
Abstract
Embryonic stem cell (ESC)–based transplantation is considered a promising novel therapy for a variety of diseases. This is bolstered by the suggested immune-privileged properties of ESCs. In this study, we used in vivo bioluminescent imaging (BLI) to non-invasively track the fate of transplanted murine ESCs (mESCs), which are stably transduced with a double fusion reporter gene consisting of firefly luciferase (FLuc) and enhanced green fluorescent protein (eGFP). Following syngeneic intramuscular transplantation of 1 × 106 mESCs, the cells survived and differentiated into teratomas. In contrast, allogeneic mESC transplants were infiltrated by a variety of inflammatory cells, leading to rejection within 28 days. Acceleration of rejection was observed when mESCs were allotransplanted following prior sensitization of the host. Finally, we demonstrate that the mESC derivatives were more rapidly rejected compared to undifferentiated mESCs. These data show that mESCs do not retain immune-privileged properties in vivo and are subject to immunological rejection as assessed by novel molecular imaging approaches.
E
Clearly, the extent to which immunologic rejection of ESCs and/or ESC derivatives occurs following allogeneic transplantation is a crucial question that has not yet been clarified. The results of aforementioned studies were primarily based on immunohistological evaluation, which provide only a “snapshot” representation rather than a comprehensive picture of cell survival over time [16]. Such limited techniques may partly contribute to the conflicting observations of mESC survival in allogeneic or xenogeneic hosts. To address these limitations, we have previously validated in vivo bioluminescent imaging (BLI) to be a reliable technique for assessing engraftment and survival of mESCs following transplantation in vivo [17]. An important advantage in using BLI to track cell transplantation is that the expression of the firefly luciferase (Fluc) reporter gene, which is integrated into the DNA of the transplanted cells, is expressed only by living cells, making it a highly accurate tool for following cell graft rejection in the living subject [18]. We have previously shown that the transduction of mESCs with reporter genes that allow in vivo tracking does not alter cellular viability and their ability to differentiate into cells of all three germ layers [19, 20]. To further delineate the novel field of ESC transplantation immunobiology, we used in vivo BLI to visualize patterns of survival and rejection of mESCs following transplantation across histocompatibility barriers.
To track the mESCs in vivo by BLI as well as ex vivo by immunohistochemistry, a double fusion (DF) reporter gene construct carrying Fluc and enhanced green fluorescent protein (eGFP) driven by a constitutive ubiquitin promoter (pUB) was transduced into undifferentiated mESCs (D3 cell line, derived from the 129/Sv mouse strain), using a self-inactivating (SIN) lentiviral vector (Fig. 1A), as described previously [20]. After transduction, mESCs robustly expressed Fluc (Fig. 1B) and eGFP (Fig. 1C). In agreement with the previous reports [9, 21], flow cytometric analysis showed the expression of pluripotency marker SSEA-1 and minimal expression of H-2kb MHC class I (MHC-I) or I-Ab MHC class II (MHC-II) antigens on undifferentiated mESCs. In contrast, mESC-derived teratoma cells (mES–TCs) isolated from 6-week-old surgically explanted teratomas by a 2-h enzymatic digestion in Collagenase D (2 mg/mL in RPMI medium) clearly expressed both MHC-I and MHC-II antigens (Fig. 1D). In culture, mES–TCs displayed a heterogeneous single cell morphology, as opposed to mESCs, which grow in tight clusters (Fig. 1E).
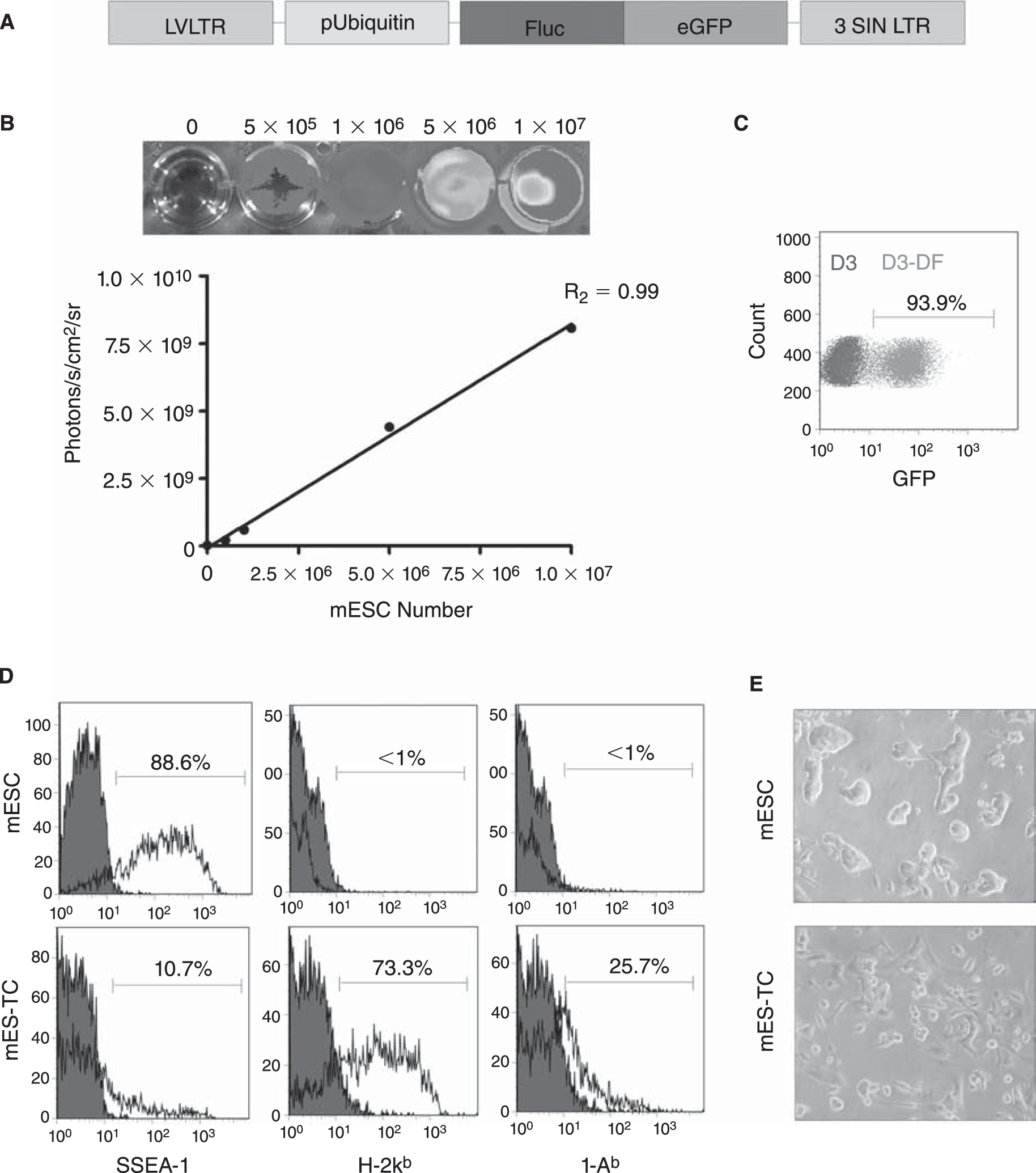
Characterization of the double fusion (DF) firefly luciferase (Fluc) and enhanced green fluorescent protein (eGFP) transduced D3 murine embryonic stem cells (mESCs). (
Transduced mESCs (1 × 106) were transplanted by direct injection into the gastrocnemius muscle of syngeneic (129/Sv, H-2kb, n = 8) and allogeneic (BALB/c, H-2kd, n = 8 recipient mice, after which BLI was performed weekly. The BLI protocol involved intraperitoneal administration of the
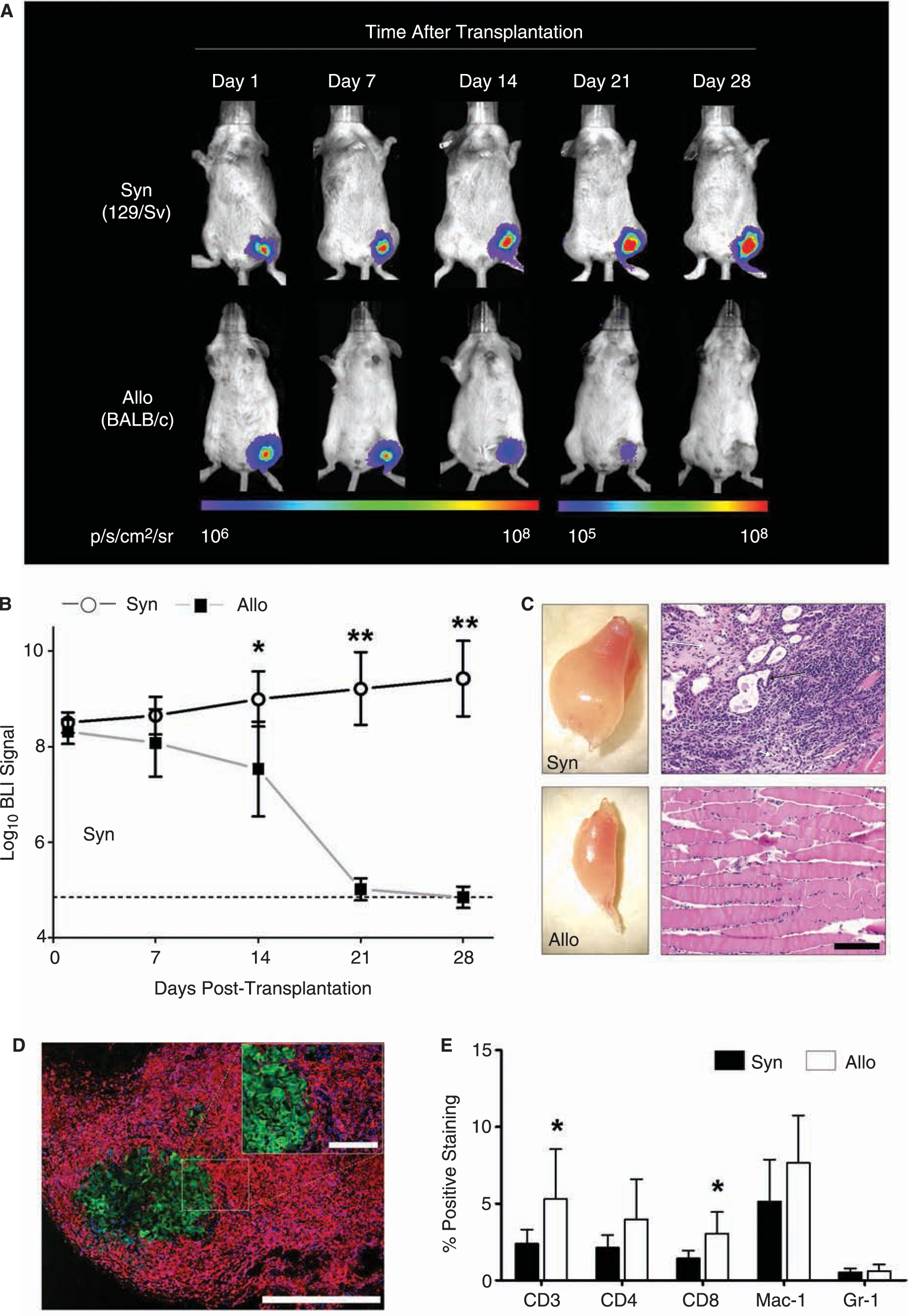
In vivo bioluminescent imaging (BLI) of transplanted mESCs. (
We next investigated the local immune response elicited by mESCs in a second group of allogeneic BALB/c and syngeneic 129/Sv mice (n = 8 per group). At 10 days following transplantation, the muscles of these mice were explanted and investigated for graft-infiltrating cells. Immunofluorescent staining of allogeneic muscle sections demonstrated massive infiltration of CD45+ inflammatory cells surrounding the eGFP+ mESC grafts (Fig. 2D). Quantification and further characterization of graft-infiltrating cells was carried out by enzymatic digestion (2 h in 2 mg/mL collagenase D in RPMI medium) of the explanted muscles followed by FACS analysis. Live cells were gated out from debris and further analyzed for the expression of a panel of hematopoietic cell surface markers. Comparison of the syngeneic to the allogeneic mESC recipient muscles confirmed severe infiltration of various types of immune cells involved in both adaptive and innate types of immunity (Fig. 2E). At this time-point, both the CD3+ cells (total T-cell population) and CD8+ cells (cytotoxic T-cells) were present at a significantly higher frequency in allogeneic versus syngeneic recipients (p < 0.05 by t-test), suggesting a prominent role for T-cell-mediated immunity in mESC rejection.
To investigate whether mESCs could induce immunologic memory, we next performed infusion of 1 × 106 nontransduced D3 mESCs or 1 × 106 irradiated (3,000 rad) 129/Sv splenocytes (as control) into the tail vein of allogeneic BALB/c recipients (n = 4 and 5 per group, respectively). Four weeks later, these same mice received intramuscular transplantation with the same number of transduced mESCs and cell survival was quantified using longitudinal BLI. Accelerated BLI signal loss was observed in both groups, with BLI signal reaching background levels as soon as post-transplant day 4 (control group) and day 7 (mESC group) (Fig. 3A and B). These findings confirm that a donor (129/Sv)-specific adaptive immune response is generated against transplanted mESCs.
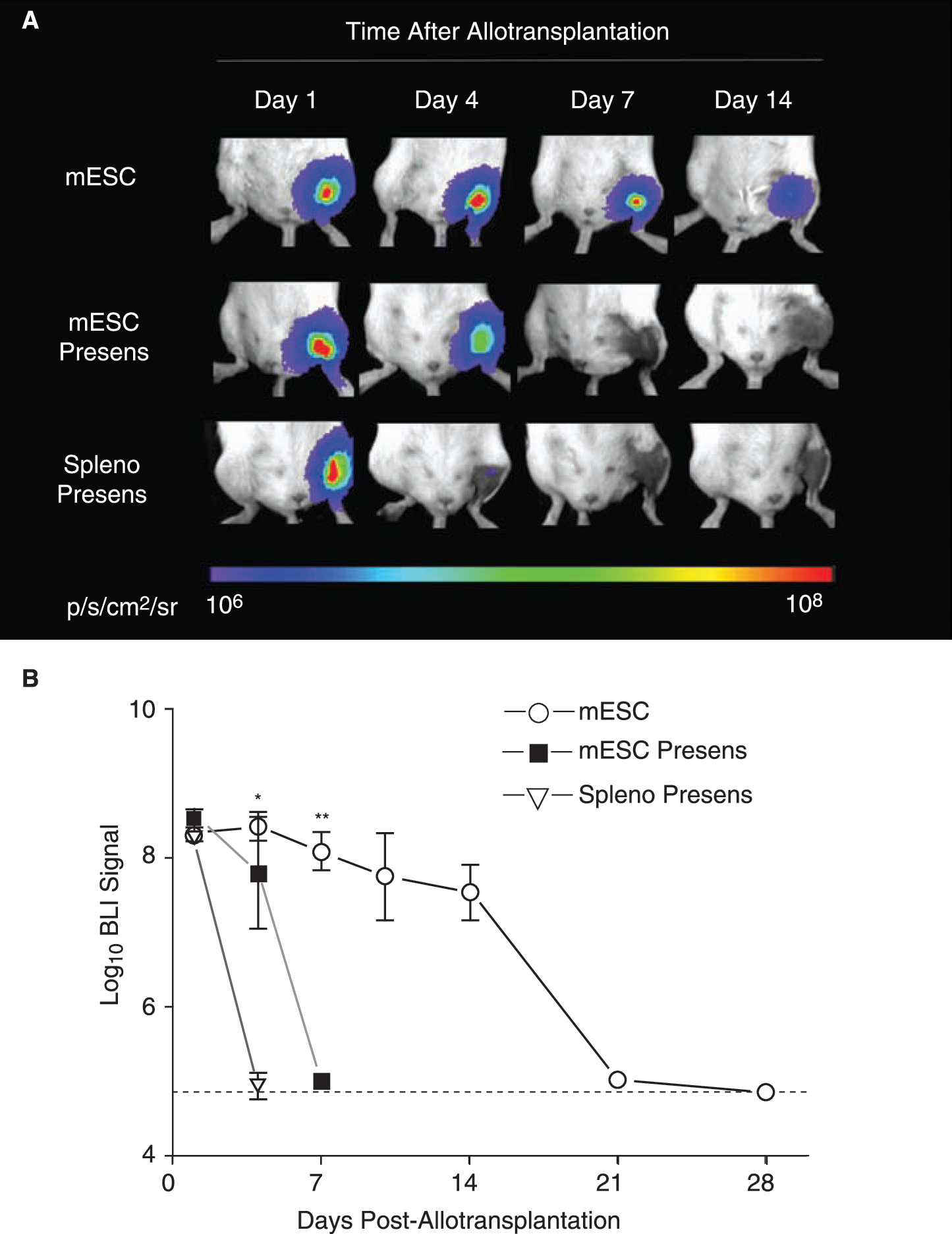
Accelerated rejection following presensitization of the host. (
Clinical ESC-based transplantation protocols will require differentiation of the donor cells into the desired cell type before transplantation, due to the risk of teratoma formation following transplantation of undifferentiated ESCs [13]. Therefore, we thought that it would be relevant to study the survival of mESC derivatives following allotransplantation. A group of BALB/c mice (n = 5) was injected with 1 × 106 mES–TCs and longitudinal BLI was performed. In comparison with undifferentiated mESCs, the BLI signal displayed a similar pattern up until 7 days following transplantation (Fig. 4A and B). However, BLI signal in the mES–TC animals decreased substantially by day 10 (mESC 7.74 ± 0.58 vs. mES-TC 5.46 ± 0.17 Log[photons/s]; p < 0.01 by t-test) and reached background levels at day 14. These results show that differentiated mESCs have impaired survival capacity as compared to undifferentiated mESCs when transplanted over histocompatibility barriers.
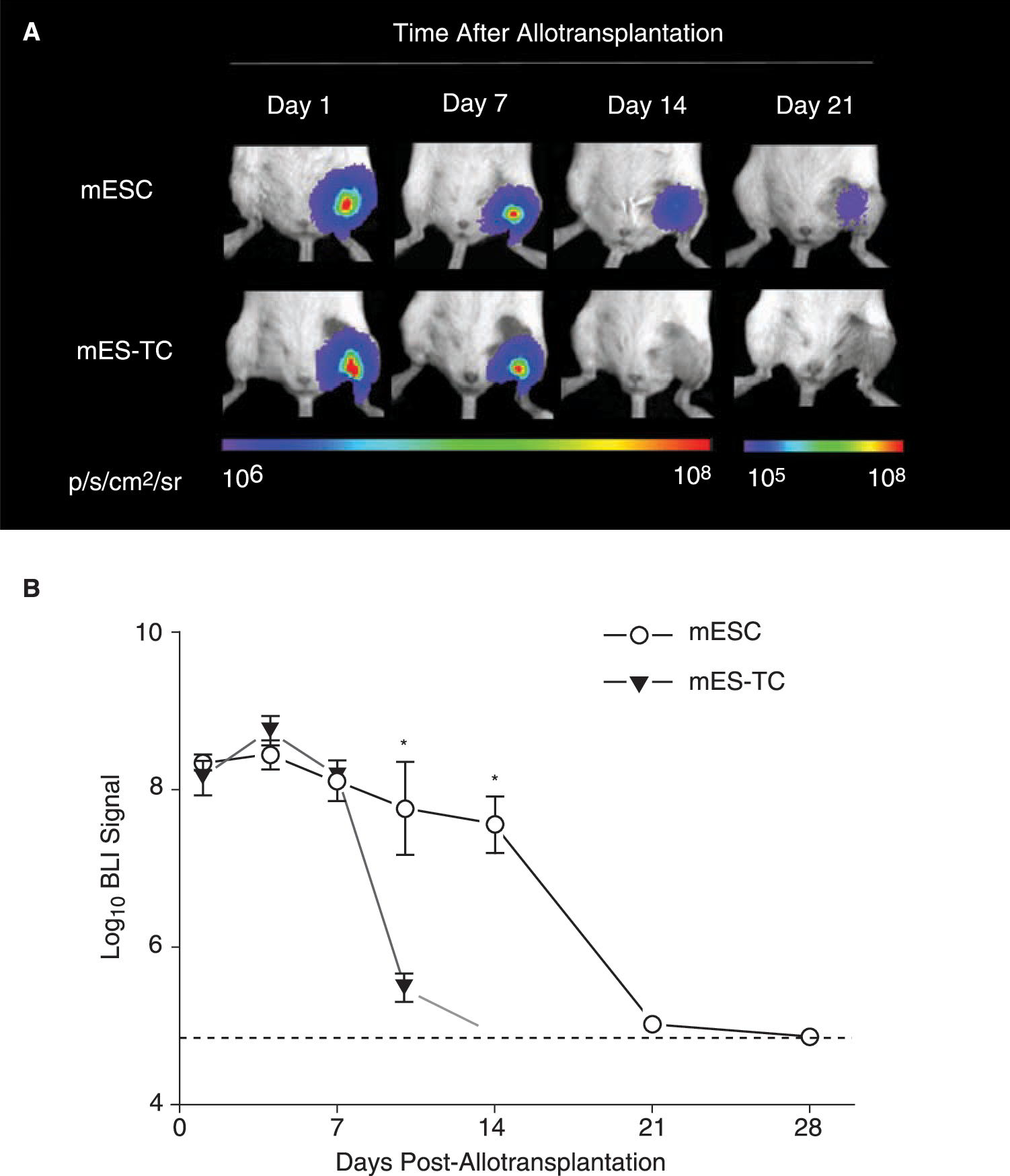
Impaired survival capacity of differentiated mESCs as compared to undifferentiated mESCs following allotransplantation. (
In a recent report, higher numbers of transplanted undifferentiated mESCs (5 × 106–20 × 106) were shown capable of survival and formation of teratoma in allogeneic recipients [22]. Similarly, in this study we found that when 10 × 106 mESCs were transplanted into allogeneic BALB/c recipient muscles, BLI signal could be followed out beyond 28 days following transplantation in 2 out of 10 animals (20%) and intramuscular teratomas could be observed in these two animals. However, when mESCs were allowed to differentiate for 14 days in vitro prior to transplantation, 10 × 106 cells were rejected by 10 out of 10 animals (100%) (data not shown).
Undifferentiated mESCs have been reported to possess immune-privileged properties, which has been attributed to the low expression of MHC molecules [21], expression of perforin-deactivating Serpin-6 [9] and/or production of lymphocyte-inhibiting TGF-β [22] by the cells. However, our study clearly shows that the transplantation of mESCs in vivo can result in donor-specific immune recognition and rejection, which leads to immunologic memory. Importantly, post-transplant cell death is accelerated when mESCs are allowed to differentiate before transplantation, a scenario likely to occur in clinical ESC-based transplantation settings in the future. Whether the latter phenomenon is truly a result of increased immunogenicity of the cells, as suggested by their increased expression of MHC antigens (Fig. 1E), or if other factors such as a decreased proliferation rate of differentiated cells play part in this process remains to be evaluated in detail. Nevertheless, the results of this study emphasize that immunologic rejection of ESCs or ESC derivatives is likely to occur and the need exists for solutions that reduce or eliminate immunologic response following ESC-based transplantation [23]. To develop and test such strategies, there will also be a need for reliable imaging technologies to track and assess behavior of the cells following transplantation. In vivo BLI serves these needs and has the potential to play a prominent role in future ESC-based transplantation research.
Footnotes
Acknowledgments
This work was supported by grants from the AHA BGIA, BWF, California Institute of Regenerative Medicine (CIRM) RS1-00322, NHLBI R21HL089027 (J.C.W.), the ISHLT Research Grant (S.S.), and by the ESOT-Astellas Study and Research Grant (R.J.S.). The contents of this publication are solely the responsibility of the authors and do not necessarily represent the official views of CIRM or any other agency of the state of California.