
Abstract
Previously, we reported that the spindle assembly checkpoint (SAC), which is coupled in somatic cells, is uncoupled from apoptosis-initiation in mouse and human embryonic stem cells (ESCs). This condition allows ESCs to tolerate and proliferate as polyploidy/aneuploid cells. Proper function of the SAC is vital to prevent polyploidy/aneuploidy during ex vivo hematopoietic stem cell (HSC) expansion. Here we address, for the first time, whether HSCs are more like ESCs or somatic cells with respect to SAC–apoptosis coupling. Using multiparametric permeablized cell flow-cytometric analysis to identify and analyze the mouse sca 1+/c-kit+/lin− (LSK) population, we found the mitotic spindle checkpoint to be functional in primary murine LSK cells, a population enriched in primitive hematopoietic stem/progenitor cells, after prolonged activation of the SAC by microtubule-depolymerizing agents such as nocodazole. HSCs can efficiently initiate apoptosis after activation of the SAC in LSK cells as indicated by increased hypodiploidy and increased levels of activated caspase 3, suggesting that HSCs behave more like somatic cells instead of ESCs with respect to this important cell cycle checkpoint. We conclude that mouse HSCs are not subject to the same kinds of chromosomal instability as are ESCs, knowledge that might aid in optimizing in vitro culture and expansion of human bone marrow or cord blood HSC for clinical applications.
Introduction
During cell division, the proper segregation of chromosomes into daughter cells is essential for survival. Accurate transmission of the genome is ensured both by intrinsic properties of the cell cycle machinery and by a series of checkpoints [1]. The entry into mitosis is controlled by checkpoints that monitor DNA damage and the replicative state of DNA while the exit from mitosis is controlled by checkpoints that monitor assembly and position of the mitotic spindle [1]. The mitotic spindle checkpoint, also known as the spindle assembly checkpoint (SAC), helps to ensure the proper segregation of chromosomes into daughter cells during mitosis. Failure of spindle attachment to a single kinetochore activates the SAC, which arrests cells at metaphase until chromosomes are equally distributed [2–4]. Once all kinetochores are attached and chromosomes are aligned on the metaphase plate, the SAC is silenced and cells progress into anaphase [5].
Chromosomal instability (CIN) leading to an aberrant chromosome number (aneuploidy) is a hallmark of many cancers [6,7]. The mechanism and molecular determinants promoting CIN are not clearly un-derstood [7]. However, there is increasing evidence that suggests a role of the SAC in promoting ane-uploidy and tumorigenesis [8]. Efficiency of the SAC is particularly important in embryonic and adult stem and progenitor cells, which generate many populations of differentiated cells. Understanding ge-nome maintenance and possible causes of CIN in these populations is necessary to ensure their safe and effective use.
Earlier, we reported that the SAC is uncoupled from apoptosis initiation in mouse and human embry-onic stem cells (ESCs) [9]. Embryonic stem cells exit mitosis and become polyploid after prolonged activation of the SAC [9]. Embryonic stem cells accumulate in M phase of the cell cycle after micro-tubule disruption with nocodazole, but exit mitotic delay and reenter a polyploid mitosis. This is in con-trast to normal somatic cells and cell lines with normal p53 responses, which do not respond to pro-longed activation of the SAC in manner. These cells initiate apoptosis after exiting the cell cycle in a G0/G1-like state with 4C DNA content, often in two nuclei, or enter senes-cence [10,11]. These data, along with findings of increased pseudomitotic index in neural and hemato-poietic progenitor cells [12], suggest that the SAC might also be uncoupled from apoptosis in hemato-poietic stem cells (HSCs). Proper function of the SAC is vital to prevent polyploidy/aneuploidy during ex vivo culture and expansion of HSC and their progenitors.
Here we address, for the first time, whether HSCs are more like ESCs or somatic cells with respect to SAC–apoptosis coupling. We conclude that mouse HSCs are not subject to the same kinds of chromosomal instability as are ESCs.
Methods and Materials
Isolation and culture of low density mononuclear cells (LDMCs) from murine BM
Murine BM was isolated from both femurs and tibias of C57/BL6 mice (Jackson Laboratories) using aseptic procedures. Mononuclear cells were isolated by Lympholyte®-M separation. Cells were washed three times with PBS and resuspended in RPMI + 10% FBS + 100 ng/ml each of TPO, Flt3, and SCF (BioVision, Mountain View, CA, USA) and then incubated at 37°C for 24 h before treatment.
Antibodies and drugs
Antibodies used for primary and secondary surface staining, as well as their isotype controls, were from BD Biosciences (San Jose, CA, USA) and BioLegend (San Diego, CA, USA). Primary and secondary antibodies for intercellular staining were from BD Biosciences, Cell Signaling Technology (Danvers, MA, USA), and Invitrogen (Carlsbad, CA, USA). Propidium iodide and Hoechst 33342 used for counterstaining were obtained from Invitrogen. Nocodazole was obtained from Sigma Chemical (St. Louis, MO, USA). The drug was dissolved and diluted in DMSO, and the concentration of Nocodazole used in experiments was 1 ng/ml.
Multivariate permeablized flow cytometry
After harvest and washing with PBS, single cell suspensions were stained with primary antibodies, fol-lowed by staining with secondary antibodies each for 30 min in the dark. Cells suspensions were washed three times with PBS + 1% BSA after primary and secondary staining. Cells were permeablized and fixed using Cytofix/Cytoperm (BD Biosciences) and then stained with various antibodies, washed with Perm/Wash Buffer (BD Biosciences), and counterstained. Flow-cytometric data were acquired us-ing LSRII (Becton-Dickenson) flow cytometer. Data was analyzed with WinList program (Verity Soft-ware House; Topsham, ME, USA).
Results
Nocodazole treatment causes cell cycle arrest in mHSCs
To see if hematopoietic stem/progenitor cells (HSPCs) function like ESCs with respect to check-point–apoptosis coupling, we examined the function of the mitotic/SAC. Mononuclear cells iso-lated from murine bone marrow were cultured in the cytokine cocktail of TPO, Flt3, and SCF for 24 h before addition of nocodazole, a microtubule-depolymerizing agent that was used to activate the SAC in mononuclear cells isolated from murine BM. Cells were incubated for 48 h after nocodazole treatment, then stained and analyzed through flow cytometry. Within the mononuclear fraction of cells, DNA con-tent and levels of phospho-histone H3 were examined in sca 1+/c-kit+/lin− (LSK) cells, a population highly enriched for HSCs. Hematopoi-etic stem cells accumulate in M phase of the cell cycle after microtubule disruption with nocodazole treatment (Fig. 1A). This is evident by the increase of phosphorylation at histone H3, which is tightly correlated with chromosome condensation during mitosis and therefore serves as a marker for mitosis [13]. This population of cells with higher levels of phospho-histone H3 appears to have 4C DNA con-tent, also indicative of cell cycle arrest in M phase (Fig. 1B).
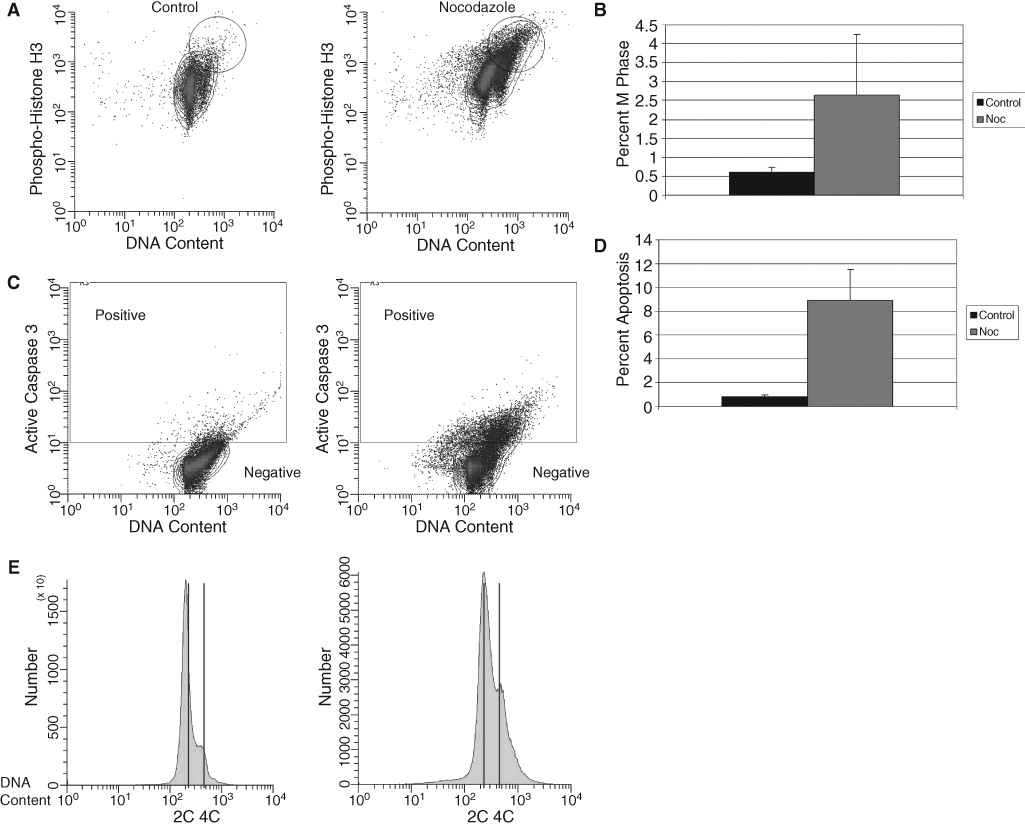
Microtubule-induced mitotic arrest in murine HSCs. (
HSCs initiate apoptosis after SAC activation
To further examine if HSPCs function like ESCs with respect to checkpoint–apoptosis coupling, we investigated apoptotic responses in HSPCs after SAC activation. Upon visual inspection, fewer no-codazole-treated cells were observed when compared to the control nontreated cells although equal numbers of cells were initially present in both groups. After SAC activation via nocodazole treatment, increased levels of activated caspase 3 (Figs 1C and D) and an increase in hypodiploidy (Fig. 1E) were observed in the treated LSK cells as compared to untreated cells. This is indicative of apoptosis activa-tion in this population of cells. These data suggest that HSC can efficiently initiate apoptosis after acti-vation of the SAC, indicating that checkpoint activation is coupled with apoptosis.
Discussion
In this study, we demonstrate that HSCs show an arrest in the M phase of the cell cycle in response to treatment with nocodazole, a microtubule-depolymerizing agent. This arrest, shown by an increase in phospho-histone H3 in nocodazole-treated cells, implies that the SAC functions in this population of cells. Also, in response to nocodazole, HSCs initiate apoptosis, evident by increased levels of activated caspase 3, subdiploid levels of DNA, and decreases in cell number. These data lead to the conclusion that HSCs exhibit coupling of apoptosis to activation of the SAC, a condition observed in somatic cells. This is in contrast to ESCs, which accumulate in M phase after nocodazole treatment, but then exit this mitotic delay and reenter a polyploid mitosis [9]. We also conclude that HSCs do not function like ESCs, which exhibit uncoupling of apoptosis from checkpoint activation and tolerance to tetraploidy/aneuploidy [9].
Differentiated cells from many tissues have a relatively short life span and are continuously replaced by new cells generated from stem and progenitor cells [12]. For example, in the human bone marrow ∼200 billion erythrocytes and 70 billion neutrophilic leukocytes are produced and released in the bloodstream every day [14]. The efficiency of the SAC is particularly important in HSCs, since these cells give rise to many different types of progenitor cell populations, including the myeloid and lymphoid lineages of cells. Uncoupling of SAC activation from apoptosis could lead to chromosome instability not only in HSCs themselves, but also in all the progenitor populations descending from these cells. As previously described, chromosome instability leading to aneuploidy characterizes many cancers [6]. Therefore, chromosome instability in HSCs and their progenitors could lead to cancers derived from these cell types, such as leukemia or lymphoma.
Ex vivo culture and expansion of hematopoietic stem and progenitor cells could prove problematic in a situation of checkpoint–apoptosis uncoupling. In numerous settings, ex vivo manipulation of HSPCs occurs before the cells are used in research or clinical applications. If these cells exhibit defects in checkpoints and/or apoptosis initiation as previously suggested [9,12], cells with genomic abnormali-ties could accumulate and negatively affect the outcome of experiments or treatments. However, the observation that HSCs efficiently initiate apoptosis after SAC activation indicates that these cells will undergo programmed cell death if problems arise during mitosis, lessening concerns that the stress of ex vivo expansion of HSCs could cause instability in HSCs that might lead to disordered growth/survival after transplantation of the expanded cells.
Acknowledgments
These studies were supported by Public Health Service Grants: RO1 HL67384, RO1 HL56416, and a Project in PO1 HL53586 to HEB from the National Institutes of Health (NIH). SR is supported by NIH Training Grant T32 CA111198.
Contribution Statement
Sara Rohrabaugh: Designed and performed the research, collected and analyzed data, and wrote the pa-per.
Charlie Mantel: Designed the research and analyzed the data.
Hal E. Broxmeyer: Worked on the paper and supported the studies through intellectual input and fund-ing.