
Abstract
The aim of this study is to analyze tree, as well as forest floor and topsoil (0–10 cm) changes, within a period of 37 years and related impact on forest growth in an oak ecosystem in northern Greece. The study was carried out in the area of the Taxiarhis University forest, which is located in central Halkidiki. In the experimental plot, the tree age was 70–72 years, the forest stand was healthy, and the site quality was medium. The parent material of the study area is schist and the topsoil is characterized by high organic matter content and a sandy clay loam texture. For the purpose of this research project, forest floor (O1 + O2) and soil (0–5 and 5–10 cm) samples were collected from 30 sampling points. To study the mycorrhizal colonization (%), 15 root samples were taken. The average tree height in 1981 was 14.2 m and in 2018, it was 19.3 m. The diameter increased by 4.05 and 5.40 cm within 17 (2001–2018) and 37 years (1981–2018), respectively, and the trees grew 5.1 m taller. Higher radial increment (4.2 cm/20 years) was observed only in the predominant trees. In 2018, the forest floor increased its weight by 18.273 kg/ha, compared with 37 years ago. Nutrient amounts increased and there was a balance between the quantity of organic matter on the forest floor and the topsoil. Most of the nutrient amounts in the forest floor and topsoil were higher in 2018 compared with 1981 and the acidity of the soil decreased. Moreover, most of the nutrient amounts were found much higher in the upper part of the topsoil (0–5 cm) than in the forest floor, except for phosphorus, iron, copper, and zinc. No significant statistical differences were found for the C/N ratio between 2001 and 2018. The study results highlight the importance of letting oak ecosystems grow and mature rather than cutting them down within the short period of 30–70 years. Also, the importance of coexistence, of ecosystems interconnections, the trees’ resilience to stressors such as drought, and the role of mycorrhizal colonization are addressed. Concluding comments address the significance of soil properties in relation to sustainability, especially in the context of the ongoing climate change.
Introduction
The role of the soil in ecosystems is of primary importance in the provision of nutrients and water, supporting plants and their growth, and regulating the water infiltration rates and water yield. There is a strong relationship between vegetative cover and soil condition. Vegetative cover exerts an important influence on soil (Grigal, 2000), whereas soil condition and characteristics also determine the condition of the vegetation. Nutrient availability determines survival, growth, and productivity of forests. The Mediterranean area is particularly affected by erosion (Pereira et al., 2017) with an associated loss of topsoil. The loss of organic-rich topsoil is regarded to be the biggest degrader of soil quality and, in most cases, cannot be restored (Alewell et al., 2015). This superficial part of the soil, the humipedon, is the most reactive to faunal and floristic changes and, for this, it is considered as a good indicator of ecosystem health and functioning (Ponge & Chevalier, 2006; Ponge, 2003).
Τhe distribution of nutrients in the ecosystem was found to be in larger quantities in soil but considerable quantities were stored in biomass. The forest floor constitutes a significant and constant supply of organic matter and nutrient elements (mineralization) for the soil and, therefore, for the ecosystems. The largest part of the annually absorbed nutrients from the forest vegetation, return to the soil as plant residues, in the form of leaves, needles, branches of various diameters, fruit, bark, etc., or by the permeable rain and crown outflow (Cole & Rapp, 1981). The quantity of forest floor nutrients entering the soil depends on the decomposition rates, the water quantity, and the nutrients’ form in the organic residues. In this way, vegetation, fungi, and water contribute in a particularly efficient way to the recycling of the nutrient elements in the forest ecosystems. Recycling is limited during the early developmental stages of a stand because the restoration of organic residues is low. However, it increases as the stand matures (Xia et al., 2021).
Higher site qualities are differentiated by abundant evidence of plant residues and resulting high concentrations of nutrient elements derived from these residues, relative to lower residues and nutrients, which define lower site qualities (Austin & Vitousek, 1998). The site quality has a great influence on the forest floors’ chemical composition, affecting the input of nutrient concentrations in the mineral soil through the rain. The site quality’s effect is attributed to the higher amount of forest floor, subsequent decomposition rates, and the differences in the rains’ chemical composition enriching the soil. During 1981, which is the first year of this comparison study, organic residues contributed 40.7 percent of their weight to the higher site qualities and 33.2 percent in the lower site qualities (at that time the study compared 3 plots-site qualities). Additionally, 67–86 percent of the mineral soils’ organic matter was found in the upper layer (0–20 cm), and at the lower site qualities there was less organic matter. Site quality also affects the amount of nutrients absorbed by the vegetation. At the best site qualities, the nutrient concentrations were found to be 34.8 percent nitrogen (N), 86.2 percent phosphorus (Ρ), 44.1 percent potassium (Κ), 38.2 percent calcium (Ca), and 48.1 percent magnesium (Mg) higher than in the lower ones (Alifragis, 1984).
The annual improvement of soil conditions, at the best site qualities in particular, further improved the site quality (feedback effect). Long-term tree—soil interactions can shape topsoil properties and organisms thus allowing for a potential indirect effect of tree species on leaf-fall decay via changes in soil environment (Aponte et al., 2010). Due to their ability to favor and improve tree growth and survival, the role of mycorrhizal networks is crucial in the restoration of degraded forest ecosystems (Itoo & Reshi, 2013). These organisms allow trees to use soil nutrients more efficiently and seedlings are favored even from the initial growth stages (especially in poor soils) given contributing protection from root pathogens (Branzanti et al., 1999). Additionally, the absorption of water and nutrients increases and the toxicity of some metals is restricted (Vasile et al. 2021).
There is an increasing interest in the use of forested areas for improving soil characteristics such as soil carbon (Aryal et al., 2013; Alvarez & Rubio, 2016) and soil nitrogen (Homyak et al., 2008). This topic has been also studied in Greece for Quercus frainetto by Alifragis (1984) and Ganatsios (2004). This study aims to enhance the knowledge of interactions and interconnections in natural forest (oak) ecosystems especially between nutrients, tree growth, mucorhizzal fungi, and drought resilience. Therefore, the study, which evaluates a 37 year period, is focused on the changes in the tree growth, in the forest floor and topsoil up to the depth of 0–10 cm, because the upper portion of the topsoil is more susceptible to observed changes in Mediterranean ecosystems (Gakis et al., 2003). Further, soil nutritional status was assessed in particular for organic carbon (C), N, pH, Ca, and P concentrations as these elements and attributes are important regulating factors of microbial activity and soil fertility (Andreetta et al. 2018; Salmon 2018). In the present literature, limited research exists on mycorrhizal colonization in Greek oak ecosystems, and this research aims to be the starting point of such measurements.
Forest restoration has become one of the most important challenges for restoration ecology in recent years and soil fungi are fundamental drivers of forest ecosystem processes, with significant implications for plant growth and survival (Adamo et al., 2022). Once trees are lost, the underground microorganisms do not recover quickly, even if more trees are planted (Kadowaki, 2023). The ability of oaks to respond to global warming will depend on the dual dispersal of acorns and of spores of mycorrhizal fungi (Southworth, 2013). Seedlings grow better when they inherit mycorrhizal fungi from parent trees living in symbiosis with the same mycorrhizal type (Kadowaki, 2023).
Materials and Methods
The Study Area
The study was conducted in the Taxiarhis University forest (TUF) (Fig. 1) which is located in central Halkidiki and is dominated by hardwoods, mostly Q. frainetto, representing 13 percent of the standing volume in Greece and the predominant tree species in the study plot. Tree age is 70–72 years (year 2022) and the forest stand is healthy. In the experimental plot (0.2 ha, 40 m × 50 m), (860 m altitude, 40°25′45.91″ N, 23°30′18.90″ E), the canopy closure reaches up to 100 percent. The site quality is medium. This distinction was based on the height of predominant trees (20 trees) in relation to their age and the soil depth (Dafis, 1966). The trees in the plot originated as sprouts and no silvicultural treatments have been applied in the plot during its conversion from coppice to high stand; therefore, it is a forest under protection. This plot is one of the control plots referenced by Ganatsios et al. (2021).
The study area
The study area has a typical temperate mesothermal (subtype CSb), according to the Köppen classification system. Moderate temperatures (average annual air temperature 11.4°C) and rainy weather prevail during the winters, whereas summers are characterized by a short hot and dry period. Long-term average annual precipitation is 756 mm, which mainly occurs from October to March. All climatic data are collected at the TUF weather station (above sea level 860 m), located only 150 m away from the boundaries of the study plot (exposure South East).
The forest floor (O1 + O2), comprised of both the O1 (litter layer composed of leaves and branches) and the O2 (decomposed organic material), has an average depth of 4–5 cm which increases infiltration and decreases overland flow (Fig. 2). The decomposition rate is a result of favorable temperature and moisture conditions (Ganatsios et al., 2021). The H layer (humus layer) follows O2 and consists of well-decomposed organic material (Fisher & Binley, 2000). It is thinner with an average depth of 0.5–1 cm and, in the context of our study and for practical reasons, it has been included in O2 layer. Once the forest floor is removed, the revealed topsoil is characterized by a high organic matter content and a sandy clay loam texture.

Soil parent material is schist (mica and talk). The soil has a subangular blocky structure. The soil belongs to the chromic luvisols classification (IUSS Working Group WRB, 2006) and could be described as immature due to a history of intense erosion. More specifically, the protective vegetation cover has degraded, as a result of successive land uses in the past, due to intensive wood harvesting, grazing, and use of the area as agricultural land until the 50s. Then, the area was restored to its forest land state, and currently the soils are well protected from erosion by the existing vegetation and forest floor. Soil has an average depth of 45–60 cm in the upper part of the plots and gets deeper (>90 cm) only at the lower part of one plot.
Sampling and Data Collection
The height and the diameter at breast height of each tree in the plot having a diameter larger than 4 cm were measured during the summer of 2001 and 2018. During the summer of 2022, due to illegal harvesting of firewood, a few trees were cut down. Due to this incident, an opportunity was given to measure the annual diameter increment and also compare it with the same nearby trees, studied by E.P (unpublished study). The study plot noted for this evaluation was previously used for a 2-year study, 1981 and 1982 (Alifragis 1984), thus providing the opportunity to compare with the present study and have an evaluation period of 37 years. Alifragis (1984), measured tree diameter and took samples in July of 1981 and 1982 without finding important differences in soil properties between these years. Considering that the thickness and the quantity of forest floor varies according to season (Paudel et al., 2015), the sampling was conducted during the same month (July 2018).
For the present study, systematical sampling was carried out with the use of a sampling grid (Köhl & Magnussen, 2016). Thirty sampling points were selected and the forest floor samples were obtained by pressing a square steel sheet sampling frame (25 cm × 25 cm = 625 cm2, 10 cm deep) to delimit the sampling points. From the same sampling points, soil samples were also collected. More specifically, at each sampling point, (1) the depth of each organic layer was measured (O1 and O2) and all the litter from each organic layer was collected separately in plastic bags, and (2) samples at soil depths 0–5 and 5–10 cm were put in separate plastic bags, labeled and transported to the laboratory. During the 2001–2003 period and due to various studies (Ganatsios, 2004), values of some soil properties are also available for comparison. To study the mycorrhizal colonization, 15 root samples were taken.
Laboratory Measurements
A stereoscope was used to measure the annual radial increment. The chemical analyses of both forest floor and soil samples were conducted in the Laboratory of Forest Soil, School of Forestry and Natural Environment, Aristotle University of Thessaloniki. Forest floor samples were dried in the oven at 75°C for 48 h and weighed. Then, the forest floor samples were ground in a Willey mill to pass through a 40-mesh stainless steel sieve to obtain a powdered and homogenized material, which were then subsequently stored in small plastic containers. In the forest floor samples, the total N was determined with the Kjeldahl method (Stevenson and Stevenson, 1982). Macronutrients [potassium (K), Ca, magnesium (Mg), and sodium (Na)] and micronutrients [iron (Fe), mManganese (Mn), copper (Cu), and zinc (Zn)] were measured by atomic absorption spectroscopy in a solution after the disorganization of the powdered forest floor samples with HNO3+HClO4+H2SO4(5:1:0.5) (Allen et al., 1986). In the same solution, P was determined by the Molybdenum Blue Method (Olsen & Sommers, 1982). The organic matter of the forest floor samples was determined with the loss on ignition method at 515°C for 4.5 h.
Soil samples were air-dried and sieved through 2 mm screens and stored in numbered plastic containers. Soil pH was determined by using a glass electrode in a soil-water suspension (1:1, by weight) (Mc Lean, 1982). Organic matter was determined by the wet oxidation method (Nelson & Sommers, 1982) and organic N was determined by the Kjeldahl method (Stevenson, 1982). Extractable P was measured by the Molybdenum Blue Method, using a Shimadzu spectrophotometer model UV-1201 V (Olsen and Sommers, 1982). Exchangeable cations (K, Ca, Mg, and Na) were determined after extraction of soil with CH3COONH4, pH 7 (Grant and Page, 1982), and micronutrients (Fe, Mn, Cu, and Zn) were determined after extraction of soil with DTPA, pH 7.3, by atomic absorption spectroscopy (Lindsay & Norvell, 1978). Mycorrhizal colonization measurement were based on the study by Phillips & Hayman (1970).
Statistical Analysis
A one-way ANOVA with Tukey post hoc test was used to assess the significance of observed differences in the average values of the year 2018. The statistical analyses were conducted with the SPSS software version 23. Significance level was set at 0.05. t-test have been used. Mann–Whitney U test (non-parametric test) was also used to compare the soil properties results of the years 2001 and 2018 for the two subgroups of topsoil, 0–5 cm and 5–10 cm.
Results
Tree Growth Results
The maximum tree height of the experimental plot measured in 1981 and 2018 was 15.1 and 22 m, respectively. The average height in 1981 was 14.2 m [standard deviation (SD) = 4.216] and in 2018, 19.3 m (SD = 5.385) respectively. Mean diameter increased from 12.5 cm (SD = 5) (1981), to 13.85 cm (SD = 2.5) (2001) and 17.9 cm (SD = 5.0) (2018). So, the diameter increased by 4.05 cm and 5.4 cm within 17 (2001–2018) and 37 years (1981–2018), respectively. The low average radius increment (1.3 cm/20 years) found by comparing the diameter values, also have been verified by stereoscopic measurements. Higher radial increment (4.2 cm/20 years) was observed only in the predominant trees. Maximum average diameter increase was found 11.7 cm. During 1981, the average diameter of the largest trees was 17.8 cm (SD = 4.717). Trees also became 5.1 m taller (Table 1).
Tree Size Changes Within 37 Years Period
Trees age of the nearby plot in 2012 (some trees of this plot where cut down) where between 62–74 years old, and the respective ones of the study plot where 61 years old.
The Soil Conditions: Year 2018
The results of the soil condition in the experimental plot are presented in Tables 2 and 3. The findings support a sufficient amount of forest floor and soil nutrients. Statistically significant differences were observed between O1 and O2 layers (confidence interval 99%), (p-value = 0.000 < 0.01). A balance between the quantity of organic matter of the forest floor (21.03 tn/ha) and the topsoil, 0–10 cm (39.03 tn/ha) was observed. Most of the nutrient quantities (Ν, Μg, K, Ca, Na, and Mn), are much higher in the topsoil than in the forest floor, except for Fe, Cu, Ζn, and P. These elements observed on the forest floor (30.77 kg/ha) were higher than the respective value in the topsoil (9.5 kg/ha). Furthermore, Fe in the forest floor was found 80.45 kg/ha and in the topsoil 56.75 kg/ha. Cu in the forest floor was found 4.82 kg/ha and in the topsoil 0.57 kg/ha. Zn in the forest floor was found 2.5 kg/ha, which is slightly higher than the 2.4 kg/ha in the topsoil. Regarding the statistically significant differences (95% confidence interval) of nutrient amounts and soil properties between 0–5 cm and 5–10 cm of topsoil, the results for pH, N, Ca, Mg, and organic matter are as follows (Table 2): pH found higher (5.34) for 0–5 cm and 5.16 for 5–10 cm. The same higher values for 0–5 cm found for organic matter (5.55% for 0–5 cm and 3.07% for 5–10 cm), Ν (2,076 kg/ha for 0–5 cm and 1,933 kg/ha for 5–10 cm), Ca (541 kg/ha for 0–5 cm and 355.5 kg/ha for 5–10 cm), and Mg (129 kg/ha for 0–5 cm and 118.3 kg/ha for 5–10 cm). All values were found higher in the upper part of the topsoil (0–5 cm). The percentage of mycorrhizal colonization was found to be 24.1 percent (sd = 6.5).
Nutrient Changes Within 37 Years (1981–2018) in the Forest Floor of the Experimental Plot
All mean differences are statistically significant at p < 0.05.
Soil Changes within a Period of 37 Years
Forest Floor Changes
The results of the forest floor changes in the experimental plot within a period of 37 years are shown in Table 2 and Figure 3. Data analyses revealed significant differences in the experimental plot comparing the years 1981–2018. Ο1 layer increased from 3.490 kg/ha (1981), 4.970 kg/ha (2001), 5.372 kg/ha (2003) to 7.898 kg/ha (2018) (more than twice). This was also the case for O2 layer, from 11.925 kg/ha (1981), 14.725 kg/ha (2001), 15.316 kg/ha (2003) to 25.790 kg/ha (2018). The highest increase for both layers of the forest floor was observed after 2001 (increased by 13.994 kg/ha). In 2001, the weight of the forest floor was 4.279 kg/ha higher than the value of 1981. Since 2001, the forest floor overtripled its weight, from 15.415 kg/ha (1981) reached 33.688 kg/ha (2018), and thus gained 18.273 kg/ha. In 2001, the value of O1+O2 was 19.694 kg/ha and became 33.688 kg/ha (2018). The O2/O1 ratio was 2.96 for the year 2001 and increased to 3.27 for the year 2018. The organic matter of the forest floor increased by 7.93 tn/ha (from 13.1 to 21.03 tn/ha). N increased by 238.6 tn/ha (from 236.6 kg/ha to 475.2 tn/ha). P increased by 18 kg/ha (from 12.8 kg/ha to 30.8 kg/ha). Mg increased by 61.1 tn/ha (from 40.4 tn/ha to 101.5 tn/ha). Κ increased by 38.8 tn/ha (from 27.5 tn/ha to 66.3 tn/ha). Ca increased by 55.4 tn/ha (from 280 tn/ha to 335.4 tn/ha). On the contrary, Μn decreased by 34.4 tn/ha (from 38 tn/ha to 3.63 tn/ha).
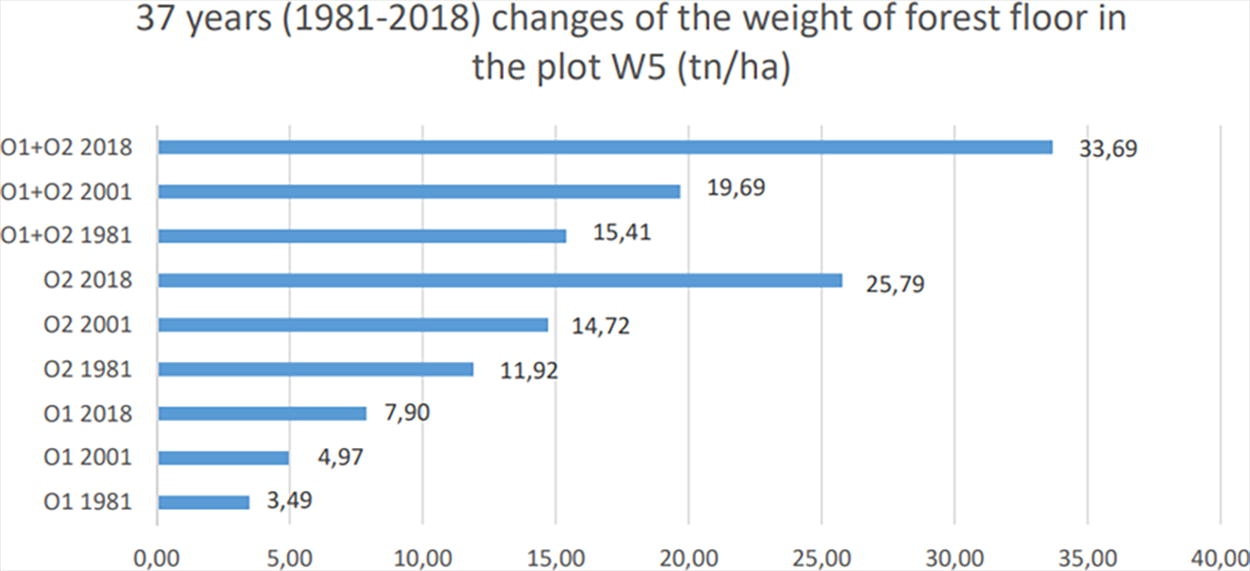
The weight changes of the forest floor within the study period
Topsoil (0–10 cm) Changes Within a Period of 37 Years
The results of the soil sampling analysis (0–10 cm) regarding the pH values as well as the organic C (%) and organic N (%) concentrations are shown in Table 3. pH increased from 4.51 (1981) to 5.25 (2018) thus, as expected, the acidity of the soil gradually decreased over the years. C (%) increased by 0.2 (from 2.3 to 2.5%). N (%) slightly increased from 0.43 to 0.48. C/N ratio remained stable between the years 1981 (5.35) and 2018 (5.17). Organic matter increased by 3.18 tn/ha (from 35.85 tn/ha in the year 1981 to 39.03 tn/ha in the year 2018). The percentage of organic matter increased from 3.96 percent (1981), to 4.31 percent (2018). Ιn the year 2018, its value reduced, which is an indication of balance between production of organic matter and decomposition, showing an indication of a mature ecosystem. All nutrient amounts increased during the study period. Ν (kg/ha) overtripled its value from 1251.7 kg/ha to 4,009 kg/ha. P increased from 8 to 9.55 kg/ha. Mg increased from 140 kg/ha to 247.46 kg/ha. K increased from 73.83 kg/ha to 115.3 kg/ha. Ca increased from 782.48 kg/ha to 896.56 kg/ha. Na increased from 12.90 kg/ha to 54.15 kg/ha. Fe, Mn, Cu, and Zn were also measured in the laboratory. Fe was found to be 56.75 kg/ha, Μn 44.69 kg/ha, Cu 0.57 kg/ha, and Ζn 2.4 kg/ha. The respective measurements for the year 1981 are missing. The higher values of all nutrients (except for Μn) during the study period, were attributed to the increase of the organic matter on the top soil as well as in the soil. The nutrient amounts of topsoil (0–10 cm) (soil weight 905 tn), [(0–5 cm), soil weight 395 tn and (5–10 cm) soil weight 510 tn] are presented in Table 3.
Topsoil (0–10 cm) Properties Changes Within 37 Years Period (1981–2018)
all mean differences are statistical significant at p < 0.05 or p < 0.1 except those with b symbol.
For the depth of 0–5 cm, C (%) was found to be 3.22 percent. Ν (%) for the experimental plots was found to be 0.54 percent (2076 kg/ha). Organic N amount was reduced with the increase of the soil depth (0–10 cm). Statistically, significant differences between the years 2001 and 2018 with confidence interval 95% found for pH (0–5 cm), (p-value 0.006 < 0.05), pH (5–10 cm), (p-value 0.028 < 0.05). Similarly, with confidence interval 90%, significant differences were found for C (0–5 cm) (p-value 0.044 > 0.1, organic matter (%) (0–5 cm), (p-value 0.044 > 0.1), organic matter (t/ha) (0–5 cm) (p-value = 0.00 < 0.1), C/N (p-value = 0.026 < 0.1).
In summary, data analyses revealed differences in the experimental plot comparing the years 1981–2018. Both O1 and O2 forest floor layers had much higher weight in 2018. Since 2001, the forest floor overtripled its weight, from 15.415 kg/ha (1981) to 33.688 kg/ha (2018), and thus gained an extra weight of 18.273 kg/ha. The acidity of the soil gradually decreased over the years. The top soils’ C (%), N (%) increased but C/N ratio remained the same. The organic matter increased by 3.18 tn/ha. All nutrient amounts in the topsoil and the forest floor increased except for Mn (in the forest floor).
Discussion
In the Mediterranean countries as well as elsewhere, coppices were abandoned after World War II, and they became high natural forests. This conversion improved their environmental, social, and productive functions (Chatziphilippidis & Spyroglou, 2006). The forest ecosystems of the study area are under sustainable management and have the chance and conditions to grow older and become mature, self-dependent, and capable of multi-purpose services. Tree diameter increased by 1.3, 4.05, and 5.4 cm within 20 (1981–2001), 17 (2001–2018), and 37 years, respectively. A possible explanation for the low average radius increment (1.3 cm/20 years) could be the high tree density and many years of drought during the period (1981–2001). Tree height increase was 1.63 and 3.48 m within 20 and 17 years, respectively. The more the trees mature the better they grow (Aponte et al., 2010). The importance of mature forests gradually gains interest and awareness. Even in terms of tree volume increase, old-growth forests of most species, exhibit considerable growth rates with increasing tree size (Stephenson et al., 2014). From this perspective, in terms of contribution to human health and various other ecosystem services, old forests are precious likely beyond present human understanding (IUFRO, 2023).
Regarding the 2018 Soil Conditions
Organic matter in topsoil is dependent on the reduction of organic residues on the forest floor (O1 and O2 layers). Τhe weight of the O2 layer was found to be three times higher compared to the O1 layer, which is an indication of relatively good decomposition conditions of organic matter. O2 is the most humified part of the litter and its presence is very important because the functional response of the forest changes when it is present compared to when it is not there. The presence and thickness of the O2 layer are linked to the response to erosion and fire (Santín et al., 2016). Zanella et al. (2020) identify the soil (especially the upper part of it) as an indispensable source for the renewal of a destroyed forest and suggest silvicultural and soil management actions that will promote soil conservation and the faster recovery of natural dynamic stability and resilience.
The rate of organic matter accumulation and the amount and weight of the forest floor were found high (33.688 kg/ha). There is a noted balance between the quantity of organic matter of the forest floor and the topsoil. Most of the nutrient concentrations (Ν, Μg, K, Ca, Na, Mn) are much higher in the topsoil than in the forest floor, except for P, Fe, Cu, and Ζn. All soil properties as well as the C/N ratio were found with higher values in the upper part of the topsoil. The C/N ratio is recognized as a good indication of the decomposition rate of organic matter and available N (the lower the better) (Ross et al., 2011; Yamakura and Sahunalu, 1990). Further research (Aponte et al., 2010; Dietzel et al., 2017; Marty et al., 2017) found the same for C, N and P. Of note, mineralization of organic C and nitrification are indicators of forest ecosystem health and evolution (Morris & Boerner, 1998).
Regarding the Soil Changes within a Period of 37 Years (1981–2018)
Alifragis (1984) found medium values of organic matter, Ν, P, low base saturation, a forest floor dry weight (O1 + O2) of 13.368 kg/ha as well as a net input of organic matter to the forest floor of 917 kg/ha for a period of 2 years (458.5 kg/ha/year). Benham, Vanguelova & Pitman (2012) within a 70-year-old oak plot, reported an average accumulation rate of 340 kg/ha/year, The ratio between dry/net weight found was nearly half, so the dry input of organic matter/year was 230 kg/ha/year, which represents a surplus of 4.600 kg/ha and 8.510 kg/ha for 2001 and 2018, respectively. If the rate of organic matter accumulation was steady, the dry weight of the forest floor must have been 17.968 kg/ha in 2001 and 21.878, in 2018, which is 2.720 kg/ha and 11.810 kg/ha less than what was found in 2001 and 2018, respectively. In 1981, the trees were 34 years old. The recorded mean value for the experimental plot was 20.688 kg/ha (2001) and 33.688 kg/ha (2018). The measures provide that since 1981, the forest floor overtripled its weight. The dry input of organic matter/year increased from 230 kg/ha/year to 366 kg/ha/year in 2001 and reached 549 kg/ha/year in 2018. The lower organic material of (1981–1982) can be attributed to the lower input of foliar litter on the ground due to the young tree age (Alifragis, 1992; Klemmedson, 1987) as well as the increased decomposition rate of the forest floor (Alifragis 1992). Desie et al. (2020), highlight that rich litter admixture improves the base saturation of oak stands on acidified soils.
The acidity of the soil gradually decreased with the years. The C (%) and N (%) increased in topsoils, but C/N ratio remained the same. Pitman (2012) in the United Kingdom, also found topsoils’ C and N increase under oak forest with increasing canopy age.
The organic matter increased by 3.18 tn/ha. All nutrient concentrations in the topsoil and the forest floor increased except for Mn (in the forest floor). Only Μn was reduced by 34.37 tn/ha, from 38 tn/ha (1981) to 3.63 tn/ha (2018). The decrease in Mn concentration might be due to the reduction of air pollution because of the introduction of filters in all industries of the country. Further, during the rain incidents of August 1981, high concentrations of Κ, Mg, and Mn were reported. This might be due to the huge forest fires of the Kassandra peninsula (summer of 1981) and the ash spread all over the area through the atmosphere. In order to compare the values, measurements from July 1981 were considered. For this reason, sampling took place in July 2018. Every month, nutrient concentrations change because there is constant movement and flow in the ecosystems. In 1981, high concentration of P was also recorded (higher than the usual literature value). This was attributed to pollution from a nearby fertilizer industry. Additionally, high Mg concentrations for the period were attributed to the Whitestone mines of Gerakini, 15 km from the study area.
In the Mediterranean oak forests, leaf-fall quality, particularly nutrient content, and decomposition into soil, distinctly modify soil fertility conditions, such as pH, N, P, and Ca concentrations, through different nutrient return pathways, and this affects species distribution and generates a positive feedback effect (Aponte et al., 2010; Andreetta et al., 2018). Chadwick et al. (1998) observed that the leaf-fall decay rate was influenced by the nutrient content of the layer of litter on which leaf-fall was incubated.
Coexistence and Symbiosis
The response of the soil system to catastrophes as well as ongoing climate change depends on the degree and type of self-organization reached by the soil at the time of the catastrophe; The rule is that the more organized the soil, the more resilient it is (Zanella et al., 2020). Therefore, coexistence and symbiosis are fundamentally important.
The importance of the role of plants-microbial communities — soil properties feedbacks as drivers of plant community composition and species coexistence is being increasingly recognized (Aponte et al., 2010). Related, an indication of tree maturity is the ability to coexist. Many studies have shown that trees are ecosystem engineers able to generate species-specific effects on soil properties, microbial biomass, and mycorhizal fungal communities that could potentially lead to this feedback effect while increasing their own fitness (Aponte et al., 2010). In line with our findings, Aponte et al., (2010) found higher levels of accumulated organic matter and decomposition and thus, higher input and retention of soil nutrients and moisture, further reinforcing the ability of oaks to positively influence leaf-fall decay and soil conditions. Plants form below-ground associations with mycorrhizal fungi in one of the most common symbioses on Earth.
Adaptive behavior of plants, including rapid changes in physiology, gene regulation, and defense response, can be altered when linked to neighboring plants by a mycorrhizal network (Gorzelak et al., 2015). Networks can facilitate early plant survival, growth, and defense responses. Mycorrhizal fungal networks occur where mycorrhizal fungal mycelia link the roots of multiple plants, including those of different species, sometimes facilitating interplant transfer of C, nutrients, or water (Simard, 2009a). The hierarchical integration of this phenomenon with other biological networks at broader scales in forest ecosystems, and the consequences observed when it is interrupted, indicate that underground tree talk is a foundational process in the complex adaptive nature of forest ecosystems (Simard et al., 2013). The communication of trees are molecular signals that fly in the air or travel in the soil and this chemical language allows trees to face adversity and dangerous parasites as well as to find resources for feeding themselves and their progenitors. Trees seem to speak to each other as in a big family that occupies a whole forest-ecosystem and to the point of questioning the possibility that species make ecosystems (a set of trees is not a forest); it would seem that only ecosystems can arise, not single species (Wohlleben, 2018). The acceptance of social development in plants highlights that radical changes need to be made in the scientific approach; a mass of data is not science, any more than human observation limited by human sensory perception is not sufficient to evaluate the natural world (Zanella et al., 2020).
Complex systems thinking has inspired both theory and applied strategies for improving ecosystem resilience and adaptability, but applications in forest ecology and management are just beginning to emerge. Some researchers are already studying the forest as an integrated social-ecological system and provide concrete examples of how to manage forests as complex adaptive systems (Filotas et al., 2014). As Zanella et al. (2020) note: “We need to deconstruct our human-sided view of Nature because the correct way stays in the future, and it does not exist a priori. The dominant scientific culture is aimed at planning the future on the basis of data acquired with experiments and observations but in biology and forestry (operating in a constantly changing environment), current knowledge does not allow us to be certain that the change in some conditions does not affect the results.”
Simard et al. (2013), suggested that mycorrhizal networks are fundamental agents of complex adaptive systems (ecosystems) because they provide avenues for feedback, cross-scale interactions, and interconnectivity that lead to self-organization and emergent properties in ecosystems. In the forests, properties at higher levels emerge from localized networks of many entities interacting at lower levels, allowing the development of multiple ecological pathways and processes (Ibarra et al., 2020).
Jones (2010) found that significant amounts of C were transferred between plant species, showing for the first time in a balanced pulse-labeling experiment that approximately three times as much C was transferred through mycorrhizal pathways than soil pathways. This C transfer could affect competitive interactions between establishing seedlings, favoring some included in this network while disadvantaging others, thus fundamentally altering our understanding of how interspecific interactions alter community structure. The existence of a root/mycorrhizal exudation—hyphal uptake pathway was also supported by Pickles et al. (2017).
Hub trees for mycorrhizal networks are foundational because they even out resource availability and create favorable local conditions for tree establishment, which is fundamental to the structuring of the whole forest community. Mature trees provide favorable conditions for seedling establishment (Heydari et al., 2022). Therefore, conserving hub trees and mycorrhizal networks during forest management is important for the conservation, regeneration, and restoration of forests.
Organisms, such as fungi, trees, and woodpeckers, and processes, such as disparate as disturbance, dispersal, facilitation/competition, and nutrient cycling, are related through cross-scale metanetworks (comprised of many individual networks) in the development of healthy functioning forests (Simard et al., 2013).
Poor mycorrhizal networks, drought, and low-nutrient availability can significantly reduce tree growth and the important factors for tree resilience to stressors such as drought.
Tree Resilience to Stressors Such as Drought
During the first part of the study period (1981–2001), the trees were younger, the tree density higher, and the nutrient concentrations lower. The summer dry season stressed the trees and as observable by dry branches on the top of the trees as well as off-season dry and fallen leaves. After 2001, as the ecosystem grew older and despite the continuity of the dry summers, the trees looked healthier and their growth rate improved (Table 1a). This improvement is attributed to the role of mycorrhizal networks, tree size-age, and nutrient concentrations.
The Role of Fungi Colonization
All these studies have important implications when considering the role of mycorrhizal networks in influencing forest health and mitigating stress from environmental conditions. Mycorrhizal networks are conduits for the transfer of resources between hosts (Birch et al., 2021) and trees. Greater growth is positively associated with the number of connections to other trees via ectomycorrhizal networks by which a tree was colonized. Simard (2009a) proposes that interacting networks function to stabilize the forest. Beneficial soil microbes, such as mycorrhizal fungi and N-fixing bacteria, can improve plant nutrient acquisition and increase plant resilience to stressors (McMahen et al., 2022). Pither et al., (2018) studied the below-ground biotic interactions—symbioses, which moderated the postglacial range dynamics of trees and addressed the importance of receptivity of host genera toward ectomycorrhizal fungi for increasing the ecosystems’ resilience.
To help address the challenges of climate change, future soil ecology research and management would benefit from a complex systems approach, where network and dynamics systems theory are used to predict plant community and ecosystem responses to disturbance. Simard (2009b) proposed a conservationist approach for managing forest mycorrhizal networks and hub trees that can bolster ecological resilience. Interdisciplinary research that integrates the dynamics of multiple, overlapping networks will help develop management practices that sustain ecosystems in the changing climate. Mycorrhizal fungi are known to provide their hosts with increased drought resistance in many circumstances, (Pickles & Simard, 2017). The ability of many mycorrhizal fungi to form mycelial networks, in which an individual mycelium connects to the roots of multiple host plants, provides a mechanism for the redistribution of water into upper soil layers experiencing low soil water potential. Soil properties, drought, and the interaction between the two are significant. Kostić et al. (2021) find reliable associations (moderate to strong) among soil properties, pedunculate oak radial increment and its drought sensitivity.
The Role of Tree Size-Age
The mycorrhizal fungal community’s structure was strongly influenced by stand age than specific soil nutrients (Twieg et al., 2009). Colangelo et al. (2017) found that the tallest trees of the anisohydric Italian oak better survived drought contrary to what was predicted by the theory. Τall trees are more able than small trees to obtain soil water with their deeper root system under intense and prolonged drought stress. Dead trees were characterized by a lower height and radial-growth trend than living trees in both study sites.
A Q. cerris forest with trees 85–180 years old, displayed stand dynamics consistent with expected patterns of decreasing tree density and increasing basal area (Wood et al., 2018). Root pathogens only become pathogenic when trees were stressed due to drought and growth conditions.
In summary, the growth (stand age) of the oak ecosystem significantly increases the input of organic material. The amount of forest floor is higher in mature forests than in young or middle-aged forests (Yanai et al., 2000). Soils of high forests and also from small trees to large trees were more fertile (total N, available P, available N, and organic matter, with a lower bulk density and a higher soil porosity) (Heydari et al., 2021).
In addition, forest covers and floors are two important components in forest ecosystems, affecting water quality/quantity in streams of watersheds, and forest floors had higher impact on rain water chemical content than canopy covers (Eisalou et al., 2013). Forest canopy covers significantly affected pH, total N, total P, Na, and K contents in the throughfall, while forest floors influenced almost all chemical properties in the forest floor solution. Drought, especially in the early stages of decomposition, can limit microbial activity; later, litter has an increased ability to retain water . Canopy significantly decreased p values of rainwater, while correspondingly forest floor retains a significant amount of it.
The Role of Nutrients
Given that mycorrhizal networks mitigate tree stress to environmental conditions such as drought, their welfare also depends on soil nutrients. The improvement of soil properties influences the prosperity of microbial communities and vice versa. The shift in fungal species composition was related to changes in litter and topsoil properties. Ca concentration, strongly related to the soil pH, emerged as the best predictor of the ectomycorrhizal community composition. Soil organic matter which is responsible for much of the cation adsorption on the surface layer, for example, retains NH4+ and prevents its loss by the downward movement of water, is the major reservoir of soil N. Other soil chemical properties may modify the N dynamics, acting as nutrients (K, P, Ca, Mg) for microorganism’s growth (Brady & Weil, 2008). Twieg et al. (2009) found significant relationships between mycorrhizal fungal diversity and both available and organic P. Under low nutrient availability tree dependence on ectomycorrhizal symbiosis for nutrient uptake increases, and also soil fungi are particularly sensitive to disturbances such as disruption of the forest floor and the changes in C and nutrient dynamics (Rodriguez-Ramos et al., 2021).
Soil nutrients can directly influence growth response of trees to drought. Interactions between water availability and soil nutrients are significant and their effects on tree growth and response to climate are considerable. A study of soil properties, throughfall and stemflow interactions in a Q. cerris forest with similar tree height and age as well as the precipitation to the study area, revealed higher stemflow where trees had a narrower branching angle and smaller canopy volume due to lower concentrations of soil nutrients (Corti et al., 2019). Low organic C/N ratios (rich soils) and high soil water-holding capacities can potentially buffer the negative effects of the increase in drought frequency and intensity on tree growth (Lévesque et al., 2015, Gessler et al., 2016). The physiological mechanisms underlying drought impacts are focused on the water- and carbon-associated mechanisms, whereas the role of nutrients is notably missing from this research agenda. Gessler et al. (2016) investigated what role forest nutrition plays in the survival and recovery of forests during and after drought and found that high nutrient availability may play a crucial role in drought survival. Low nutrient supply favors C starvation under long-term water stress. Nutrients are major limiting factors for plant and forest productivity (Fisher et al., 2012).
Conclusions
This study focuses on the widely distributed but not extensively examined Mediterranean oak ecosystems and aims at increasing the awareness and the knowledge about them importance of their interconnections. It is significant to note that only ecosystems can arise and face climate change, not single species (humans included). Due to the global multiphase crisis, these ecosystems are in danger, and it is crucial to let them mature and grow.
In the present study, which spanned 37 years of changes specific to trees, forest floor, and topsoil, the soil properties were found to be essential in the context of sustainable forestry. As the trees grew older, all soil properties were found to be more nutrient rich with temporal improvement attributable to interconnected links and interactions (feedback effect). The aging of the trees involves an important change in the soil which must be safeguarded; essentially, the more the forest grows in biomass and biodiversity, the more resilient it becomes. The latter can be attributed to the tree and its relationship with other trees and coinhabiting species. Given the symbiosis found in forest ecosystems, oak and fungi symbiosis and the role of nutrients and stand age interactions should be further studied, as this may address natural adaptation to climate change.
Footnotes
Acknowledgment
The authors would like to thank Prof. Dimitrios Alifragis for the comparison data he provided.
Authors’ Contributions
H.P.G.: Conceptualization (Ideas; formulation or evolution of overarching research goals and aims.). Data curation (Management activities to annotate (produce metadata), scrub data and maintain research data (including software code, where it is necessary for interpreting the data itself) for initial use and later reuse. Methodology Development or design of methodology; creation of models.) Project administration Management and coordination responsibility for the research activity planning and execution. Supervision (Oversight and leadership responsibility for the research activity planning and execution, including mentorship external to the core team. Validation (Verification, whether as a part of the activity or separate, of the overall replication/reproducibility of results/experiments and other research outputs. Visualization Preparation, creation and/or presentation of the published work, specifically visualization/data presentation. Writing—original draft (Preparation, creation and/or presentation of the published work, specifically writing the initial draft (including substantive translation. Writing—review and editing Preparation, creation and/or presentation of the published work by those from the original research group, specifically critical review, commentary or revision—including pre- or post-publication stages. C.V.: Formal analysis (Application of statistical, mathematical, computational, or other formal techniques to analyze or synthesize study data. H.P.G., E.A.P., E.P.: Funding acquisition (Acquisition of the financial support for the project leading to this publication). No financial support. Investigation (Conducting a research and investigation process, specifically performing the experiments, or data/evidence collection.). H.P.G., E.A.P., E.P.: Resources (Provision of study materials, reagents, materials, patients, laboratory samples, animals, instrumentation, computing resources, orother analysis tools. H.P.G., C.V.: Software (Programming, software development; designing computer programs; implementation of the computer code and supporting algorithms; testing of existing code components.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.