market, and its costs could approach over $2.6 billion USD in the early 2010s. 1 These high costs are due to time-consuming and costly experiments as well as low success rates. To address these challenges, many machine learning-based computational models have been developed for drug target identification, drug design, and toxicity prediction. 2 –4 Despite computational assistance, many drugs/candidates are withdrawn during the preclinical stages or from the market owing to the toxicity issues of synthetic chemicals.
With the desire to avoid toxicity issues and accelerate drug discovery, researchers have been showing increasing interest in natural herbs and their products because of their comparatively low toxicity levels and the abundance of historical medical documentation regarding their use. Natural products have made major contributions to pharmacotherapy throughout history, but the difficulty in the extraction, isolation, and analysis of structurally complex natural compounds has been a barrier to natural product drug discovery. 5
Nevertheless, natural product drugs/extracts or derivatives have been marketed for treating various human diseases for which synthetic chemical drug treatments have not yet been developed. 6 Similar to conventional drug discovery, the demand for in silico approaches to facilitate the discovery of natural products continues to increase.
Alzheimer's disease (AD) is a progressive neurodegenerative disorder characterized by neuronal cell death induced by the protein aggregates, such as amyloid-β (Aβ) and tau, and cognitive dysfunctions. 7,8 The number of AD sufferers experiencing cognitive decline is increasing worldwide; this has resulted in an increase in the severe social and economic burdens necessary for their care.
Despite extensive studies on AD pathophysiology, many drugs have been withdrawn during development because they showed no significant improvement or caused severe side effects. 7,8 Only two types of drugs are currently used for AD treatment: cholinesterase enzyme inhibitors and N-methyl-D-aspartate (NMDA) antagonists. 8
However, these drugs are effective only in mild-to-moderate AD cases, often failing to prevent AD progression; moreover, they may even cause severe adverse effects. For example, memantine, an NMDA antagonist, improves memory impairments in AD sufferers. 9 Nonetheless, it does not prevent the progression of AD and can cause side effects such as dizziness and headaches. 10 Tetrahydroaminoacridine (Tacrine), a cholinesterase inhibitor, has been approved for AD treatment but is hepatotoxic. 9
Thus, regarding AD drug discovery, it is necessary to identify other effective therapeutic drugs for AD treatment that will not induce severe adverse effects. Many modern drugs have natural origins (e.g., herbs that have a long history of proven safety and efficacy). 11 Artemisinin is a well-known natural drug derived from the plant Artemisia annua; it is used to treat malaria and has been commonly used in Chinese traditional medicine. 12
Further, artemisinin can be produced from metabolically engineered microorganisms instead of being extracted from plants; this reduces the cost of production. 13,14 Some herbs and herbal compounds have also been found to possess therapeutic benefits for AD treatment because of their anti-inflammatory, antioxidative, and neuroprotective properties.
For example, Panax ginseng, Gingko Biloba, and Salvia officinalis have been found effective in the treatment of AD symptoms through in vitro/in vivo experiments or clinical studies. 15 –17 However, the relationship between the herbs and their therapeutic effects has not been scientifically investigated to a sufficient degree.
This study introduced a simple but practical in silico approach for identifying effective herbs for the treatment of AD symptoms. To this end, we collected potential therapeutic target proteins of AD from a database and searched for herbs that contain natural products similar to the approved drugs modulating the potential targets. Using an in silico approach, we identified Corydalis ternata Nakai as a potential therapeutic herb for AD treatment. C. ternata was fed to AD mice (5XFAD) and evaluated for its therapeutic effects based on behavioral and molecular analyses.
Materials and Methods
Datasets
Natural herbs and their compounds were obtained from a database of natural products—COCONUT. 18 Therapeutic protein targets and known drugs were downloaded from DrugBank 19 and the Therapeutic Target Database (TTD). 20 AD-related proteins were obtained from Neurocarta. 21
The Tanimoto similarity between herbal compounds and known drugs
To calculate the chemicophysical similarity between a herbal compound and a drug, a molecular fingerprint representing the molecular structure was introduced. The extended connectivity fingerprint, a string of 1024 bits (1 or 0), was calculated using Dragon software (www.talete.mi.it/products/dragon_description.htm).
22
The Tanimoto similarity
23
was calculated using the fingerprint bits as follows:
where a denotes the number “1” in the fingerprint of a natural compound, b denotes the number “1” in the fingerprint of a drug, and c denotes the number “1,” which is at the same position as that of the fingerprints. If the similarity was below 0.45, the compounds were considered different and not used for herb identification.
Herb score
To identify herbs that are therapeutically effective for AD treatment among various herbs, we hypothesized that compounds with similar physicochemical properties are likely to target the same protein and this approach, known as quantitative structure activity relationship, is widely accepted in ligand-based drug discovery to design new chemical compounds binding to the same target protein. 24,25
This implies that compounds resembling those known to target AD-related proteins could also target the same proteins, and thus suggests that herbs containing such similar compounds could potentially be effective for AD treatment. Consequently, we devised a herb score, H(m) to quantify such potential. This score is calculated using the Tanimoto similarity between the compounds found in an herb and drugs known to target AD-related proteins. Here, m denotes an herb, and the herb-score formula is as follows:
c(m,i) denotes a compound (i) contained in a herb m, and d(j,k) denotes a drug (j) interacting with an AD-associated protein k. Thus,
5xFAD mice and treatment
Many experimental mouse models have been used for investigating potential therapeutic strategies for treating AD. 26 Given the unclear etiology of AD, 5XFAD transgenic mice expressing five AD-linked mutations (K670N/M671L [Swedish], I716V [Florida], and V717I [London] in APP, and M146L and L286V in PSEN1) have been widely used. 27,28 The 5XFAD mice were purchased from Jackson Laboratory (Bar Harbor, ME). Four to five mice were housed in cages and acclimated for 10 days before the herb administration.
Wild-type and 5XFAD mice were divided into three experimental groups: 5XFAD mice fed a regular normal diet (AD-C, n = 15), 5XFAD mice fed a C. ternata herb diet (AD-H, n = 15), and wild-type mice fed a normal diet (WT, n = 16). The regular normal diet was purchased from DBL Co. (Eumseong, Korea), and the C. ternata diet was a regular normal diet containing 5% C. ternata powder (5 g of C. ternata powder per 100 g of normal diet).
Dried root powder of C. ternata was purchased from the Gyeongdong market (Seoul, Korea). Dietary treatment was started at ∼5–6 months of age, and behavioral tests began at ∼8–9 months of age. Food intake and body weight were measured once in 2 weeks. Every group was kept in standard conditions at a temperature of 22°C ± 1°C and humidity of 50% ± 5% with 12 hours of light/dark cycle and free access to food and water. All mice were used according to the guidelines of the Sejong University Institutional Animal Care and Use.
Behavioral tests
Hind limb clasping test
The hind limb clasping test is a motor coordination measurement test. 29 The mice were suspended by their tails for 15 seconds. Their clasping score was estimated manually from 0 to 429; the scoring was as follows: 0 when a mouse splayed its hind limbs and toes; 1 when one hind limb was not splayed; 2 when its two hind limbs and toe were not splayed; 3 when both limbs were retracted to the abdomen; and 4 when forelimbs grabbed the hind limbs and curled up. The clasping test was conducted in 9-month-old mice (early stage of AD) and 12-month-old mice (late stage of AD).
Morris water maze test
The MWM is a spatial learning and memory test. In an open swimming arena (stainless steel; diameter: 91 cm; depth: 45 cm), the test comprised 6 consecutive days, including a training day. A hidden invisible platform (round shaped; diameter: 6 cm) was installed 1 cm below the water surface, and white-colored water (temperature: 23°C) was filled in the pool. The swimming arena had three cues. A mouse was placed at one of four locations (North, South, East, and West), and the time spent to find the hidden invisible platform was measured.
The tests were conducted using four trials, with each trial starting at four locations. If the mouse could not find the platform within 1 minute, it was placed on the platform for 15 seconds to memorize its location. After each trial, the mouse rested for at least 1 minute. To avoid human error, every test was recorded as a video, and the video was analyzed to calculate the time spent using the Any-maze software (Stoelting, Wood Dale, IL). The water maze test was conducted on 8-month-old (early stage of AD) and 12-month-old mice (late stage of AD).
RNA isolation and quantitative real-time PCR analysis
Total RNA was extracted using TRIzol reagents (Invitrogen). The total RNA concentration was quantified using a Nanodrop spectrophotometer (ND-2000 Spectrophotometer; Thermo Fisher Scientific). RNA was reverse-transcribed to cDNA using a high-capacity RNA-to-cDNA kit (Thermo Fisher Scientific), and cDNA was used for real-time PCR.
The real-time PCR was conducted using the QuantStudio™ 1 Real-Time PCR Instrument (Thermo Fisher Scientific) and PowerUp™ SYBR™ Green Master Mix (Thermo Fisher Scientific). Relative quantities (ΔΔCt) were obtained by normalizing their quantities using GAPDH as a reference. The primer sequences used for quantitative real-time PCR are listed in Table 1.
Primer Sequences for Quantitative Real-Time Polymerase Chain Reaction
Western blotting
Tissue samples were suspended in RIPA buffer containing 100 × Halt™ Protease Inhibitor Cocktail (Thermo Scientific) and 100 × Xpert Phosphatase Inhibitor Cocktail Solution (GenDEPOT) and then homogenized using T18 digital ULTRA TURRAX homogenizer (IKA). The homogenized samples were incubated on ice for 30 minutes and centrifuged at 4°C for 10 minutes at 13,000 rpm; their supernatants were collected for protein analysis.
The protein concentration was quantified using a BCA assay kit (Abbkine); 1 mg/ml of total protein lysate was suspended with 5 × sample buffer and RIPA buffer and boiled at 96°C for 5 minutes. Protein samples were separated using sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto polyvinylidene difluoride membranes. After employing membrane blocking with 5% skim milk in Tris-buffered saline with 0.1% Tween 20 detergent (TBST) buffer, the membrane was incubated at 4°C overnight with the primary antibodies listed in Table 2.
Antibodies Used for Western Blotting
After washing with TBST buffer, the membrane was incubated with an HRP-conjugated secondary antibody (Table 2) for 2 hours at room temperature (RT) and visualized using Fusion Solo (Vilber Lourmat). Band intensities were quantified using the ImageJ software (National Institutes of Health) and normalized using β-actin as a reference.
Immunohistochemistry
The brain samples were fixed in 4% paraformaldehyde (PFA; Biosesang) at 4°C for 16 hours and incubated with 30% sucrose in PBS buffer at 4°C until the samples were sunk. They were embedded in an optimal cutting temperature (OCT) compound (Sakura Finetek), mixed with 30% sucrose in PBS buffer, and stored at −80°C. These OCT-embedded brain samples were stored at −20°C for 24 hours and then cut into 30 μm-thick slices using a cryostat microtome (Thermo Scientific).
Sliced samples were stored in the cryoprotectant buffer (300 g/L of sucrose, 10 g/L of PVP40, 500 mL of PBS, and glycerol) at −20°C until use. For immunohistochemistry, a sample in a cryoprotectant buffer was washed twice with PBS buffer. The sample was soaked in 10 mM sodium citrate buffer (pH 8.5) and incubated in an 80°C oven for 30 minutes for antigen retrieval. After antigen retrieval, the sample was consecutively washed with PBS, PBS with 0.1% of Triton-X100, and PBS with 0.4% of Triton-X100.
For blocking, the sample was treated with PBS buffer containing 10% of donkey serum and 0.1% of Triton-X 100 for 3 hours at RT. The sample was incubated with a primary antibody dissolved in PBS buffer containing 5% of donkey serum and 0.1% of Triton-X 100 at 4°C overnight and incubated with a secondary antibody dissolved in PBS buffer containing 5% of donkey serum for 3 hours at RT.
After the secondary antibody incubation, the sample was stained using Hoechst solution (Thermo Scientific) for 10 minutes and washed twice with PBS containing 0.1% of Triton-X100, washed twice again with PBS, and rinsed with 100 mM sodium phosphate (PB buffer; Biosesang) before the sample was placed on a glass slide. The samples were covered with a mounting medium (Fisher Chemical) and a cover glass. The slide glass was dried for 2 hours at 4°C before observation.
Thioflavin-S staining
For thioflavin S staining, a sliced sample was placed on glass slides and dried at RT. The glass slide was washed with 70% and 80% ethanol (each for 1 minute) and incubated for 15 minutes with 0.4% thioflavin S (Sigma-Aldrich) dissolved in 80% ethanol. The glass was washed with 80% and 70% ethanol (each for 1 minute) and rinsed with deionized water. It was incubated with Hoechst staining solution for 10 minutes and covered with a mounting medium (Fisher Chemical) and a cover glass. The glass slide was dried for 2 hours at 4°C before observation. The number of amyloid plaques and thioflavin S+ areas were graphed using the ImageJ software (NIH ImageJ).
Statistical analysis
The statistical tests were conducted on three groups [WT control (WT), AD mice (AD-C), and AD mice fed with C. ternata (AD-H)] using one-way ANOVA with Tukey's honest significant difference (HSD) test, a post hoc test, since ANOVA test does not provide which specific groups are different from each other. If ANOVA test's p-value is less than the significance level, Tukey's HSD test is conducted on every pair of groups to determine which specific groups shows significant difference. A p-value of <0.05 was considered statistically different.
Values of p < 0.05 and p < 0.005 are represented with * and **, respectively. Outliers outside the interquartile range were excluded from the behavioral test results. GraphPad Prism software (version 8.0, GraphPad Prism) was used for statistical analysis.
Results and Discussion
Datasets
This study used an in silico approach for identifying potential herbs (Fig. 1A). We hypothesized that if a herb contained many compounds similar to known drugs targeting AD-related proteins, it would be effective for AD treatment. To test this hypothesis, we collected herbs and their compounds along with drugs and their target proteins using various data sources. Natural herbs and their compounds were collected using the COCONUT database. 18

The overall in silico process for identifying potential effective therapeutic herbs for AD treatment.
Among the herbal compounds, to discard compounds non-absorbable in the intestine and non-permeable in the blood-brain barrier (BBB), the permeabilities were predicted using ADMETlab 2.0, and LightBBB. 30,31 We obtained 4183 herb species and 7851 herbal compounds. As the number of known compounds per herb varied, for consistent analysis results, we merged the compounds of the herb species into their respective genera and consequently obtained 1626 genera of herbs for use in this study.
Drugs and their target proteins were obtained from DrugBank and TTD: 1434 therapeutic target proteins and 9095 drug compounds. 19,20 In total, 397 AD-related proteins were obtained from Neurocarta, 21 and 127 were included in DrugBank and TTD, indicating that they had been used as therapeutic targets and that drugs modulating these proteins were available. Potential therapeutic herbs for AD treatment were identified using the 127 AD-related proteins and the drugs targeting these proteins.
Herb score calculation
Natural herbs contain numerous compounds that can target multiple proteins. This multi-component and multi-targeting approach is beneficial for treating diseases because multiple compounds can modulate the proteins in the same pathway and/or modulate several functions in an orchestrated manner. Thus, we calculated herb scores as described in Materials and Methods (Fig. 1B), assuming that a herb containing many compounds similar to drugs targeting AD-related proteins will get a higher herb score.
For the given herb and AD-related protein, the pairwise Tanimoto similarities of its compounds with drugs targeting the protein were calculated, and the maximum similarity was taken as a score for the AD-related protein. If the similarity was lower than 0.45, the similarity was discarded. This pairwise similarity calculation was repeated for all AD-related proteins, and the sum of all scores was taken as the herb score. As the number of discovered compounds varied from one herb to another, the herb score was normalized to the number of compounds in a given herb, and this normalized score was used as the per-compound herb score.
Of the collected 1626 natural herbs, 1014 herbs had a herb score higher than zero. The remaining herbs had scores of zero, indicating that their compounds had no significant similarity to known drugs that interact with AD-related proteins. Thus, only 1014 herbs were used for further analyses.
Figure 1C lists the 15 herbs with the highest herb scores. Of the top-rated herbs, Corydalis grabbed our attention because of its high herb score and the relatively large number of associated effective species. As the herb scores were calculated at the genus level, we also investigated how many species contained at least one or more compounds similar to drugs targeting AD-related proteins. There were 25 Corydalis species that contained compounds similar to known AD protein-targeting drugs (Fig. 1C, white).
The second and third most effective herbs were Sophora and Myristica, but the number of associated effective species was lower than that of Corydalis. This result indicates that the Corydalis species may contain potential therapeutic compounds that are common across species. Owing to this similarity, it is possible to discover a common biosynthetic pathway for synthesizing compounds and utilizing them for metabolic engineering. 32 Consequently, we selected Corydalis through in silico analysis—specifically, one of the Corydalis species available in Korea, C. ternata, which has not been previously evaluated.
Potential therapeutic Corydalis compounds similar to approved drugs
Table 3 lists several Corydalis compounds similar to known drugs that inhibit AD-related proteins, such as phosphodiesterases, acetylcholinesterases, TLRs, 5-hydroxytryptamine receptors, and estrogen receptors. Thus, those Corydalis compounds may serve as potential inhibitors of AD-related proteins. Phosphodiesterases are a family of enzymes involved in the regulation of cAMP and cGMP in intracellular signaling and are widely expressed in the brain.
Corydalis Compounds Similar to Known Drugs Targeting Alzheimer's Disease-Associated Proteins
Phosphodiesterase inhibitors showed improved memory, learning, and cognition in mice with AD when cellular gene expression and protein distribution were altered. 33,34 Cholinergic abnormalities, such as a decreased number of acetylcholine receptors, form one of the major characteristics of AD. 35,36 Thus, inhibition of acetylcholine downregulation is one of the strategies for treating AD. Acetylcholinesterase inhibitors, which suppress the degradation of released acetylcholine by acetylcholinesterase, are used for treating AD. 37,38
Toll-like receptors (TLRs) can reportedly contribute to the pathogenesis of age-related AD. 39,40 The expression of TLR4 in aged or neurodegenerative neurons increases, and brain cell damage or infection may activate TLR-mediated Ca2+ signaling, leading to apoptosis. A reduction in TLR4-mediated signaling decreases the risk of late-onset AD. 41 The 5-hydroxytryptamine receptors are highly expressed in the brain tissue of mice and patients with AD, and inhibition of these receptors partially reverses cognitive deficits, reduces Aβ plaques, and decreases neuroinflammation in mice. 42
Estrogen receptors are also neuroprotective against AD. 43 Several mechanisms can be involved in the protective role of estrogen receptors, including estrogen receptor-mediated calcium signaling, inhibition of apoptosis, and prevention of glutamate excitotoxic injury by Aβ. Metalloproteinases are endopeptidases produced by neurons and glial cells. Certain metalloproteinases can degrade Aβ, and their expression is stimulated in response to Aβ.
Effects of C. ternata on motor and memory functions of AD mice
To confirm the effectiveness of C. ternata for AD treatment in terms of motor coordination and memory impairment, a regular food mixed with C. ternata was fed to 5XFAD mice that harbored five key mutations: K670N/M671L (Swedish), I716V (Florida), and V717I (London) within APP and M146L and L286V within PSEN1. 28
To test the potential toxicity of C. ternata, we fed mice with two different concentrations of C. ternata, 5% and 10%. As shown in Figure 2A, 5% C. ternata showed no difference in body weight compared with normal mice and 10% C. ternata showed slightly decreased body weight. Though both concentrations showed no statistically significant difference from normal mice (p > 0.3), due to the slightly reduced averaged body weight by 10% C. ternata, we fed mice with 5% C. ternata in this study.
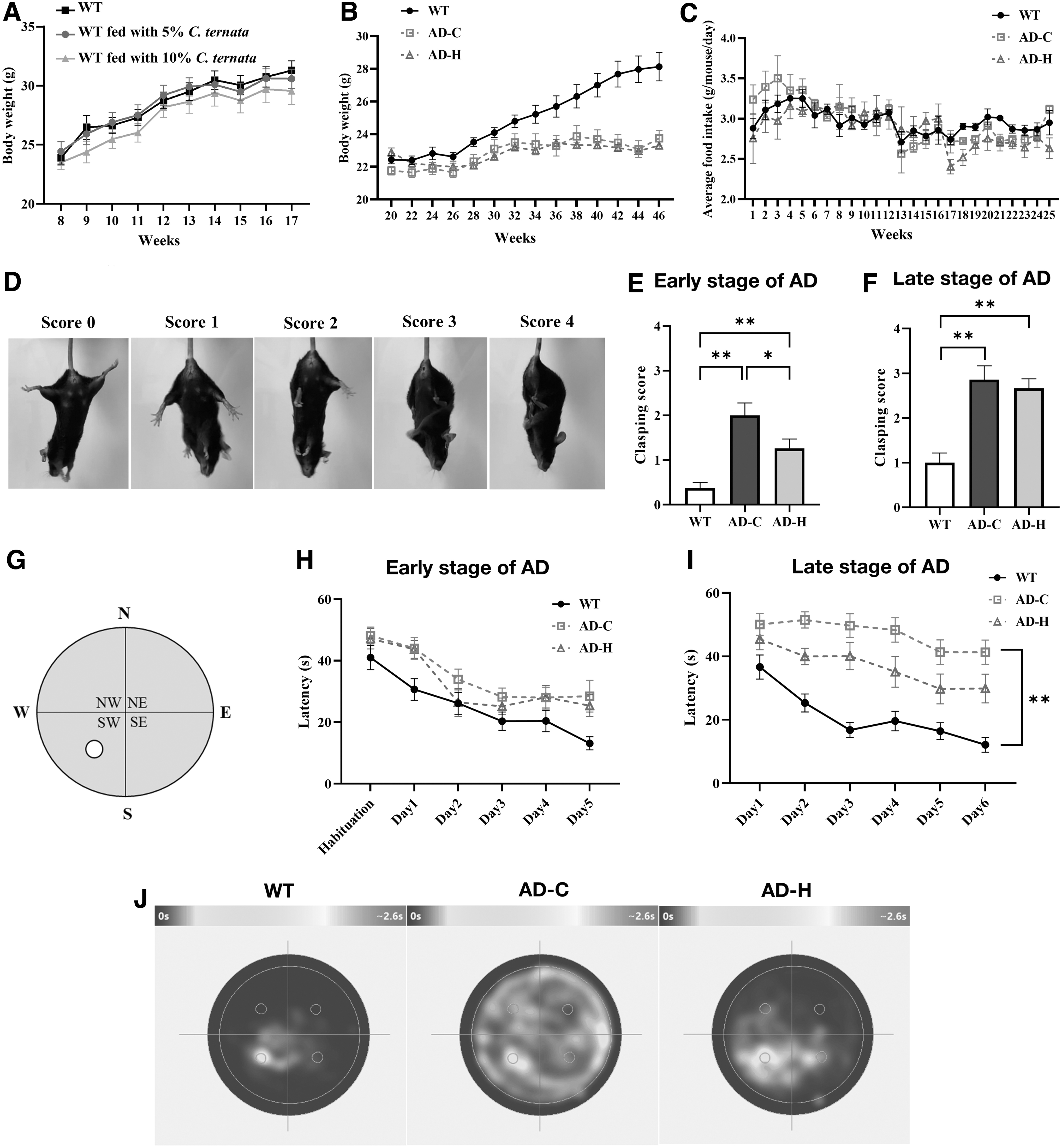
Effect of Corydalis ternata on AD-related behaviors.
In Figure 2B, C, AD mice showed decreased body weight compared with normal mice, but there was no difference in body weight or food intake between AD mice fed with the normal diet (AD-C) and AD mice fed with 5% C. ternata (AC-H), indicating that C. ternata had no effect on body weight or food intake in AD mice.
To investigate motor coordination, we conducted a hind limb clasping test in 9-month-old (early-stage AD) and 12-month-old (late-stage AD) mice. An example clasping score for the mouse is shown in Figure 2D. Healthy mice normally show a score of 0, with a higher score indicating decreased sensorimotor function. First, the test was conducted in mice in the early stages of AD. As shown in Figure 2E, AD mice showed higher clasping scores than WT mice, indicating impaired motor function in AD mice.
When C. ternata was fed to the mice, the clasping score significantly decreased, indicating an improvement of motor function in AD mice. Although AD mice fed with C. ternata showed motor function impairment compared with WT mice, there was a profound recovery of motor function (AD-C vs. AD-H in Fig. 2E). When tested with mice in the late stage of AD, there was no significant difference in the clasping score (Fig. 2F). These results indicate that C. ternata improves the motor activity of AD mice only at the early stage of AD.
Similar to the clasping test, the Morris water maze test was performed on 8-month-old mice (early stage) and 12-month-old mice (late stage) to test learning and memory. The water maze is illustrated in Figure 2G, where the white circle denotes the hidden platform underneath. The Morris water maze test measures the time required to find a white platform from a random location. If the mouse learns and memorizes the location of the platform, it quickly finds the platform. Otherwise, more time is taken to find the platform.
As shown in Figure 2H, I, the time spent to find the platform (latency) decreased over time because the mice learned the location of the hidden platform. Interestingly, unlike the motor coordination test, C. ternata showed no significant improvement in the memory of mice in the early stage of AD (Fig. 2H). When the same mice were tested again at the late stage of AD (Fig. 2I), AD mice took more time to find the hidden platform than WT mice, indicating that they suffered from severe memory impairment.
In AD mice, those fed C. ternata took less time to find the platform, and the latency decreased over time, indicating that C. ternata improved learning and memory functions in AD mice. To provide an intuitive representation of the Morris water maze test results, the color-coded moving maps on Day 6 for mice in the late stage of AD are shown in Figure 2J. Cyan denotes where and for how long a mouse was swimming. The WT mice quickly found the hidden platform and spent most of their time around it because they memorized its location.
However, as the AD mice (AD-C) could not memorize the location, they randomly staggered around the water maze. AD mice fed with C. ternata (AD-H) spent more time close to the platform than AD mice fed with the normal diet (AD-C), indicating that C. ternata could improve memory function in AD sufferers. Consequently, C. ternata could be beneficial for memory improvement for late-stage AD sufferers; nonetheless, C. ternata may require long-term treatment to show beneficial effects on memory function.
Beneficial effect of C. ternata on pathology
The pathogenesis of AD is associated with loss of synapses and decreased synaptic plasticity related to memory formation. 44 To investigate how C. ternata alleviates the pathology of AD, mouse brains were analyzed after behavioral tests for protein aggregates and neuronal plasticity. When the level of Aβ—a representative pathology of AD—was measured, it was halved in the brain of a mouse fed with C. ternata compared with that of a control AD mouse (Fig. 3A). This indicates that C. ternata can decrease the number of aggregates. We also investigated the expression of the postsynaptic density protein-95 (PSD95), a postsynaptic marker of synaptic plasticity.

Effect of C. ternata on AD pathology and neuronal plasticity.
Neuronal plasticity involves a rewiring of neuronal circuits and changes in the function of the brain. 45 Specifically, synaptic plasticity plays a role in memory function, and its dysfunction is related to neuropsychiatric disorders. As the pathology progresses in AD mice, the PSD95 level gradually decreases over time. 46 Consistent with a previous report, the levels of PSD95 in AD mice (AD-C) were lower than those in WT mice (WT) (Fig. 3A). However, C. ternata did not improve synaptic plasticity. Consequently, C. ternata can decrease Aβ accumulation but cannot improve synaptic plasticity significantly.
For further investigation, two parts of brain tissues (the cortex and hippocampus) were stained with thioflavin S to identify Aβ plaques (Fig. 3B). The cortex is associated with neuropsychiatric symptoms such as depression and anxiety, 47,48 and the hippocampus is associated with memory function. 49 AD mice showed Aβ plaques in the cortex and the hippocampus. When fed C. ternata, the level of plaques in the hippocampus decreased by 2.1-fold, but there was no statistical difference in the cortex. Considered together, C. ternata can reduce Aβ in the hippocampus; this suggests that reduced Aβ in the hippocampus results in the recovery of memory function.
Effect of C. ternata on neuronal apoptosis, neuroinflammation, and autophagy
Pathological changes such as apoptosis, neuroinflammation, and autophagy-dependent cell death are the main features of neurodegenerative diseases such as AD. 50 Neuroinflammation plays a critical role in the development and progression of AD. This occurs primarily through the chronic activation of the brain's immune cells, such as microglia and astrocytes, in response to Aβ plaques. 51 This chronic activation leads to the prolonged release of pro-inflammatory cytokines.
This exacerbates neuronal damage, synaptic dysfunction, and cognitive deficits characteristic of AD. Moreover, neuroinflammation can impair the clearance of Aβ, which promotes further neuroinflammation and neuronal death. Firstly, to investigate whether C. ternata could recover the number of neurons and inhibit neuroinflammation, we analyzed neuronal (neuron-specific nuclear protein, NeuN) and astrocyte markers (glial fibrillary acidic protein, GFAP), which are proportionally related to neuron number and neuroinflammation, respectively. 52 NeuN is a nuclear protein expressed in neuronal cell types throughout the nervous systems of adult mice. 53 GFAP is a type III intermediate filament protein expressed in the cells of the central nervous system, including astrocytes.
These markers are commonly found in AD mice and patients and are related to brain function. 51,54 As shown in Figure 4A, the level of NeuN decreased in AD mice, indicating that the number of neurons decreased significantly. Furthermore, the number of astrocytes dramatically increased, indicating increased neuroinflammation. However, the administration of C. ternata to AD mice resulted in a significantly reduced number of astrocytes (GFAP), indicative of decreased neuroinflammation, although no significant improvement in the neuronal population (NeuN) was observed (Fig. 4A).
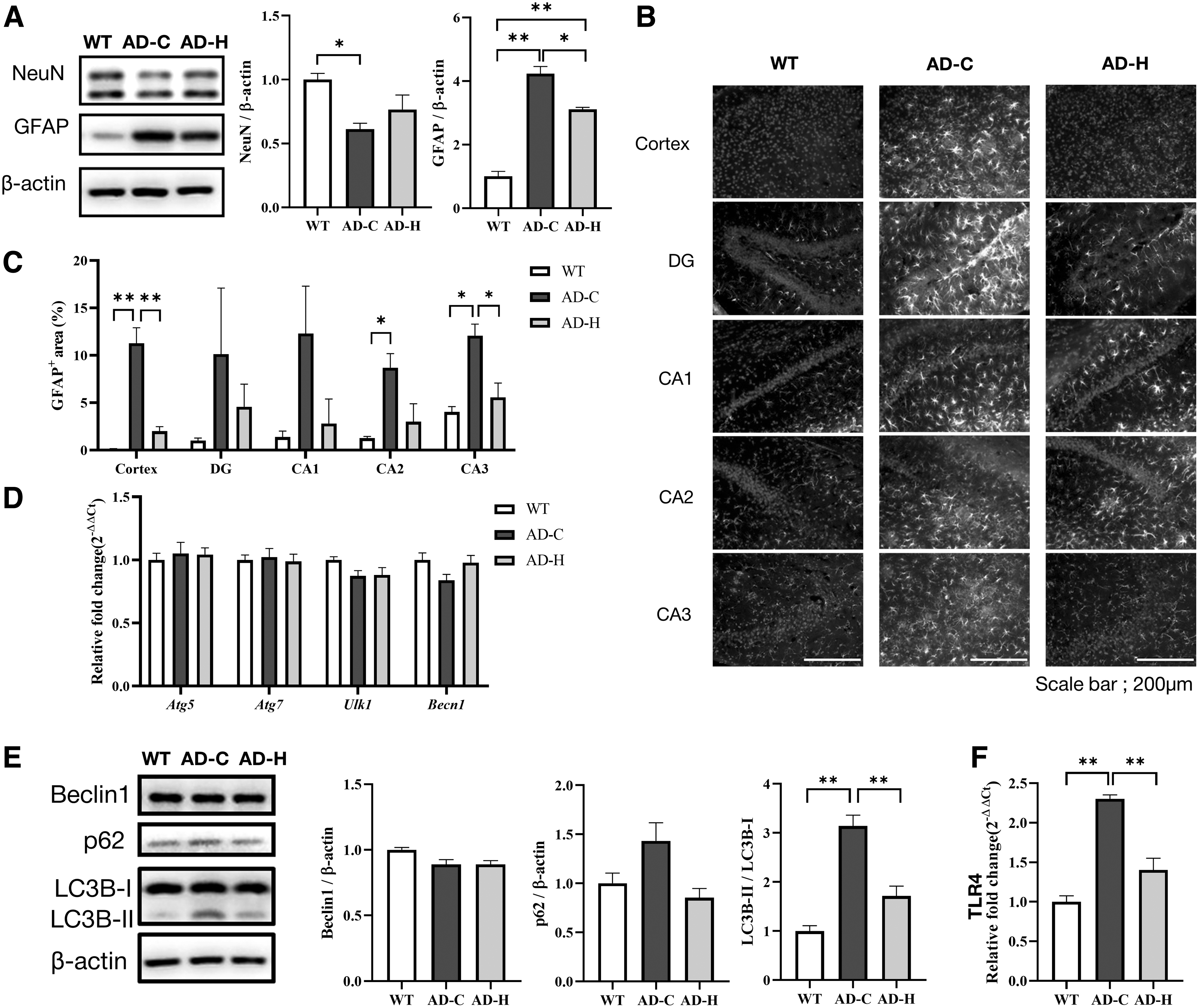
Effect of C. ternata on neuron population, neuroinflammation, and autophagy.
We further investigated GFAP distribution in the cortex region and four subregions of the hippocampus (dentate gyrus; DG, Cornu ammonis 1–3; CA1, CA2, and CA3). We found that AD mice showed extensive neuroinflammation in these brain regions (Fig. 4B, C). Interestingly, inflammation was significantly reduced in the cortex and hippocampus when fed with C. ternata. This finding suggests that C. ternata ameliorates learning and memory function by modulating neuroinflammation, not neuronal death.
Autophagy, a cellular process that eliminates cellular abnormal or damaged proteins, is a key regulator of Aβ generation and clearance in AD, since this process removes Aβ aggregates, thereby potentially slowing down the disease's progression. The decreased expressions of autophagy-related genes were observed in the brains of AD mice or patients, and the reduction or knockout of the autophagy genes resulted in accumulation of Aβ and neurodegeneration. 55 –58
For example, Beclin1 (Becn1) mediates the initiation of autophagy, and its reduced expression was found in AD. 56 Becn1 is also involved in neuroinflammation and thus its reduced expression increases microglia inflammatory response. 59 Therefore, impaired autophagy can lead to increased accumulation of this toxic protein, contributing to neuronal damage, inflammation, and neurodegeneration observed in AD. Enhancing autophagy could offer a potential therapeutic strategy for AD treatment by improving the clearance of pathological proteins. 60
Unlike our expectation, as shown in Figure 4D, there were no significant differences in the levels of autophagy-related genes involved in autophagy initiation (Atg5, Atg7, Ulk1, and Becn1), indicating that C. ternata did not affect autophagy initiation. Thus, we shifted our focus on the overall autophagic flux by investigating LC3 gene family and p62, because AD mice showed increased p62 levels and LC3B-II/LC3B-I ratios but C. ternata recovered to the levels close to those observed in normal mice (Fig. 4E)
The increased levels of LC3 proteins and p62 indicate that autophagic flux did not flow efficiently, leading to cargo accumulation. The LC3 gene family encodes structural proteins (LC3A, LC3B, and LC3C) of the autophagosome membrane and p62 acts as a cargo receptor recognizing ubiquitinated proteins for degradation. When autophagy begins, autophagosomes engulf the cytoplasmic target components to be degraded.
Simultaneously, a cytosolic form of LC3 (LC3-I) is converted to an LC3-phosphatidylethanolamine conjugate (LC3-II), which is recruited to the autophagosomal membrane. 61 The adapter protein p62 binds to the LC3-II recruited to the autophagosomal membrane and transports ubiquitinated proteins and organelles for degradation. The autophagosomes formed by p62 and LC3B are bound to lysosomes to decompose their contents. 62 If autophagosomes are not degraded normally, p62 and LC3-II accumulate. 63 Taken together, the recovery of autophagic flux may be related to the decrease in amyloid plaques in the C. ternata group, though C. ternata did not affect autophagy initiation.
This study confirmed the alleviating effects of C. ternata on the pathology of AD mice. Specifically, C. ternata alleviated motor and memory impairment by decreasing Aβ, reducing neuroinflammation, and promoting autophagy flux. Considering these results, the applicability of our in silico approach was confirmed because C. ternata discovered through the in silico method actually showed an effect in alleviating the pathology of AD animal models. However, since our in silico method does not consider the effects of individual C. ternata compounds on AD pathophysiology, it should be investigated how the compounds modulate AD pathophysiology in future studies.
Investigation of the link between suggested potential target proteins and AD pathophysiology
We suggested potential target proteins of Corydalis compounds through our in silico analysis in Table 3. Of the potential target proteins, the TLR4 grabbed our attention because the transcription level of TLR4 was significantly increased in AD mice (AD-C in Fig. 4F). TLR4 is a type of protein playing a crucial role in the innate immune system. TLR receptors recognize molecules broadly shared by pathogens and self-derived molecules released upon tissue damage.
Thus, TLR4 is believed to mediate the neurotoxic actions of the self-derived molecules released upon neuronal damage 39 and, therefore, TLR4 modulates AD pathology by mediating inflammation, autophagy, and apoptosis. 64 Specifically, aggregated Aβ can bind to TLR4 on astrocytes and microglia. Microglia are immune cells in the brain and the Aβ-mediated activation leads to increased phagocytosis and production of cytokines, resulting in neuroinflammation. 65,66
In this study, the neuroinflammation induced by Aβ was reduced when fed with C. ternata (Fig. 4A), representing that TLR4 modulated by the suggested C. ternata compound would be one of the factors that alleviate AD and neuroinflammation. Interestingly, the transcription of TLR4 was also decreased when fed with C. ternata (AD-H in Fig. 4F). Therefore, the alleviation of AD pathophysiology was due to synergic outcome of TLR4 inhibition by the compound and TLR4 repression by another unknown compound. However, it should be elucidated further how C. ternata compounds modulate the transcription of TLR4. In addition, it should be also elucidated whether other target proteins are also modulated by C. ternata compounds.
Conclusion
In this study, our in silico approach successfully identified C. ternata as a promising candidate for AD treatment, demonstrating its therapeutic potential in alleviating cognitive decline through in vivo validations. This highlights the utility of integrating computational methods with traditional herbal medicine to discover new therapeutic agents for complex diseases such as AD. Despite the successes, our approach, relying on chemical similarity rather than more detailed but computationally intensive methods like molecular dynamic simulations, faces limitations in precision and comprehensiveness. Future enhancements by incorporating extensive datasets of herbal compounds and advanced in silico techniques promise to refine our understanding and identification of therapeutically potent herbs.
Moreover, while C. ternata's ability to mitigate the key pathological features of AD—autophagy and neuroinflammation—provides valuable insights into its action mechanism, the full therapeutic potential of C. ternata remains to be undiscovered. Future research should delve into the molecular basis of its effects, focusing on the identification and interaction of active compounds with AD-related targets. Such investigations are crucial for advancing our quest for effective AD treatments, illustrating the powerful synergy between traditional herbal medicine and modern scientific methodologies.
Footnotes
Authors' Contribution
Conceptualization, M.Y., D.N.; Methodology, M.Y., H.L, and D.N.; Validation, B.L, J.G.S.; Investigation, B.L., M.Y., J.G.S., and H.L.; Formal analysis, J.G.S., H.L., H.W.K., and D.N.; Writing—original draft, B.L., M.Y., and D.N.; Writing—review & editing, H.W.K., D.N.; Supervision, H.W.K., D.N.; and Funding acquisition, D.N.
Author Disclosure Statement
The authors declare that they have no competing financial interests or personal relationships that could have influenced the work reported in this article.
Funding Information
This research was supported by the Bio & Medical Technology Development Program of the National Research Foundation (NRF) and funded by the Korean Government (MSIT) (NRF-2022M3A9B6082687, NRF-2023R1A2C1008156, NRF-2021R1I1A1A01056588, and NRF-2021R1F1A1059574).
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.