
Abstract
Spleen stromal cells are critical determinants of dendritic cell (DC) development in spleen. The spleen stromal line, namely STX3, supports DC differentiation in vitro from overlaid bone marrow cells while the lymph node stromal line, namely 2RL22, does not. Here we have characterised the hematopoietic support capacity of each stroma, and analyzed lineage origin of the stromal cell lines by gene profiling using microarrays. Stromal co-culture experiments were performed using bone marrow cells as a source of hematopoietic progenitors. A characteristic immature myeloid-like CD11c+CD11b+CD86+MHC-II−/loB220−CD8α− DC is produced after 14 days in STX3 cocultures, while 2RL22 cocultures produce only monocyte/macrophage-like cells. No other hematopoietic cell type is produced. The STX3 and 2RL22 stroma were compared by transcriptome analysis utilizing Affymetrix Murine U74Av2 genechips to identify gene expression related to differential hematopoietic support function. Data mining was used to determine cell surface marker expression reflecting endothelial cells and fibroblasts, as well as adhesion molecules contributing to the microenvironment. STX3 shows gene expression reflective of early endothelial cells, while 2RL22 expresses markers specific to fibroblasts. The expression of genes like Flt1, CD34, Mcam, and Eng distinguishes STX3 as an early immature endothelial cell lacking markers of angioblasts or hemangioblasts like Tal1/SCL, Tie1, Tie2, Kdr or Prom1/AC133. The absence of expression of genes like Vwf and Cd31 distinguishes STX3 from fully differentiated vascular endothelial cells. In contrast, the 2RL22 lymph node stroma specifically expresses genes related to fibroblastic-like cells like osteoblasts with expression of Vdr (Vitamin D receptor), and epithelial cells with expression of Krt13 (keratins). Gene expression data identifies STX3 as splenic endothelial cells, independently able to support the outgrowth of immature, myeloid DC-like cells from progenitors present in bone marrow, while 2RL22 lymph node fibroblastic cells provide support for development of monocytes/macrophages.
Introduction
The role of extrinsic factors in dendritic cell (DC) development from precursors has been mapped out. For example, GM-CSF and TNF-α can induce differentiation of DC from bone marrow precursors [1] or from monocytes under inflamma-tory conditions [2], and Flt3L appears to be essential for the differentiation of DC precursors into conventional myeloid-like DC as well as plasmacytoid (p) DC [3]. However, the deterministic factor in hematopoiesis of specific cell lineages now appears to depend on a specific stromal cell niche, composed of cells producing extracellular matrix proteins and secreted growth factors. Recently, splenic stromal cells were also identified as critical determinants of DC differentiation and maturation [4–7]. The lineage origin of stromal cells which support DC hematopoiesis is not clearly defined and methods to purify and classify these cell types vary across different reports. One difficulty is the lack of information on the development of endothelial type cells and the common gene expression between developing endothelial cells and fibroblastic cells.
In this lab, splenic stromal cells supportive of DC development have been characterised as endothelial-like on the basis of cuboidal morphology, capacity to form tube-like structures in Matrigel and marker expression [4]. In another study, a splenic endothelial cell subset isolated on the basis of expression of mature endothelial cell markers as a CD11b−VCAM+CD31+ cell population was found to support the maturation of immature DC into suppressive or regulatory DC [5]. Another report has defined the DC supportive stromal population in spleen as fibroblasts expressing the ER-TR7 antigen, together with CD68+ macrophages [6]. Previous reports from this laboratory have described the production of immature myeloid-like DC in long term stroma-dependent cultures of spleen [8–10]. A splenic stromal line, namely STX3, was derived from a productive long term culture (LTC) of spleen which had ceased production of DC after loss of hematopoietic pro-genitors with passage. It has been shown to support in vitro production of DC-like cells from overlaid bone marrow. STX3 cells are morphologically cuboidal and endothelial-like [11]. In con-trast to STX3, the morphologically distinct 2RL22 lymph node stroma which is a non-supporter of DC hematopoiesis is composed mainly of elongated fibroblastic-like cells. To further delineate the stromal environment conducive to DC development in vitro, we performed transcriptome analy-sis using Affymetrix Murine U74Av2 Genechips to identify genes related to the DC supportive function of STX3 [11]. Previously we published data on gene expression specific to a supportive niche. In par-ticular differential gene expression analysis was used to identify gene expression in STX3 but not in 2RL22 stroma lines. A subset of 154 genes reflecting categories important in determination of microen-vironments for differentiation was then selected, and from this a smaller subset of genes of interest was selected for further analysis as specific markers of STX3. In this report, the Affymetrix dataset has been further investigated with a view to characterising the type of cell(s) which constitutes a niche environ-ment for DC development. Analysis included expression of cell surface markers reflective of fibroblas-tic versus endothelial cells, as well as endothelial cells in different states of maturation.
Materials and Methods
Cell lines and animals
Animals were bred under specific pathogen-free conditions in the John Curtin School of Medical Re-search (Canberra, ACT, Australia) and used when 6–8 weeks of age. They were bred and handled according to protocol no. F.BMB.09.04 approved by the Animal Experimentation Ethics Committee of the Australian National University (Canberra, ACT, Australia). Derivation of the splenic stroma STX3 and the lymph node stroma 2RL22 have been previously described [8–11]. They were derived from LTC established from spleen and lymph nodes of B10A.(2R) mice. STX3 derived from a produc-tive spleen LTC which had lost hematopoietic progenitors over time and ceased production of DC. 2RL22 was derived from a lymph node LTC which failed to support hematopoiesis. Both lines have been passaged for many years and represent homogeneous cell populations, STX3 reflective of endo-thelial cells and 2RL22, fibroblast-like cells. For experiments, stromal cells with low number of pas-sages (<7) were used in coculture assays. Stromal cells were cultured at 37°C 5%CO2 in air in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal calf serum, 5 × 10−4 M 2-mercaptoethanol, 10 mM Hepes, 100 U/ml penicillin, 100 µg/ml streptomycin, 4 mg/l glucose, 6 mg/l folic acid, 36 mg/l L-asparagine, 116 mg/l L-asparagine HCl. Stromal cell lines were maintained by scraping attached cells for passage into a new flask.
Coculture assays
For coculture assays, stroma was grown to 80% confluency prior to addition of dissociated, red blood cell lysed suspensions of bone marrow cells. Overlaid cells were isolated from C57BL/6J mice and plated above stroma at a concentration of 5 × 104 cells/ml. This plating concentra-tion was selected as that giving the highest yield of viable cells over time after 14 days. Cocultures were maintained by partial medium exchange when needed, with replacement of non-adherent cells after cen-trifugation. Viable cell recoveries after 14 days of coculture with overlaid bone marrow was 50–75% of input cell number for STX3. Cocultures could be maintained for up to 35 days but were routinely analysed for cell production by collection of non-adherent cells at 14 days for productive bone marrow cocultures. Cell production in cocultures was monitored by phase contrast microscopy and photography using an inverted phase microscope (DMIREZ: Leica, North Ryde, NSW, Australia) equipped with a digital camera (DFC: Leica).
FACS analysis
Cells produced in cocultures were characterised after collection of non-adherent cells released into the supernatant. Cell recovery and phenotype was determined by flow cytometry after staining cells with propidium iodide (PI: 100 ug/ml) to detect live cells, along with antibodies specific for CD11c (N418, hamster IgG, allophycocyanin (APC) conjugate), CD11b (M1/70, rat IgG2b, fluorescein isothiocyanate (FITC) or biotin conjugate), B220 (RA3-6B2, rat IgG2a, phycoerythrin-Cy-5 (PE-Cy-5) or biotin conjugate), CD8α (53-6.7, rat IgG2a, phycoerythrin (PE) con-jugate), MHC-CII (IAb) (25-9-17, mouse IgG2a, biotin conjugate) and CD45RB (C383.16A, rat IgG2a, PE conjugate). Isotype control antibodies with the same fluorescent conjugate were included in all experiments. These included hamster Ig (eBio299Arm, APC conjugate), rat IgG2b (A95-1, biotin conjugate or KLH/G2b-1-2, FITC conjugate), rat IgG2a (R35-95, PE conjugate), mouse IgG2a (eBM2a, biotin conjugate). Fluorescent conjugates used included PE-streptavidin and APC-Cy7-streptavidin. All antibodies and staining reagents were purchased from eBiosciences (San Diego, CA, USA). Multicolour FACS analy-sis was performed on a LSRII flow cytometer equipped with dual lasers for analysis of up to 6 colour fluorescence (Becton Dickinson, Franklin Lakes, NJ, USA). Single colour controls involving bone mar-row or spleen cells staining the same antibody were used to automatically compensate the machine. Data analysis involved gating live (PI−) cells, and multicolour analysis of large cells gated on the basis of Forward scatter (FSC) and Side scatter (SSC).
Microarray analysis of gene expression
Both STX3 and 2RL22 stroma were harvested at 80–90% confluency followed by extraction of total RNA using Trizol (Invitrogen Life Technologies, Mount Waverley, VIC, Australia). Synthesis of cDNA involved T7-(dT)24 primers and SuperScript II according to manufacturer’s instructions (Invitrogen Life Technologies), followed by second strand synthesis with DNA polymerase 1 (Promega, Annandale, NSW, Australia). In vitro transcription and biotin labelling were performed using the BioArray High Yield RNA Transcript Labelling Kit (Affymetrix, Santa Clara, CA, USA). Labelled cRNA was fragmented and hybridised to Test 3 chips (Affymetrix), prior to hybridisation to Murine Genome U74Av2 Genechips (Affymetrix). Hybridisation involved 0.05 µg/µl biotin-labelled cRNA in hybridisation buffer (100 mM MES, 1N [Na+], 20 mM EDTA, 0.01% Tween-20) supplemented with 0.1 mg/ml herring sperm DNA and 0.5 mg/ml acetylated BSA for 16 hours. Washing and staining with streptavidin-phycoerythrin were performed on the flu-idics station according to the manufacturer’s instructions (Affymetrix). Preparation of label and hybridization of genechips was performed by Dr Kaiman Peng (Biomolecular Resources Facility, Aus-tralian National University, Canberra, ACT, Australia).
The Affymetrix dataset was generated by analysis of scanned images of hybridised genechips. Binding of target cRNA to its probe set was quantified by phycoerythrin intensity. Signal value, reflecting abun-dance of mRNA in the sample, and p-value, the probability of detection of mRNA within the sample, were calculated using Affymetrix Microarray Suite 5.0 (MAS5.0) software and managed in Ex-cel (Microsoft).
Literature searching was used to identify genes of interest. Gene searching in Affymetrix datasets in-volved gene names from the NCBI database (
Assessment of gene expression
The first criterion used to assess detection of a given probe set was p-value. In this study, a gene was considered expressed if the p-value was 0.005 and the signal value was >20. This represents a more stringent criterion for detection of gene expression than recommended by MAS5.0 which uses p-value ≤0.04 as the standard level for detection. Previously, retrieval of specific datasets on the basis of p-value was used to identify genes specifically expressed by STX3 and 2RL22 [11]. Some probe sets for a given gene gave discrepant results. In particular, this was the case for genes like Cd44, Itgav/CD51 and Itga3/CD49C. Each case was considered individu-ally to determine gene expression. The target sequences of probe sets were downloaded from the Affy-metrix database (
Reverse transcription-polymerase chain reaction (RT-PCR)
RNA was prepared from several cell types including STX3, 2RL22, the D10.G4.1 T cell line, C57BL/6J spleen cells and human umbilical vein endothelial cells (HUVEC) using TRIzol as described in the manufacturer’s instructions (Invitrogen Life Technologies). Purified RNA was DNase-treated using RQ1 RNase-free DNase (Promega, Annandale, NSW, Australia). RNA was then purified using standard phenol/chloroform procedures followed by ethanol precipitation, and then reverse tran-scribed using SuperScript™ First-Strand Synthesis System for RT-PCR (Invitrogen Life Tech-nologies) according to the manufacturer’s instructions.
One microlitre of cDNA product was used for each PCR reaction (∼100 ng). Template was combined with 1X reaction buffer (67 mM Tris-HCl [pH 8.8], 16.6 mM [NH4]2SO4, 0.45% Triton X-100, 0.2 mg/ml gelatin), 1.5 mM MgCl2, 0.2 mM dNTP mix, 0.5 mM of forward and reverse primer and 1U Taq poly-merase in a final volume of 15 ul. PCR conditions were; initial denaturation of 95oC for 3 min, followed by 30 cycles of 95oC for 30 secs, 60oC for 30 secs, 72oC for 30 secs, and a final extension of 72oC for 2 mins. All primers had an annealing temperature of 60oC with the exception of primers for Vcam1 (58oC). Control reactions comprised spleen, D10.G4.1 or HUVEC cDNA (positive con-trols), or no DNA template (negative control). Primers were designed from sequences in either the NCBI or the Ensembl Mouse database (
Primer sequences used for RT-PCR
Results
Differential hematopoietic support capacity of STX3 and 2RL22 stroma
To compare hematopoietic support capacity of stromal lines, dissociated bone marrow cells were cul-tured above near-confluent cultures of STX3 and 2RL22 stroma to define the best conditions for analy-sis. Cocultures were maintained by half medium change as needed. Cell production was monitored by photomicroscopy and analysis of cell surface marker expression on cells using antibody staining and FACS analysis. Cocultures established above STX3 stroma were characterized by the presence of large numbers of cells with dendrites, medium size round non-adherent cells, and scattered foci of smaller cells attached to stroma (Figure 1A). Cocultures of bone marrow established above 2RL22 showed no production of non-adherent cells, although these cultures did support the outgrowth of small to medium size adherent cells which could be dislodged by gentle pipetting (Figure 1C). Stroma cultured without bone marrow overlay cells showed no cell production above stroma (Figures 1B & 1D).
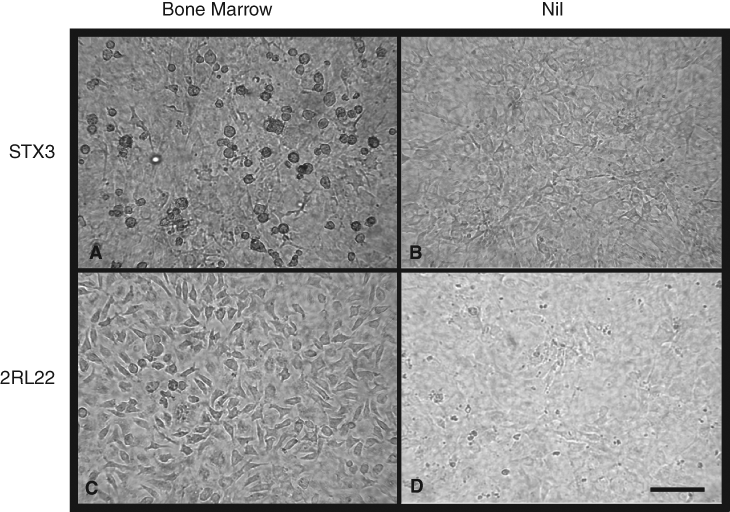
STX3 spleen stroma supports hematopoiesis of bone marrow cells. The STX3 spleen stroma was com-pared with the 2RL22 lymph node stroma for capacity to support hematopoiesis in overlaid bone mar-row cells. Cocultures were maintained for 14 days before photography under phase contrast microscopy (
Over time, non-adherent and loosely adherent cells were collected and counted to determine percent-age of live cell recovery and to assess phenotype of cells produced. After 10 days, cultures begun to produce cells consistently. For the experiment shown in Figure 2, STX3 cocultures gave ∼70% recov-ery, and 2RL22 gave 175% recovery of cells relative to input cell number after 14 days. Clear differ-ences in the cell type produced above the different stroma was evident using multicolour FACS analysis. PI staining was used to gate live cells and live large cells were then gated on the basis of FSC and SSC profile (Figure 2). Two-colour FACS plots shown in Figure 2 indicate expression of c-kit, Sca-1, CD11b, CD11c, CD8α, B220 and MHC-II positive cells amongst CD11c+ DC produced by 14 day cocultures of bone marrow over STX3. Almost 75% of cells had a CD11c+CD11b+ myeloid DC phenotype. These DC were also B220− and CD8α− indicating no production of pDC or CD8α+ cDC. This cell population also lacked progenitors expressing c-kit and Sca-1. Absence of MHC-II expression suggested production of immature myeloid DC, similar to cells produced in stroma-dependent spleen long term cultures [7].
After 14 days, cocultures of bone marrow over 2RL22 produced cells of a different phenotype. The predominant expression of CD11b and no other markers reflected the production of mono-cyte/macrophage lineage cells, but not of DC (Figure 2). These results confirm the unique potential of STX3 stroma to support DC hematopoiesis, but of 2RL22 to support production of mono-cytes/macrophages.
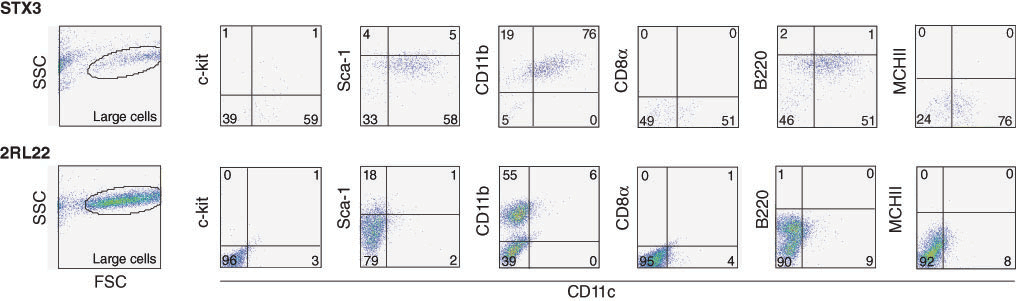
Production of immature myeloid-like DC in cocultures of bone marrow over STX3 stroma. Cocultures were established by overlay of dissociated bone marrow over STX3 splenic stroma and the 2RL22 lymph node stroma. Non-adherent cells were collected at 14 days for analysis of marker expression by multicolour FACS analysis. Staining involved antibodies specific for CD11c, CD11b, B220, CD8α, Sca-1, c-kit, and MHC-II. FACS analysis was used to gate populations of live (PI−), large sized cells based on forward scatter (FSC) and side scatter (SSC) profiles. Isotype control antibodies were used to detect background binding and to set cross hairs such that % cells in relevant quadrants was <0.5%. Percentage of cells staining with specific antibody are shown on each plot. STX3 stroma cultures alone and bone marrow cells cultured alone yielded no cells after 14 days (not shown).
The contribution of bone marrow-derived stoma to cocultures was discounted since previous experi-ments have shown lack of stromal growth when a bone marrow cell suspension was cultured alone over a 14-day period [4]. Recent results from this lab using isolated subsets of bone marrow cells give simi-lar cell production (unpublished data), again arguing against possible contribution of bone marrow-derived stromal contaminants.
Genes specifically expressed by STX3 and 2RL22 stroma
The aim of this study was to identify gene expression unique to these 2 stromal cell types having differ-ent hematopoietic support capacity. Initially, Affymetrix U74Av2 genechips were used to profile gene expression. The procedure used for p-value selection of probe sets specifically expressed by one and not the other stromal cell type is stringent, and gives only those genes which have high certainty of specific expression. The extracted list of genes is therefore not comprehensive in terms of mirroring gene expression. It does, however, serve to delineate with certainty several functions and molecules unique to each of the 2 cell types. When the selected genes were annotated using Gene Ontology or Conserved Domain Database, 154 different genes were identified for STX3 and 70 for 2RL22. These genes were classified into groups according to function. The data files showing genes grouped under functional categories are available as Supplementary files available on the journal website. For STX3, many genes specifically expressed, such as Thy1, Mcam, Cd34, Acvrl1, Fbln1 and Col18a1, indicated an endothelial cell origin [11]. Genes involved in choles-terol metabolism including Mvk and Hmgcr further support this view (Supplementary Figure 1). The gene expression profile for 2RL22 is quite distinct both in terms of gene function and number. Several functional categories present in the STX3 specific dataset were not represented includ-ing chromatin interactions, antigen presentation, intracellular proteases and the ubiquitin pathway (Sup-plementary Figure 2). However, 2RL22 specifically expressed several genes related to the fibroblast lineage, such as vitamin D receptor (Vdr) related to osteoblast as well as keratin 13 (Krt13) and tuftelin 1 (Tuft1) for epithelial cells. These differences equate to the differ-ent cell type present for each stroma and reflect the distinctive hematopoietic support function described for these 2 stroma.
Data mining for expression of endothelial lineage markers
Since genes specifically expressed by STX3 stroma were indicative of an endothelial cell type, STX3 was further investigated for endothelial cell lineage origin by investigating genes known to be ex-pressed by endothelial cells in different states of differentiation. Formation of the endothelium during embryonic development arises by hemangioblast differentiation. During adulthood, renewal of vessel walls occurs throughout life via blood circulating endothelial progenitors. Since heterogeneity exists amongst endothelial cell types, STX3 was investigated for gene expression of cell surface markers re-lated to the hemangioblast, circulating endothelial cell progenitors, and mature endothelial cells (Figure 3).
The expression of genes encoding markers described above was investigated in STX3 and 2RL22. The Affymetrix database showed expression of genes related to hemangioblasts in STX3 but not in 2RL22. These included Cd34, Runx1/AML and Flt1/VGEFR1 (Figure 3A) [12–14]. The specific expression of Cd34 and Flt1 in STX3 over 2RL22 was fur-ther confirmed by RT-PCR analysis (Figure 3B). However, RT-PCR analysis showed that Runx1 was expressed in both STX3 and 2RL22 albeit in apparently much lower levels in 2RL22 than in STX3 (Figure 3B). However, the STX3 splenic stroma lacked the major hemangioblast determinant Tal1/SCL which was expressed weakly by 2RL22 (Figure 3A). Prom1/AC133 and Kdr were not expressed by either cell line (Supplementary Table 1). Furthermore, both cell lines failed to show expression of the adult hemangioblast markers c-kit (Figure 3B), CD45, or the Tie1 and Tie2 angioblast markers (Sup-plementary Table 1). Overall, the data obtained do not support the phenotype of hemangioblast in either STX3 or 2RL22 stromal lines.
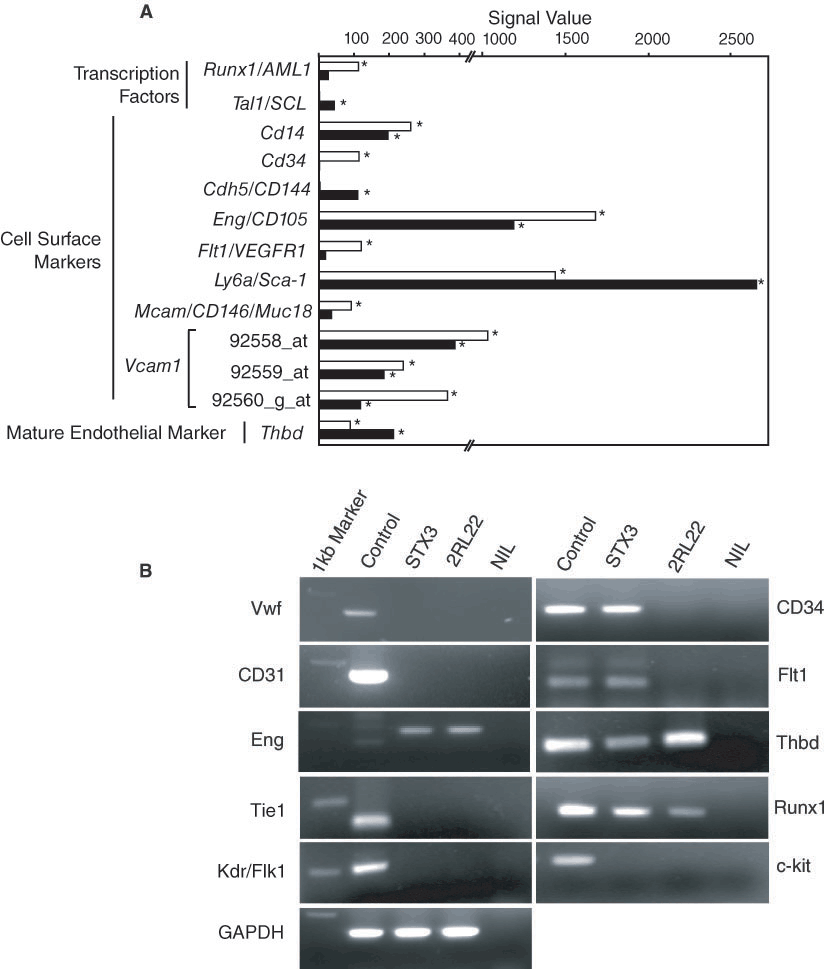
Expression of endothelial markers by stroma. (
Markers for endothelial progenitor cells on various populations include Kdr/Flk1 and CD14 [15,16]. Both endothelial progenitors and monocytes are identified by a phenotype of AC133−Kdr/Flk1+Eng/CD105+CD14+CD34−c-kit−CD45+CD11b+ and can give rise to endothe-lial cells [13]. STX3 expressed transcripts of several of these markers, including Cd14, Cd34 and Eng (Figure 3A). RT-PCR analysis also confirmed the lack of expression of Flk1 and c-kit in both STX3 and 2RL22 (Figure 3B). Absence of expression of Prom1/AC133, Kdr/Flk1, CD45, CD11b and Kit/c-kit by either stromal line does not support the model of an endothelial progenitor cell type (Supplementary Table 1).
STX3 does not reflect a mature endothelial cell. These form blood vessels and are also very heterogene-ous in terms of cell surface marker expression [17]. Markers constitutively expressed include CD34, ICAM2/CD102, Eng/CD105, Pecam1/CD31, Cdh5/CD44/VE-cadherin, CD36, Mcam/CD146/Muc18, Ly6a/Sca1 [17] and Pcdh12 [18]. Of these genes, Cd34, Eng, Ly6a/Sca-1 and Mcam were all expressed at various levels in STX3, with all but Cd34 expressed by 2RL22 (Figure 3A). Endothelial lineage-specific genes not detected in STX3 included Cd36, Pecam1/CD31, Icam2, Cdh5, Pcdh12, Ace, Ace2, F8 and Nos3 (Supplementary Table 1). Absence of gene expression for the common ma-ture endothelial cell marker CD31 in both STX3 and 2RL22 was confirmed by RT-PCR analysis (Figure 3B). Thbd is expressed by STX3 and 2RL22 and this result was confirmed by both Affymetrix gene profiling and RT-PCR analysis (Figure 3). STX3 does not express Vwf which is a common marker of mature endothelial cells and this result was confirmed by RT-PCR (Figure 3B). STX3 there-fore lacks expression of many genes associated with specific functions of mature endothelial cells.
Adhesion molecules, such as ICAM1/CD54, VCAM1, Sele/CD62E and Selp/CD62P are also expressed by mature endothelial cells during inflammation [17]. Both STX3 and 2RL22 lack expression of these genes (Supplementary Table 1) but do express Vcam1, a result confirmed by Affymetrix and RT-PCR (Figure 4). Endothelial cells also express integrins as adhesion molecules. Expression of genes encoding several α and β integrin subunits was therefore investigated in STX3 and 2RL22 [25]. Gene subunits expressed by STX3 included Itgav, Itga3, Itga5, Itga6, Itgb1 and Itgb5 (Figure 5). These were also expressed by 2RL22. Itga1 was not tested on the U74Av2 Affymetrix genechips and Itga2, Itgb3 and Itgb4 were not detected (Supplementary Table 1). It is therefore possible that STX3 expresses the heterodimers α3β1, α5β1, α6β1, αVβ1 and αVβ5, consistent with the endothelial cell morphology of STX3 stroma.
Expression of fibroblasts markers by stroma
The 2RL22 lymph node stroma comprises cells showing elongated shape, reflective of fibroblastic cells (Figure 1C & 1D). Genes specifically expressed by 2RL22 were suggestive of fibroblast cell type. However, investigation of expression of known fibroblast genes within the Affymetrix database was used to further delineate the cell type of 2RL22. Many reports indicate considerable heterogeneity amongst fibroblasts for cell surface marker expression. Synovial fibroblasts are located within the limb articulation/joint, and express VCAM1, ICAM1, CD40 and CD44 [20]. Circulating fibroblastic precur-sors or fibrocytes express CD11a, CD11b, CD13, CD45, ICAM1, CD58 and CD71 [21]. Fibroblasts also express receptors of the complement pathway [22]. Adventitial fibroblasts and myofibroblasts re-spectively express the cytoskeleton proteins vimentin (Vim) and the α smooth muscle ac-tin (Acta2) [23]. Fibroblast functions include mechanical support for surrounding tissue and production of extracellular matrix proteins and growth factors. Consequently, fibroblasts express sev-eral procollagen proteins of types I, III and V [22], along with the collagen modifying enzyme proline 4-hydroxylase [21]. The fibroblast growth factors Fgf6, Fgf7 and Fgf10 are also expressed by fibro-blasts [24].
Expression of these genes was not detected in either STX3 or 2RL22, and only those expressed in at least one cell line are shown in Figure 4. Both 2RL22 and STX3 showed expression of transcripts for several procollagens of types I, III and V (Col1a1, Col1a2, Col3a1, Col5a1, Col5a2), the α and β subunits of proline 4-hydroxylase (P4hb, P4ha1 and P4ha2) and the cytoskeleton protein Acta2. Amongst the adhesion molecules investigated, only Vcam1 and Cd44 transcripts were expressed by both cell types. The growth factor Fgf7 was expressed at low levels in both cell lines. Lack of expression of vimentin, CD40, ICAM1, Fgf6 and Fgf7, as well as CD45, is shown in Supple-mentary Tables 1 and 2. Although the STX3 splenic stroma appears to contain mostly endothelial cells, it expressed all of these genes as did the 2RL22 stromal line (Figure 4A). The common expression of VCAM-1, Col1a1, Col3a1 and Fgf7 by both stroma was also confirmed by RT-PCR analysis (Figure 4B). One possibility is that STX3 may maintain a minor subset of fibro-blast-like cells. The more likely explanation is that some of these genes are expressed in common by the different cell lineages.
Discussion
Cocultures of bone marrow over STX3 induced DC development and those over 2RL22 stroma induced monocyte/macrophage development. The production of DC in a stromal culture in the absence of added growth factors is a unique model for dendritic cell development, particularly since the DC-like cells produced have a distinct immature myeloid phenotype. The influence of spleen stroma on DC develop-ment is made even more notable since it also supports the production of DC-like cells of the same phe-notype in long term cultures (LTC) of spleen cells [7]. The production of cells of similar CD11c+CD11b+B220−CD8α−MHC-II− phenotype in cocultures of bone marrow over spleen stroma and in spleen stroma-dependent LTC has been consistent over more than 50 experiments. Only myeloid DC and not plas-macytoid-like DC are produced, so supporting a unique role for endothelial elements in spleen in DC hematopoiesis. Our current hypothesis is that certain splenic endothelial cells support the early devel-opment of myeloid DC from progenitors/precursors resident in spleen. Some of these may traffic from bone marrow and differentiate after residence within a spleen endothelial niche.
DC development supported by stroma differs from the development of DC from monocytes in blood or myeloid precursors in bone marrow which respond to cytokines like GM-CSF, IL-4 and TNF-α. Indeed STX3 stroma does not produce any of these factors [11], and their addition to LTC established from GM-CSF−/− mice [25] does not increase the already excellent production of DC by stroma deficient in GM-CSF production. Indeed this type of evidence suggests early devel-opment of DC from a progenitor/precursor restricted to production of only myeloid DC. Furthermore, DC progenitors/precursors of this particular DC subset reside in both spleen and bone marrow, and there appears to be a unique role for a subset of spleen endothelial cells in development of this novel DC subset. Our further analysis of 2RL22 indicates that, in contrast to STX3, it supports development of only monocyte-like cells from precursors also present in bone marrow via production of soluble fac-tors, particularly M-CSF [11]. It does not support the production of DC.
The STX3 stromal line appears to comprise primarily endothelial cells expressing early markers like Flt1, CD34, MCAM and Eng/CD105 (Figure 3). Markers of fully mature endothelial cells like von Willebrand factor (Vwf) (Figure 3) and Weibel-Palade bodies (data not shown) were found to be absent in STX3. Some constitutive markers of the endothelial lineage like CD31, Cdh5 and ICAM2 were also not expressed by STX3, although VCAM1 was expressed in low levels (Figure 4). These data indicate that STX3 comprises immature endothelial cells, some of which could have angiogenic properties. The absence of expression of some of the known constitutive markers of endothelial cells by STX3 suggests that it may reflect a distinct endothelial cell type in spleen. However, the possibility that cultured cells adopt a different gene expression profile to in vivo cell equivalents cannot be ruled out. However such an hypothesis would depend on identification of definitive markers, and in vivo analysis of spleen endothelial-like cells.
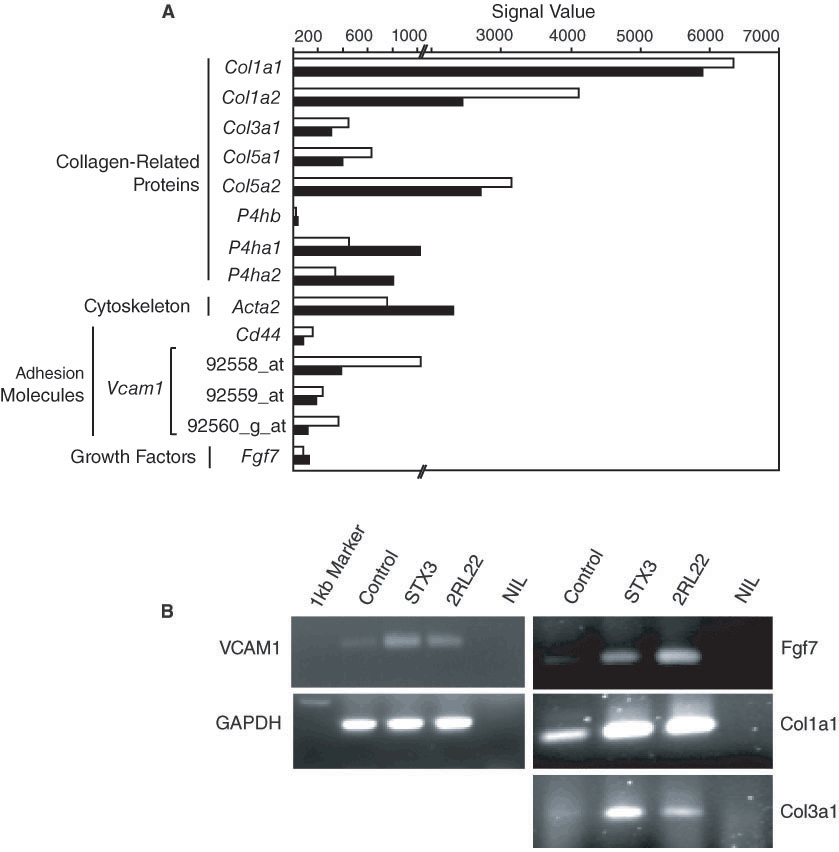
Expression of fibroblast markers by stroma. (
The distinct lymph node stroma, 2RL22, comprised cells with the morphology of fibroblasts. Cells ex-press cytoskeletal genes like Acta2 as well as genes encoding extra-cellular matrix including Col1a1, Col1a2, Col3a1, Col5a1, Col5a2, P4ha1 and P4ha2. However they do not express genes like Vim, C1qr1, Cd40, Fgf6 and Fgf10 which are also associated with fibroblasts. Transcripts of several endo-thelial-related markers were also found to be expressed by 2RL22 stroma including Tal1/SCL, Cd14, Eng/CD105, Cdh5/CD144/VE-cadherin, Ly6a/Sca-1 and Vcam1 (Figures 3 and 4). Several integrins showed gene expression in 2RL22, including Itgav/CD51, Itga3/CD49C, Itga5/CD49E, Itgb1/CD29 and Itgb5 (Figure 5). Existing evidence shows that α5β1 integrin is expressed on fibro-blasts [26]. Although 2RL22 expressed some genes for cell surface markers of the endothelial lineage, none of the genes for endothelial cell-specific surface receptors including Flt1, Kdr, Tek and Tie1 were expressed. Altogether, these results support the view that 2RL22 comprises fibroblast-like cells and not endothelial cells. The expression of some genes expressed by endothelial cells is not surprising given the difficulty of defining the phenotype of connective tissue-forming cells.

Expression of integrins by stroma. Signal values from Affymetrix U74Av2 Genechips repre-senting integrins expressed by either or both STX3 (spleen) and 2RL22 (lymph node) stroma. Calcula-tion of signal value and p-value was made using MAS5.0 software. Probe sets with p-value 0.005 and signal value > 20 were considered as expressed and are identified (*). Open bars: STX3; black bars: 2RL22.
Investigation of many genes expressed by endothelial cells has proven useful in delineating the cell lineage origin of the STX3 stromal cell population. This has been less informative for 2RL22 lymph node stroma. Fibroblast markers investigated were not specific for 2RL22 and some also appeared to be expressed by STX3. In particular, expression of Cd34, Mcam, Eng, Flt1, Vcam1 and genes for several integrins has revealed an immature endothelial cell phenotype for STX3. However, the pattern of gene expression in STX3 is distinct from both the he-mangioblast, and the terminally differentiated and functional endothelial cell.
Recently, Zhang et al. [5] reported splenic endothelial-like cells which support the proliferation of DC of a specific regulatory phenotype. Endothelial cells in that study were prepared from spleen by depletion of CD11b+ myeloid cells followed by positive selection for VCAM-1+ cells. These cells supported further differentiation of immature DC expanded from bone marrow by culture with GM-CSF and IL-4. Following culture above fresh endothelial stroma, these cells differentiated to become suppressive or regulatory DC [5]. Freshly isolated splenic endo-thelial lines described above, differ from STX3 both in terms of their phenotype and in type of DC produced in cultures. The freshly isolated endothelial cells support DC with a distinct phenotype and function reflecting regulatory DC: CD11c+CD11b+MHC-IIloCD45RB+ DC [5]. In contrast, STX3 stroma described here, supports development of DC from progenitors in bone marrow having a distinct immature DC phenotype: CD11c+CD11b+MHC-II−CD45RB−. The phenotype of those cells resembles the DC population produced in long term cultures derived from spleen [7]. These cells are highly endocytic and have limited capacity to activate or induce CD4+ T cell proliferation [27]. Limitations on longevity of STX3 stromal cocultures prevents analysis of the self-renewal capacity of progenitors, although production of DC has been maintained in one coculture for 37 days (data not shown). Evidence to date suggests that no CD45RB+ regulatory DC are produced in cocultures of bone marrow over STX3, which distinguishes this stromal system from those described by others [5,6].
In line with freshly isolated splenic endothelial cells described by Zhang et al [5], STX3 expresses transcripts for Vcam1 (Figure 4) but not for Itgam/CD11b (data not shown). Freshly isolated splenic endothelial cells were shown to have low expression of Pecam1/CD31, Cdh5/CD144, Vwf and ICAM1/CD54 but clear expression of VCAM-1 [5]. STX3, did not express any of these genes except Vcam1 (Figure 4). Absence of expression of Pecam1/CD31 and Vwf in STX3 was confirmed by RT-PCR [5]. Furthermore, no Weibel-Palade bodies were detected by electron microscopy either in freshly isolated splenic endothelial cells [5] or in cloned stromal cell lines derived from STX3 [4]. While some common endothelial-like cell properties can be delineated, the hematopoietic support capacity in terms of type of DC produced above stroma, was quite distinct. It is therefore possible that different endothelial cell types may exist in spleen which have distinctive function in that they support hematopoiesis of DC of different phenotype and function.
Acknowledgments
This work was supported by a grant to TO and HO from the Australian National University. GD was supported by a Ph.D. fellowship from the Fonds de la Recherche en Santé du Québec, Québec, Canada.
Disclosures
The authors report no potential conflict of interest.