
Abstract
Mesenchymal stem cells (MSCs) are multipotent progenitor cells found in bone marrow that have the capacity of differentiating into bone, cartilage, fat, muscle, and other tissues. Chicken MSCs were isolated from 1- to 14-day-old chickens. Microscopically, the cultured cells showed morphology resembling fibroblasts and divided actively. Chicken MSCs expressed the transcription factors PouV, Sox2, and Nanog, which have been shown to be critical for stem cell self-renewal and pluripotency. The multilineage differentiation potential of chicken MSCs was revealed by their ability to undergo adipogenic, osteogenic, and chondrogenic differentiation. Like mammalian MSCs, chicken MSCs also had immunoregulatory activity and inhibited in vitro mitogenic response of T cells. The inhibition of mitogenic response of T cells correlated with the production of nitric oxide (NO) in cultures containing MSCs and T cells. Our data show for the first time that MSCs can be isolated from postnatal chicken bone marrow and these cells are capable of in vitro multiplication and multilineage differentiation, thus making them a suitable model in the field of stem cell research.
Introduction
M
MSCs constitute a small population (∼0.001–0.01% nucleated cells) of adult human bone marrow cells. MSCs have been isolated from several tissues including adipose tissue, skeletal muscle, synovium, spleen, thymus, blood, lung, fetal blood, and amniotic fluid [10]. These cells have the ability to produce cytokines and growth factors that support and regulate haematopoiesis [11,12]. Several studies using a variety of animal models have shown that MSCs may have a role in the repair and regeneration of infarcted myocardium, damaged bone, tendon, and cartilage [13 –16]. These characteristics indicate that MSCs can be used as powerful tools in reconstructive medicine. Pluripotent stem cells are an attractive tool for tissue engineering and regenerative medicine.
MSCs isolated from humans as well as from other mammalian species have immunoregulatory function and inhibit inflammation and immunological responses both in vitro and in vivo. MSCs suppress T-cell proliferation probably via mechanisms that are independent of major histocompatibility complex (MHC). Several soluble factors such as transforming growth factor-β (TGF-β), hepatocyte growth factor, indoleamine 2,3-dioxygenase (IDO), nitric oxide (NO), and prostaglandin E2 (PGE2) have been reported to mediate T-cell suppression by MSCs [10]. MSCs improve the outcome of allogeneic transplantation by promoting hematopoietic engraftment [17] and also reduce graft-versus-host disease [18,19]. In vivo infusion of MSCs improved the course of chronic autoimmune encephalomyelitis in multiple sclerosis and other neurodegenerative disorders [20].
Chicken is the first farm animal with a completely sequenced genome. Because of its in ovo embryonic development rather than in utero, chicken is an important model for embryology and development studies. Chicken serves as a model organism for the study of viruses and cancer. Rous sarcoma virus, the first tumor virus, and oncogene (src) were identified in the chicken. The chicken immune system provided the first indication of the distinctions between T and B cells, with the B-cell nomenclature based on the avian bursa of Fabricius [21]. Thus, chicken is an important model system for studies of evolution, embryology, immunology, oncology, cell biology, virology, and gene regulation.
In the present study, we have, for the first time, isolated and characterized MSCs from hatched specific pathogen-free chickens. Chicken MSCs may be useful as a model system for studies on development, physiology, and disease.
Materials and Methods
Isolation and expansion of MSCs
Femur bones were obtained from 1- to 14-day-old specific pathogen-free chickens. MSCs were isolated by previously described methods [2,22]. In brief, the tip of each bone was removed and the marrow was harvested by inserting a syringe needle into one end of the bone and flushing with Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Carlsbad, CA). The bone marrow cells were filtered through a 70-µm nylon mesh filter (BD, Falcon, San Jose, CA) and mononuclear cells were obtained by density gradient centrifugation over Ficoll-Hypaque (gradient density 1.090). Cells (1–5 × 105/cm2) were plated in 25-cm2 cell culture flasks in DMEM containing 10% fetal bovine serum (FBS; Atlas Biological, CO), 2 mm
Colony-forming unit-fibroblast (CFU-F) assay
The colony-forming potential of MSCs was tested by plating 100 or 1,000 cells in a 60-mm2 cell culture dish. After 7–8 days of incubation at 37°C, the cells were fixed and stained with 1% crystal violet in methanol for 30 min. All visible colonies greater than 3 mm were counted [22].
Proliferation of MSCs
The proliferation rate of chicken MSCs was determined by cell count. For cell count: a total of 1 × 104 cells were plated in a 24-well plate and a total of three replicas were prepared for each observation. At intervals, the cells were trypsinized and a total cell count was determined by Trypan blue dye exclusion method.
Differentiation assays
Adipogenic differentiation. Adipogenic differentiation was assayed by the method of Pittenger et al. [2 ]. In brief, ∼104 cells/well in 24-well plates were cultured in expansion media until the cells reached 70%–80% confluence. Adipogenic differentiation was induced by culturing confluent MSCs cultures in DMEM supplemented with 10% FBS, 1 µM dexamethasone, 10 µg/mL insulin, 0.2 mM indomethacin for 21 days. The media was completely replaced every 3–4 days. Adipogenic potential was assessed by Oil Red O staining. The cells were washed with PBS and fixed with 10% formalin for 30 min at room temperature. Then the cells were incubated with filtered 0.3% Oil Red O (Sigma-Aldrich, St. Louis, MO) in 60% isopropanol for 30 min. After three washes in PBS, the cells were photographed.
Osteogenic differentiation. Osteogenic differentiation was assayed by the method of Pittenger et al. [2 ]. In brief, MSCs were seeded in 24-well plates at a density of ∼104 cells/well and cultured in expansion media until the cells reached 90%–100% confluence. Osteogenic differentiation was induced by culturing the cells for 21 days in medium consisting of 100 nM dexamethasone, 10 mM β-glycerophosphate (Sigma-Aldrich, St. Louis, MO), 0.05 mM
Chondrogenic differentiation. Chondrogenic differentiation was assayed by the method of Pittenger et al. [2 ] with minor modifications. The chondrogenic induction of chicken MSCs was examined using high-density pellet cultures (2 × 105 cells/pellet, 500 × g for 5 min) in DMEM medium supplemented with 50 mM ascorbic acid, 0.5 mg/mL insulin (Sigma). Recombinant human transforming growth factor-β1 (Invitrogen, Carlsbad, CA) was added to the pellet cultures at a final concentration of 10 ng/mL. The medium was replaced every 2–3 days for 21 days. Sulfated glycosaminoglycans were detected by 1% Alcian blue staining. The pellet was washed with PBS, fixed with 10% formalin for 30 min at room temperature, washed three times with distilled water, and incubated in 1% Alcian blue (pH 1) (Sigma-Aldrich) overnight. The cell pellet was washed three times with distilled water and photographed.
RNA extraction and RT-PCR
RT-PCR was performed to examine the expression of transcription factors and to confirm gene expression indicative of cell differentiation [2,23]. RNA was isolated using the TRIzol total RNA isolation reagent (Life Technologies, Rockville, MD). The extracted RNA (1 µg) was reverse transcribed by adding 5 mM of random hexamer oligonucleotides (Life Technologies), 200 units of SuperScript reverse transcriptase (Life Technologies), 0.5 mM deoxyribonucleotide triphosphates (dNTP) (Life Technologies), and 10 mM dithiothreitol (Life Technologies). The cDNA was then amplified by PCR using primer sequences shown in Table 1. After PCR, 10 µL of the reaction mixture was subjected to electrophoresis on a 1.5% agarose gel, and the PCR products were visualized by ethidium bromide staining.
T-cell proliferation assay
Splenocytes were prepared by centrifugation over Ficoll-Hypaque gradients [24]. The cells were suspended in triplicate in 100 µL RPMI 1,640 medium supplemented with 5% FBS (Sigma), 2 mM
ConA was purchased from Sigma (St. Louis, MO). Con A was used at 5 µg/mL. PMA and ionomycin were from Sigma and were used at concentrations of 50 ng/mL and 1 µg/mL, respectively.
Measurement of NO production
NO production was determined by measurement of nitrite in the medium as described earlier [25]. In brief, 50 µL of the spent medium from the cultures mentioned in the T-cell proliferation assay was mixed with an equal volume of 1:1 mixture of 1% sulfanilamide in water and 0.1% N-1-naphthylethylenediamine dihydrochloride in 5% phosphoric acid. The absorbance was then read at 570 nm. Sodium nitrite dissolved in the culture medium was used to construct the standard curve. NO concentration was measured in micromolars (µM). All experiments were performed in triplicates.
Statistical analysis
Statistical analysis was performed by analysis of variance (ANOVA) followed by comparison of individual means by Tukey’s test. P < 0.05 was considered to be statistically significant.
Results
Cell culture
We isolated chicken MSCs from the bone marrow based on their characteristic property of attachment to a plastic surface. MSCs showed a fibroblast-like morphology. The initial adherent spindle-shaped cells appeared as individual cells on the third day of culture. Within 2 weeks, the cells reached 70%–80% confluency (Fig. 1A). To confirm that the isolated cells had MSCs characteristics, we examined their differentiation capacity and immune-modulating capabilities as shown below.

Morphology, colony formation, and proliferation potential of chicken mesenchymal stem cells (MSCs). (
CFU assay
MSCs isolated from humans and other animal species have the ability to self-renew and propagate. The CFU assay was used to assess the proliferation and colony-forming potential of chicken MSCs. Chicken MSCs formed 30 ± 8 and 65 ± 10 colonies when plated at 100 and 1,000 cells per 60-mm culture dish, respectively (Fig. 1B).
The proliferation potential of MSCs is shown in Figure 1C. These cells are continuously in culture for >8 months. These data indicate that chicken MSCs are capable of self-renewal and have high proliferation property.
To examine the phenotype of chicken MSCs, we performed RT-PCR using specific primers for various markers (Table 1; Fig. 1D). The cells showed positive reactions for CD44, CD90 (Thy-1), and CD105 (Endoglin), but did not express CD45 indicating that these cells originated from mesenchymal lineage cells.
Expression of PouV, Sox2, and Nanog transcription factors in chicken MSCs
Several transcription factors are known to regulate the development and differentiation of MSCs. In mammals, MSCs pluripotency is under the control of transcription factors that include Oct4 [26], Nanog [27], Sox2 [28], and FoxD3 [29]. Oct4, the octomer-binding transcription factor 4, is an important binding transcription factor present in undifferentiated embryonic and adult stem cells with a high proliferative capacity. We detected transcription factors, PouV, Sox2, and Nanog, in chicken MSCs but not in skin fibroblasts. PouV is a chicken homolog of mammalian Oct4 and like Oct4 has been shown to regulate the pluripotency and self-renewal of chicken embryonic stem cells [30]. As shown in Figure 2, like mammalian MSCs [23,31], PouV, Nanog, and Sox2 gene transcripts were expressed in chicken MSCs for as long as eight passages in vitro (highest passage level tested).

Transcription factor expression in chicken MSCs. Total cellular RNA was extracted from mesenchymal stem cells (MSCs) and subjected to RT-PCR by using specific primers to PouV, Sox2, Nanog, or β-actin. PouV, Sox2, and Nanog gene expression was detected up to passage 8. Total RNA prepared from chicken skin fibroblasts (SF) was used as a negative control.
Differentiation of chicken MSCs
To examine the multipotentiality of chicken MSCs, the cells were cultured in the media specific for the induction of adipocytes and osteocytes.
For the generation of adipocytes, 70%–80% confluent cultures were incubated with adipocytes induction media. After 21 days of incubation, differentiation of MSCs into adipocytes was determined by staining with Oil Red O. Differentiated cells contained multiple lipid vacuoles (Fig. 3A and B). The accumulation of lipid vacuoles in the cells was first detected between 48 and 72 h after the addition of adipogenic medium. The expression of adipogenic-specific genes, aP2 and PPAR-γ, was induced in the adipocytes, thereby confirming the adipogenic differentiation of MSCs (Fig. 3E).
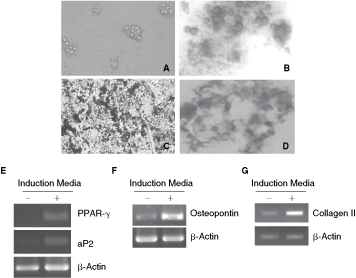
Multilineage differentiation of chicken mesenchymal stem cells (MSCs) in vitro. (
Following incubation of MSCs in osteogenic media, tightly packed colonies forming nodule-like structures were observed. Deposition of calcium in these cells was demonstrated by staining with von Kossa stain (Fig. 3C). The osteocyte-specific gene, osteopontin was also up-regulated in differentiated cells (Fig. 3F).
After 14 days of culture, MSCs exposed to TGF-β1 formed a compact and homogeneous pellet structure, showing an increased cellular density. Alcian blue staining revealed a homogeneous deposition of proteoglycan within the whole section of the pellet culture (Fig. 3D). The differentiated cells showed gene expression of collagen II (Fig. 3G).
Characteristics of T-cell suppression by MSCs
MSCs isolated from humans, mice, and other animal species suppress T-cell proliferation. Likewise, chicken MSCs also inhibited ConA-induced T-cell proliferation (Fig. 4A).
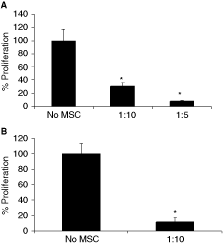
Mesenchymal stem cells (MSCs) suppress T-cell proliferation induced by ConA (
We next induced T-cell proliferation using a combination of PMA and ionomycin. PMA and ionomycin induce the differentiation and expansion of immune cells by activating the protein kinase C and inducing Ca2+ influx, respectively, bypassing the TCR signaling. PMA and ionomycin-induced proliferation was also suppressed by MSCs (Fig. 4B), suggesting that MSCs can modulate signals downstream of protein kinase C and Ca2+ influx and the T-cell receptor complex is not a target for the suppression.
T-cell suppression and NO
The exact mechanism of MSCs-induced immunosuppression is yet to be defined, but several chemokines and molecules of interest have been identified. NO produced by macrophages has been reported to suppress T-cell proliferation. In a recent study, Sato et al. [32] showed that murine MSCs T cells co-culture produce NO. This prompted us to examine the production of NO in the chicken MSCs splenocytes co-culture system. Co-culture of chicken MSCs with T cells induced a significant (P < 0.05) and dose-dependent production of NO (Fig. 5). NO production was not significant when MSCs were co-cultured with T cells in the absence of ConA or by ConA-treated MSCs or T cells. High levels of NO were produced in the co-cultures of T cells and MSCs and this was accompanied by a strong suppression of T-cell proliferation.
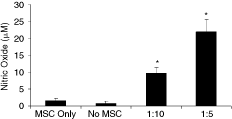
NO production by splenocytes in the presence of mesenchymal stem cells (MSCs). Splenocytes (2 × 105) were stimulated with ConA (5 µg/mL) in the presence or absence of the indicated number of MSCs for 48 h. After the incubation, the supernatant was harvested and examined for NO production by Griess assay. The values are averages ± SD of triplicate determinations. The data are representative of three separate experiments. *Significantly different from the MSC Only and No MSC group, P < 0.05.
Discussion
In the present study, we isolated and expanded adherent cells obtained from chicken bone marrow. The isolated cells showed features consistent with MSCs. The bone marrow-derived MSCs expressed transcription factors PouV, Sox2, and Nanog and differentiated along osteogenic and adipogenic lineages when cultured under the appropriate conditions. Also, the bone marrow-derived MSCs inhibited mitogen-induced proliferation of splenocytes. Therefore, we conclude that these cells isolated from chicken bone marrow are MSCs and exhibit features similar to those of mammalian MSCs.
We detected gene expression of PouV in cultured chicken MSCs at passages 2 and 8. PouV is a chicken homolog of Oct4 in mammals [30]. In mammals, Oct4 is expressed in embryonic stem cells where it inhibits tissue-specific genes and regulates self-renewal and pluripotency [33]. Recent reports have indicated the expression of Oct4 in MSCs [23,34]. Oct4 interacts with other embryonic regulators, such as Sox2 and Nanog, which are also involved in the regulation of pluripotency and inhibit differentiation [35]. Multilineage differentiation potential of stem cells is strongly correlated with the expression of Oct4. Oct4 regulates the expression of several genes expressed in stem cells, such as fibroblast growth factor 4 and transcription factors Rex1 and Sox2 [36 –39]. Expression of PouV in MSCs, like chicken embryonic stem cells may be associated with pluripotency and self-renewal of these cells.
In this study, adipogenic differentiation was induced with insulin, indomethacin, and dexamethasone, a method routinely used to stimulate the adipogenic differentiation of stem cells. The exact mechanisms of action are not fully known yet; it is believed that the insulin/IGF-1, glucocorticoids, and cAMP signaling pathways mediate the lipid accumulation in the cells [40]. Differentiation was apparent by the accumulation of lipid vacuoles within cells, which eventually filled the entire cell over a period of time. Cellular differentiation was accompanied by the expression of aP2 and PPAR-γ2. PPAR-γ2 is an adipocyte-specific transcription factor, induced early during the differentiation of adipocytes [41]. It plays an important role in adipocytes differentiation and also stabilizes the metabolic function of differentiated adipocytes [42]. aP2 is also an adipose tissue-specific gene and is expressed in the terminal stage of adipocyte differentiation [40].
The osteogenic potential of chicken MSCs was confirmed by demonstration of mineralization in the extracellular matrix produced by the differentiated cells, and by the expression of osteopontin gene. Treatment of MSCs with osteogenic induction media resulted in transformation of cells from elongated fibroblastic to shorter cuboidal cells. In primary culture, differentiated osteoblasts grew as colonies and formed mineralized nodules, a characteristic of osteogenesis in MSCs of humans [43].
Several mechanisms of T-cell suppression by MSCs have been proposed including mediation by regulatory T cells, antigen-presenting cells, and immunosuppressive cytokines [44 –46]. Recently, Sato et al. [32] showed the involvement of NO in the suppression of T cells by murine MSCs. They showed that NO suppresses T-cell proliferation by inhibiting Stat-5 phosphorylation. We also detected NO from T cells and MSCs co-cultures and the extent of T-cell suppression correlated with the amount of NO production, suggesting the important role of NO in immunomodulation. The physiologic role of NO produced in MSCs T-cell co-culture system is unknown. Speculatively, NO might be involved in the mechanisms by which MSCs in bone marrow protect hematopoietic stem cells from T-cell-mediated destruction by inhibiting T-cell proliferation.
In conclusion, we have demonstrated for the first time that MSCs can be isolated from the bone marrow of hatched chicken and these cells are capable of in vitro multiplication and multilineage differentiation, thus making them a suitable model in the field of stem cell research.
Footnotes
Author Disclosure Statement
No competing financial interests exist.