
Abstract
My concept of cell differentiation involves genetic information from DNA being transcribed into mRNA protein complexes-morphogens (mRNAs + homeodomain proteins), and stored in the ovoplasm as maternal inheritance, or cytoplasmic genetic memory. Feedback mechanism(s) allow these morphogens to selectively unlock new genes, regulating the development of the embryo. The blastomeres and the embryonic pluripotent cells of the inner cell mass of early (5 day) blastocysts are loaded with morphogens which hamper the production of cell lines and are responsible for the formation of embryoid bodies in vitro and teratomas in vivo. There are therefore legitimate concerns as to proposals to use embryonic pluripotent cells for cell therapy and regenerative medicine. An alternative cell therapy would involve the production of tailored morphogens, and hence selective in vitro differentiation of adult de-differentiated cells.
Aprerequisite for a complete understanding of embryonic pluripotent cells and their properties is a comprehensive knowledge of normal fertilization, egg maturation, cleavage, and the processes of cell differentiation and chromatin remodeling during embryonic development, as well as an appreciation of the role of ovoplasmic maternal factors in directing embryonic development after fertilization. During the pachytene and diplotene phases of the first meiotic division [1], homologous chromosomes fuse, forming the “synaptonemal” complex [2 (Fig. 20.15)]. The chromatids of the central element exchange segments, an event known as “crossing over.” The two lateral chromatids, or elements, then develop thousands of side loops [3 (Fig. 15-14)], which display intensive transcription, a process that continues in the next diplotene phase [4 (Fig. 11-9)]. The nascent RNA transcripts of the transcriptionally active chromosomes—“lampbrush chromosomes” [5–7]—associate with specific proteins known as translation repressor proteins [3]. They can prevent the translation of mRNAs. The lampbrush chromosomes are commonly observed and studied in amphibians, birds, mammalian and human oocytes. I contend that all of the genetic information transcribed into mRNAs, bound and protected by special nuclear homeodomain proteins, is stored in the ovoplasm as mRNP particles [8–10], which mediate ovoplasmic maternal inheritance [11–13].
The homeodomain nuclear proteins contain a 184 bp sequence called the homeobox or Hox-box [14]. The Hoxproteins have been conserved for over 500 millions years and are very similar in all animals studied, including man. They orchestrate the body plan, segmentation, and cell differentiation of organisms [15]. There is an astonishing amount of information on the proteins that pattern the axes and segments of an animal, but we do not yet understand how Hox-proteins combine to produce the numerous stereotypical, morphological patterns. Feng et al. [16] recently reported a “novel mechanism whereby transcription is controlled by the cooperative actions of noncoding RNA (ncRNA) with a homeodomain protein. The possibility that a subset of ultraconserved regions may function [in vertebrate development] at both the DNA and RNA level to control key developmental regulators may explain why such sequences exhibit 90% or more conservation even after 450 million years of vertebrate evolution.” The findings of Feng et al. coincide and fit very well with my new concept of cell differentiation, as previously reported [17]. The same homeodomain proteins that bind the nascent mRNA in the lampbrush chromosomes bind the complementary DNA gene sequences in the replicative fork, because the complementary mRNA attaches to them selectively within the corresponding gene. As a result, the gene is now deblocked, and ready to participate in the developmental process.
There is a misconception that the process of transcription is equivalent to differentiation. “There are two distinct levels of regulation in mammalian somatic cells. One level is concerned with genomic commitment (i.e., determination) and the other level with the expression of tissue specific proteins” [3 (p247)]. Hence, it is critical when considering epigenesis not to confuse gene deblockage and the processes of differentiation with gene on/off switches and the processes of gene expression or transcription. Differentiation must await transcription and translation of new proteins, in a highly orchestrated process induced by prior gene deblockage.
During cleavage, the mRNPs are asymmetrically segregated in the blastomeres, which, at least in the amphibian, are proposed to contain different maternal information [18]. “Nucleo-cytoplasmic interactions are believed to be of great importance in early animal development, since the response of genetically identical nuclei to different regions of egg cytoplasm provides the most satisfactory explanation for the initial appearance of cell differences” [19,20]. The blastomeres, including the true embryonic pluripotent cells of the blastocyst inner cell mass, undergo proliferating symmetric mitoses, keeping the nuclei of all cells genetically identical. It is necessary to distinguish between asymmetric self-renewal of stem cells giving rise to two different daughter cells, and self-replication, or simple duplication, resulting in two equal daughter cells. A new hypothesis about the molecular mechanisms responsible for stem cell’s self-renewal is presented in Figure 1 [21].
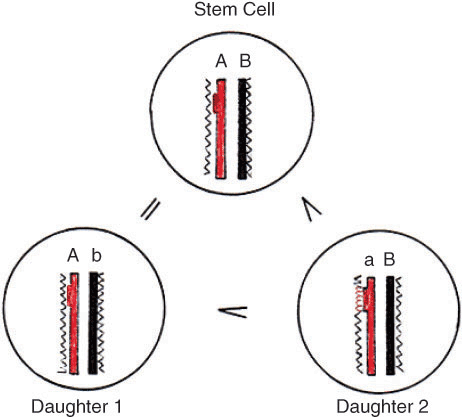
A new hypothesis for stem cell self-renewal and differentiation. Sense DNA strand (A) with one structural gene and antisense strand (B). Both DNA strands are blocked by histones (^^^^). In daughter cell #1 on the template of the parental strand A, a complementary strand b is synthesized, but it is an antisense strand and does not carry the gene. Daughter #1 is a replica of the mother cell and depicts the self-renewal of the stem cells. In daughter cell #2, the structural gene is in the newly synthesized sense strand a, accessible for morphogens. The complementary mRNA serving as chaperone will read and add the Hox-protein (^^^^) to the cDNA. Thus the gene will be unlocked and under activation can start transcription. The two daughter cells are not equal, #2 undergoing differentiation.
After gastrulation, the three primary germ layers—ectoderm, mesoderm, and endoderm—are formed. What is the difference between committed stem cells and the pluripotent embryonic cells, which I have argued elsewhere [22] are incorrectly referred to as embryonic stem cells (ESC)? The committed stem cells are partially differentiated, tissue specific, and persist to the end of an individual’s life. Under normal circumstances, they divide asymmetrically: one daughter cell is a replica of the mother cell (self-renewal) and maintains the pool of stem cells; the second undergoes further differentiation. The ESC are undifferentiated, not yet tissue specific, and do not persist. They divide symmetrically by proliferating mitosis, keeping the nuclei in all cells genetically identical.
All structural genes in the zygote are blocked by histones, except the housekeeping genome, whose genes are packed with nonhistone homeodomain proteins. The histones bind the nascent DNA strands forming nucleosomes in the following order: H2A, H2B, H4, H3, H3, H4, H2B, and H2A. Thus, the nucleosome core particle is an octamere and clearly has a bipartite structure [22 (p679)]. During replication, the nucleosomes split into two tetrameres, each attached to the parental DNA strand (Fig 2). In the past the semi-conservative distribution of the nucleosomes between the two DNA strands was refuted, due to experiments with radioactive-labeled histones in favor of the conservative attachment of the entire octamere (nucleosome) to one of the parental DNA strands. Figure 2 shows why the authors reached this erroneous conclusion; they did not consider the binding of nonhistone Hox-proteins by the newly synthesized gene caring sense strand. The array of histones in the nucleosomes and their bonds to each other and to the DNA support the semi-conservative distribution of nucleosumes in the replicative fork. On the other hand, the presence of histones in the transcriptionally active euchromatin then becomes more understandable, since the nonhistone proteins bind the newly synthesized DNA strand only, while the old one remains with the histones of the nucleosomal tetramere. The attachment of nonhistone proteins makes the formation of a new nucleosome impossible and clarifies why the active euchromatin is deprived of nucleosomes [23–25].
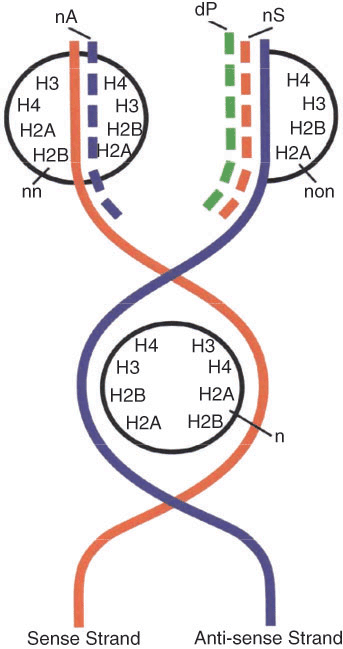
Molecular mechanism of cell differentiation and re-modeling of heterochromatin (inactive) into euchromatin (active). During replication, the nucleosomal octamere (n) splits into two tetrameres, each attached to the old (parental) DNA strand. The gene in the old sense strand remains inactive. The newly synthesized antisense DNA (nA) will adopt histones and form a new nucleosome (nn) with the old tetramere, a process accompanied with rotation, responsible for the double winding of the DNA helix around the nucleosome. If the applied histones are radioactive, this double helix will be labeled. On the template of the old antisense strand a new sense strand (nS) caring a gene sequence will appear. In the presence of a morphogen (mRNP particle), the complementary mRNA will attach the nonhistone homeodomain protein (dP) to this gene, which will prevent the formation of a new nucleosome (non). The deblocked gene now belongs to the active euchromatin, which is missing nucleosomes. This double helix is not going to be radioactively labeled and will leave the false impression for conservative distribution of the nucleosomes.
DNA packed with histones (H1 in the spacers and the core histones in the nucleosomes) is transcriptionally inactive and represents the heterochromatin. An essential question widely treated in the related literature is how heterochromatin turns into euchromatin. My concept about chromatin remodeling is depicted in Figure 2. It necessarily accompanies the molecular processes of cell differentiation, which for the first time was presented in my paper [17]. The mRNP particles, synthesized by the lampbrush chromosomes of the meiotic oocyte, are stored in the ovoplasm, where they represent the morphogens responsible for the selective unlocking of new genes during embryogenesis. The mRNA brings the homeobox protein to the nascent complementary gene, and if DNA and protein are complementary and therefore form a hydrogen bond, a stable complex forms, which keeps the gene unlocked [23 (p152)]. Thus no nucleosomes can be formed and heterochromatin is turned into active euchromatin.
The cytoplasm of the so-called embryonic stem cells of the inner cell mass is loaded with morphogens. I propose that when these cells are removed from their natural environment, the morphogens cause chaotic differentiation, giving rise to embryoid bodies in vitro and to teratomas, when transplanted in vivo. Therefore, I contend these cells are not suitable for creation of cultured cell lines or for treatment of different diseases, because they do not transform themselves spontaneously into cells of the organ where they are transplanted.
Logically, then, the further destruction of human embryos for collecting ESCs has not yet been justified, as these cells cannot be used for regenerative medicine. By the same token, efforts to produce ES cells by introducing an adult nucleus into the enucleated egg are irrelevant as well, because the transplanted nucleus, independent of its origin, is subject to the same controlling elements, that is, the morphogens in the surrounding ovoplasm.
I propose a new approach to regenerative medicine: specific morphogens, obtained by PCR, allow the mRNAs to bind homeodomain proteins and to form the mRNP complexes, which could be added to the culture of adult dedifferentiated cells (virgin cells). The morphogen would then selectively unlock the complementary gene, as happens in the developing embryo (Fig 2). This kind of gene engineering will produce highly differentiated cells in vitro for the special needs of the patient.
Another method to obtain specific morphogens would be possible due to the following fact: if a nucleus from a differentiated cell is injected into the cytoplasm of another differentiated cell, the transplanted nucleus acquires the characteristics of the host cell [3 (p900)]. Each differentiated cell contains small amounts of mRNP particles, the messengers being transcribed on the template of the active genes specific for the cell type. These mRNPs (morphogens) are the cytoplasmic genetic memory, which, during replication, maintain the specific genes permanently unlocked and the cells permanently differentiated. The same deblocking factors in the host cell are responsible for the transformation of the donor nucleus. Therefore, an extract of given differentiated cells would contain the necessary factors (morphogens), able to redifferentiate in vitro dedifferentiated cells from the patient (see review [26]).
In conclusion, my new concept for cell differentiation consolidates all previous experimental data about homeodomain proteins, mRNP complexes, morphogens, chromatin remodeling, and cell transdifferentiation into a workable model. It opens new avenues, placing powerful tools into the hands of scientists who could produce specially differentiated autologous cells for the needs of cell therapy and regenerative medicine.