
Abstract
Electromagnetic fields (EMFs) are used to treat bone diseases. Herein, the effects of static EMFs on chondro-osteogenesis and vasculogenesis of embryonic stem (ES) cells and bone mineralization of mouse fetuses were investigated. Treatment of differentiating ES cells with static EMFs (0.4–2 mT) stimulated vasculogenesis and chondro-osteogenesis and increased reactive oxygen species (ROS), which was abolished by the free radical scavengers trolox, 1,10-phenanthroline (phen), and the NAD(P)H oxidase inhibitor diphenylen iodonium (DPI). In contrast, EMFs of 10 mT field strength exerted inhibitory effects on vasculogenesis and chondro-osteogenesis despite robust ROS generation. EMFs of 1 mT and 10 mT increased and decreased vascular endothelial growth factor (VEGF) expression, respectively, which was abolished by DPI and radical scavengers. EMFs activated extracellular-regulated kinase 1/2 (ERK1/2), p38, and c-jun N-terminal kinase (JNK), which was sensitive to DPI treatment. The increase in VEGF by EMFs was inhibited by the ERK1/2 inhibitor U0126 but not by SB203580 and SP600125, which are p38 and JNK inhibitors, respectively, suggesting VEGF regulation by ERK1/2. Chondro-osteogenesis and vasculogenesis of ES cells was blunted by trolox, DPI, and the VEGF receptor-2 (flk-1) antagonist SU5614. In mouse fetuses 1 mT EMFs increased and 10 mT EMFs decreased bone mineralization, which was abolished in the presence of trolox. Hence, EMFs induced chondro-osteogenesis and vasculogenesis in ES cells and bone mineralization of mouse fetuses by a ROS-dependent up-regulation of VEGF expression.
Introduction
A
To understand the biological mechanisms of EMF exposure, not only epidemiological and animal studies but also in vitro investigations on cell culture models are required. Differentiating embryonic stem (ES) cells represent in vitro models for early postimplantation embryos and are ideally suitable to analyze signaling pathways initiated in differentiating tissues upon exposure to EMFs. Significant efforts are currently undertaken to understand signaling pathways that regulate cell type-specific differentiation of ES cells and may be utilized in the future to support cell transplantation approaches. Recently, several studies have shown that ES cells differentiate spontaneously into chondro-osteocytes; however, no insight into underlying molecular mechanisms was given [6 –10]. It is generally accepted that bone formation is dependent on angiogenesis that is blood vessel growth. Both the two decisive processes of development and formation of the skeleton, that is, intramembranous and endochondral bone ossification, occur in close proximity to growing blood vessels [11]. Recent studies have shown that vascular endothelial growth factor (VEGF) may act as an essential mediator of osteogenesis. During osteogenesis VEGF exerts multiple functions, not only in bone angiogenesis, but also in different aspects of bone development, including chondrocyte differentiation, osteoblast differentiation, and osteoclast recruitment [12,13]. VEGF is known to be an angiogenic factor in vivo and in vitro, a mitogen for endothelial cells, and exerts effects on vascular permeability [14]. Only in the last few years attention has been focused on VEGF synthesis and VEGF receptor expression in nonendothelial tissues [15]. As far as the chondro-osteogenic lineage is concerned, VEGF was described as a binding but not proliferative agent for cultured osteoblasts, inducing migration, alkaline phosphatase increase, and parathyroid hormone-dependent cyclic adenosine monophosphate (cAMP) accumulation, thus suggesting a possible role in osteoblast differentiation [16]. The essential role of VEGF in endochondral bone development was assessed by inactivating VEGF through systemic administration of soluble chimeric protein to mice [17]. It was concluded that VEGF-mediated vascularization is an essential signal to regulate growth and bone morphogenesis through coupling resorption of cartilage with bone formation. The important role of VEGF in cartilage vascularization and bone development has been supported by several authors [18 –20]. Several mechanisms for VEGF controlling osteoblast activity have been proposed. First, VEGF could couple angiogenesis and osteogenesis by manipulating the angiogenic response to osteoblastic activity. Second, VEGF could act as an autocrine regulator of osteoblastic differentiation and activity. Third, by expressing VEGF, osteoblasts could induce cells in the vicinity to express factors that regulate osteoblastic activity [13].
The effects of EMFs on chondro-osteogenesis of ES cells and the role of angiogenesis in this process are so far not investigated. Previously it has been shown that EMFs induce angiogenesis in endothelial cells [21], which may be related to fibroblast growth factor-2 (FGF-2) release as shown in vitro and in vivo [22]. Furthermore it has been demonstrated that EMFs give rise to the generation of reactive oxygen species (ROS) [23], which may stimulate VEGF signaling pathways as well as VEGF expression and/or may activate mitogen-activated protein kinase (MAPK) pathways that regulate VEGF signaling. In the present study chondro-osteogenesis was studied upon exposure toward static magnetic fields. It is demonstrated that EMF-induced chondro-osteogenesis is dependent on ROS generation, which stimulates VEGF expression through the activation of MAPK pathways.
Materials and Methods
ES cell culture
The ES cell line CCE was grown on mitomycin C (Sigma, Taufkirchen, Germany)-inactivated feeder layers of primary murine embryonic fibroblasts for a maximum of eight passages in Iscove's medium (Gibco, Life Technologies, Gaithersburg, Maryland) supplemented with 20% heat-inactivated fetal calf serum (FCS; Boehringer-Mannheim, Mannheim, Germany), 2 mM Glutamax (Gibco), 100 μM β-mercaptoethanol (Sigma, Deisenhofen, Germany), 1% MEM nonessential amino acids 100×; stock solution (Gibco), 1,000 U/mL leukemia inhibitory factor (LIF) (Sigma), 100 IU/mL penicillin, and 100 μg/mL streptomycin (both Gibco). At day 0 of differentiation, adherent cells were enzymatically dissociated using 0.2% trypsin/0.05% EDTA in PBS (Gibco) and ES cell differentiation was started by using the hanging drop method (400 cells/20 μL drops prepared in 20% complete Iscove's medium without LIF). After 48 h embryoid bodies were collected and further differentiated in 20% complete Iscove's medium without LIF in suspension cultures in bacteriological Petri dishes. For certain experiments embryoid bodies were cultured in suspension cultures for additional 4 days and then plated onto tissue culture dishes to grow in adherent conditions till day 20. Cell culture plates were placed in the center of the exposure system situated in a 37°C humidified incubator containing 5% CO2 or in a control incubator for the exposure time period according to the experimental conditions.
EMF exposure system
The EMF exposure system was manufactured to provide a uniform EMF distribution in a long cylindrical coil when current passed. The coils were wound on tube from PVC, which completely shielded against emission of electric fields. The exposure system was calibrated in order to obtain the uniform EMF continuously within the exposure time. A variable FH 51 Gauss-/Teslameter (MAGNET-PHYSIK, model no. 2000510, Cologne, Germany) with a small probe 761 was used to measure the strength of EMF. Three-day-old embryoid bodies were exposed to static EMFs at different field strengths ranging from 0.4 to 10 mT, 8 h per day for 17 days.
Mouse embryos
Pure strain BALB/c mice were housed at approximately 23 ± 2°C and 50 ± 10% humidity with a 12:12 h light–dark cycle, and with access to standard chow and water ad libidum. Animals were allowed to mate overnight and the females were examined the following morning for vaginal plug and the vaginal smear carried out. This day was assigned as the day 0 of gestation [24]. For the current study six pregnant groups of mice were used. The first two groups functioned as a control, one was injected intraperitoneally (i.p.) once daily with trolox 50 mg/kg [25]. All other groups were exposed to static EMFs at either 1 mT or 10 mT for 8 h/day from third day of gestation till day 20. Two of these groups were injected with trolox as described. Dams were killed on day 20 and the embryos were fixed in 95% ethanol for skeletal staining. Each treatment group included 10 animals.
Measurement of ROS generation
Intracellular ROS levels were measured using the fluorescent dye 2,7, dichlorodihydrofluorescein diacetate H2DCF-DA (Molecular Probes Eugene, OR), which is a nonpolar compound that is converted into a non-fluorescent polar derivative (H2DCF) by cellular esterases after incorporation into cells. H2DCF is membrane impermeable and is rapidly oxidized to the highly fluorescent DCF in the presence of intracellular ROS. For the experiments, embryoid bodies in suspension were incubated in E1 buffer (containing in mM: NaCl 135, KCl 5.4, CaCl2 1.8, MgCl2 1, glucose 10, HEPES 10 (pH 7.4 at 23°C), and 10 μM H2DCF-DA dissolved in DMSO was added. After 20 min, the embryoid bodies were rinsed three times in E1 buffer and DCF fluorescence was recorded by laser scanning confocal microscope (C-LSM 410, Zeiss, Jena, Germany) connected to an inverted microscope (Axiovert 135, Zeiss).
Immunohistochemistry
For immunofluorescence stainings, differentiating embryoid bodies were fixed for 1 h at 4°C in 4% formaldehyde in PBS. For immunofluorescence and western blotting monoclonal mouse anti-VEGF (Biogenesis, Poole, UK), monoclonal mouse anti-bone gamma-carboxyglutamate protein (BGLAP) (Biogenesis, Poole, UK), monoclonal mouse anti-phospho-ERK (pERK), phospho-p38 MAPK (p-p38), and phospho-JNK (pp54 and pp46) (all from Cell Signalling Technology, Frankfurt, Germany) antibodies were used.
Alcian blue and Alizarin red S staining of skeletons of mouse embryos
Alizarin red S was used as a biochemical assay to determine osteoblast differentiation and the presence of calcific deposition by cells of an osteogenic lineage. Cartilage positive for type II collagen, which is the principal collagenous component of cartilage, stains strongly with Alcian blue. The embryos were washed with PBS and fixed with 95% ethanol (4°C, for 24 h). Cartilage and bones in whole mouse embryos (20 days) were visualized by light microscopy after staining with Alcian blue and Alizarin red S (Sigma) and clarification of soft tissue with potassium hydroxide [26,27].
Treatment with inhibitors
Embryoid bodies were incubated during the time of EMF treatment with the specific p38 MAPK, c-jun N-terminal kinase (JNK), and extracellular-regulated kinase 1/2 (ERK1/2) inhibitors SB203580 (1 μM), SP600125 (1 μM), and U0126 (10 μM), respectively (Calbiochem, Bad Soden, Germany); with the ROS scavengers trolox (100 μM) (Calbiochem), and 1,10-phenanthroline (10 μM), or with the NAD(P)H oxidase inhibitor diphenylene iodonium (DPI) chloride (10 μM) (Sigma, Deisenhofen, Germany). For inhibition of VEGF receptor signaling SU5614 (1 μM) (Calbiochem, Bad Soden, Germany) was used.
Reverse transcription-polymerase chain reaction (RT-PCR)
RNA isolation from embryoid bodies was performed using Trizol reagent (Invitrogen, Karlsruhe, Germany). cDNA was synthesized from 2 μg total RNA using a Superscript II reverse transcriptase (Invitrogen) according to manufacturer's instructions and using random primer. The RT product was diluted 1:10 and PCR was performed with following primers (Sigma, Geonysis, Germany): VEGF splice variants, 5′-TCC-ACC-ATG-CCA-AGT-GGT-3′ (sense) and 5′-TCG-GGG-TAC-TCC-TGG-AAG-AT-3′ (antisense). β-Actin served as the internal control with the specific primer pair 5′-GAT-GAC-CCA-GAT-CAT-GTT-TGA-G-3′ (sense) and 5′-CCA-TCA-CAA-TGC-CTG-TGG-TA-3′ (antisense) with 40 cycles performed at 58°C annealing temperature. Gel images were subsequently captured using a 1% agarose gel.
Western blot assay
The western blot assays were carried out after washing the embryoid bodies in PBS and lysed in lysis buffer (20 mM Tris–HCl [pH 7.5], 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1% Triton X-100, 2.5 mM sodium pyrophosphate, 1 mM β-glycerophosphate, and 1 mM Na3VO4) that contained 1 mM phenylmethylsulfonyl fluoride (a protease inhibitor) for 30 min on ice. Samples were centrifuged at 13,000g for 10 min to pellet the debris. After determination of the protein concentration using a Bio-Rad protein assay (Bio-Rad, Munich, Germany), 20 μg of protein samples was boiled, separated in 12% SDS polyacrylamide gels, and transferred to nitrocellulose membranes by electroblotting at 80 V for 3 h. Membranes were blocked with 5% (wt/vol) dry fat-free milk powder in Tris-buffered saline with 0.1% Tween (TBST) for 60 min at room temperature. Incubation with primary antibodies was performed at 4°C overnight. After washing with TBST, the membrane was incubated with a horseradish peroxidase (HRP)-conjugated secondary antibody (Cell Signaling Technology, Frankfurt, Germany) for 60 min at room temperature. The blot was developed using the ECL detection kit (Amersham, Freiburg, Germany) to produce a chemiluminescence signal.
Statistical analysis
Data were given as mean values ± SD, with n indicating the number of experiments. In each experiment at least 15 embryoid bodies were analyzed. GraphPad InStat-3 software (GraphPad Software Inc., San Diego, CA) was applied for t-test unpaired data analysis. A value of P < 0.05 was considered significant (*). Data are expressed as percentage of expression relative to control values, which were set to 100%.
Results
Stimulation of vasculogenesis and chondro-osteogenesis in ES cells by static EMFs
The present study was undertaken to investigate the impact of static EMFs on chondro-osteogenesis of ES cells and the role of vasculogenesis in this process. Therefore differentiating 3-day-old embryoid bodies were exposed to static EMFs at different field strengths ranging from 0.4 to 10 mT, 8 h per day for 17 days. Subsequently the platelet endothelial cell adhesion molecule-1 (PECAM-1) (CD31)-positive cell areas (Fig. 1A, n = 3), the number of embryoid bodies displaying cartilage nodules (Fig. 1B, n = 3), the number of cartilage nodules per embryoid body (Fig. 1C, n = 3), the cartilage nodule area (Fig. 1D, n = 3), and the expression of BGLAP, which is indicative of osteogenic differentiation, were assessed (Fig. 1E, n = 3). Apparently treatment of embryoid bodies with static EMFs significantly and dose-dependently increased vasculogenesis as well as chondro-osteogenesis in embryoid bodies with maximum effects between 1 and 1.6 mT. Interestingly, a field strength of 10 mT resulted in adverse effects, although no obvious cytotoxicity was observed at 10 mT field strength (data not shown).
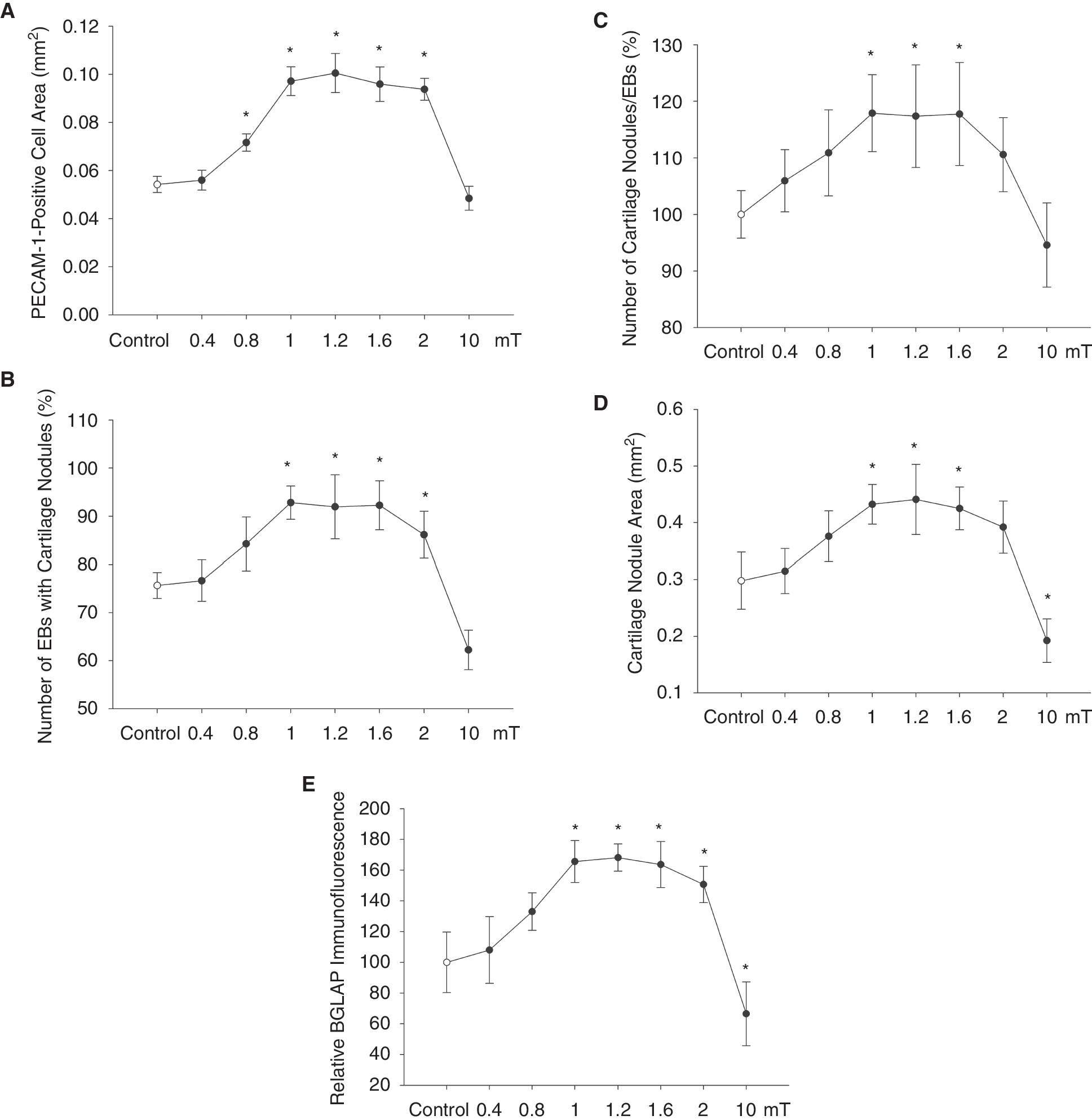
Stimulation of vasculogenesis and chondro-osteogenesis by static electromagnetic fields (EMFs). Three-day-old embryoid bodies were treated for 8 h/day until day 20 of cell culture with 0.4, 0.8, 1, 1.2, 1.6, 2, and 10 mT EMFs, respectively. Subsequently, cell areas positive for PECAM-1 (CD31) (
Static EMFs induce ROS generation in embryoid bodies
Previously it has been shown that ROS are generated following EMF treatment in several preparations [23,28]. Recent studies from our group have shown that ROS are involved in vasculogenesis [29] as well as cardiomyogenesis [30,31] of ES cells. To investigate whether static EMFs would increase ROS generation 5-day-old embryoid bodies grown from ES cells were treated with a magnetic field strength of 1 and 10 mT and intracellular generation of ROS was investigated using the fluorescent ROS indicator H2DCF-DA. It was obvious that static EMFs dose-dependently increased the production of endogenous ROS in embryoid bodies (Fig. 2A, n = 4). The strongest increase in the oxidation kinetics of H2DCF was observed in embryoid bodies exposed to a 10 mT EMF for 8 h/day. Exposure of embryoid bodies to 1 mT likewise showed significantly increased ROS generation, but to a lower extent as compared to exposure to 10 mT. The generation of ROS was inhibited by the NAD(P)H oxidase inhibitor DPI (10 μM) as well as the ROS scavengers 1,10-phenanthroline (10 μM) and trolox (100 μM) (Fig. 2B, n = 4), indicating that ROS may be generated through Nox enzymes.
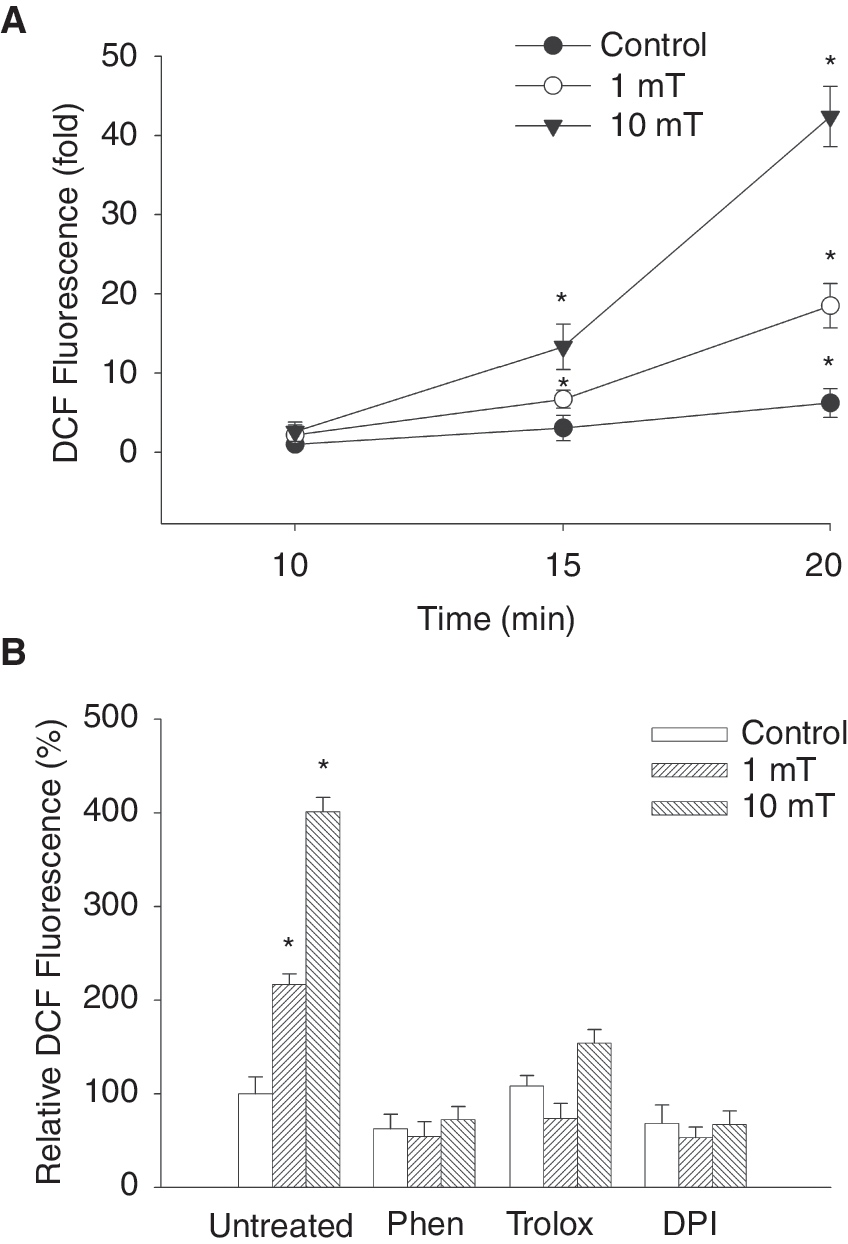
Generation of reactive oxygen species (ROS) upon treatment of embryonic stem (ES) cell-derived embryoid bodies with 1 mT and 10 mT electromagnetic fields (EMFs). (
Static EMFs increase VEGF in a ROS-dependent manner
ROS are known to regulate the expression of the VEGF gene as well as to be involved in VEGF-R signaling. To investigate whether the EMFs applied in the present study would change VEGF levels and whether this process would be dependent on ROS generation VEGF expression was assessed by immunohistochemistry as well as western blot. This was done after treatment of 5-day-old embryoid bodies for 8 h with EMFs of 1 and 10 mT (Fig. 3A and B, n = 4). Interestingly exposure to a low EMF of 1 mT field strength significantly increased VEGF expression, whereas a decrease was observed with 10 mT. To determine whether the increase in VEGF expression upon treatment with 1 mT EMFs was due to ROS generation, embryoid bodies were incubated with either the NAD(P)H oxidase inhibitor DPI (10 μM) or the free radical scavenger trolox (100 μM) during treatment with EMFs. As shown in Figure 3A, B preincubation of embryoid bodies with DPI (10 μM) resulted in significant inhibition of VEGF under the exposure to 1 mT whereas a partial recovery of VEGF expression toward the untreated control was observed at 10 mT. Comparable results were achieved upon treatment with free radical scavenger. Embryoid bodies preincubated with trolox (100 μM) showed significant inhibition of VEGF expression at 1 mT, whereas significant increase in VEGF expression was observed upon 10 mT exposure (Fig. 3A and B). From these experiments it was concluded that VEGF upon EMF treatment is regulated by ROS generation presumably through NAD(P)H oxidase activity.
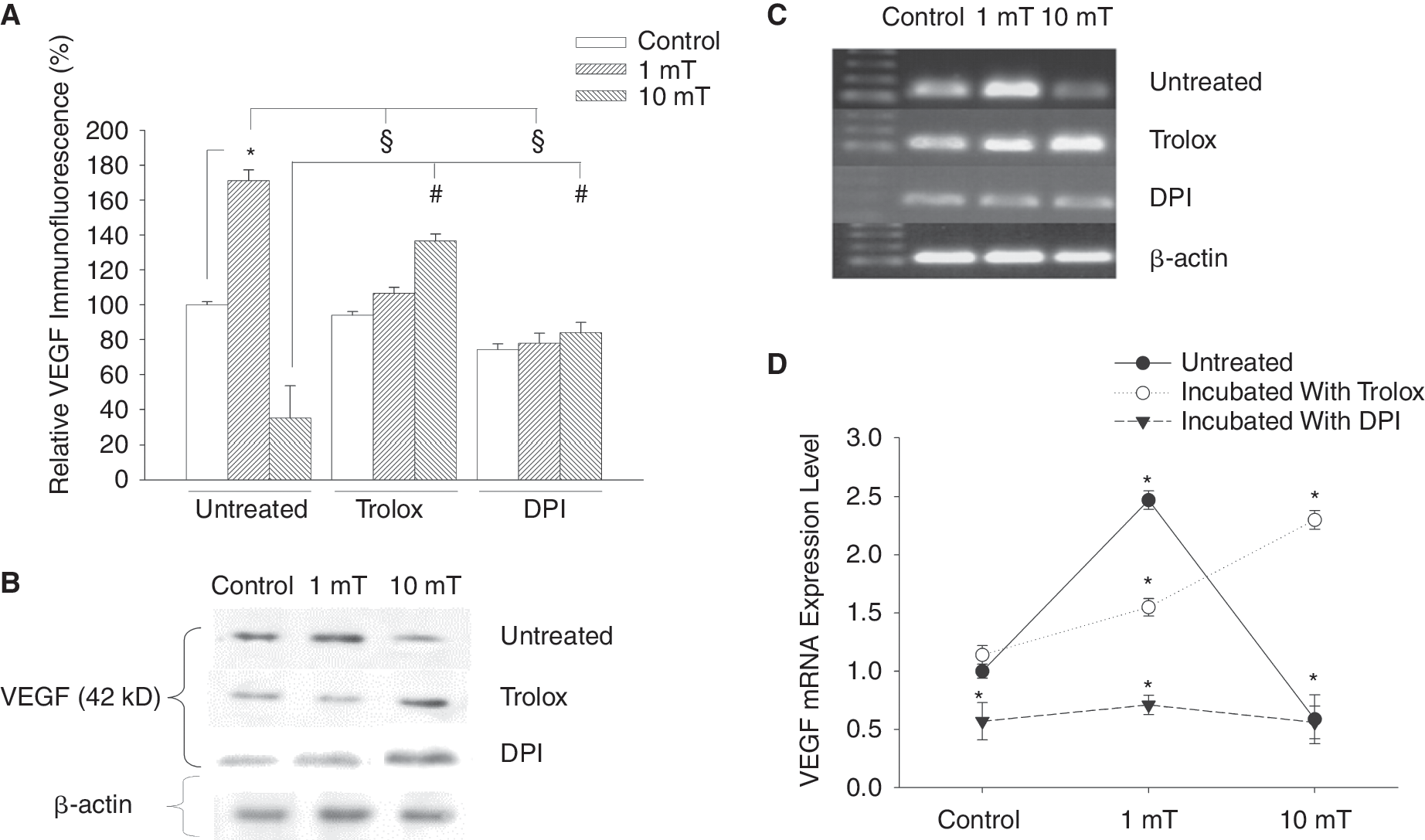
Stimulation of vascular endothelial growth factor (VEGF) protein and mRNA expression in embryonic stem (ES) cell-derived embryoid bodies upon treatment with electromagnetic fields (EMFs). Five-day-old embryoid bodies were treated for 8 h/day with either 1 or 10 mT EMFs and were analyzed after 24 h for VEGF expression in immunohistochemical analyses (
To examine the changes in VEGF gene expression in the process of chondrogenesis under static EMFs, RNA was isolated from 20-day-old embryoid bodies and subjected to RT-PCR analysis using specific primers designed to amplify VEGF. PCR analysis showed a high rate of VEGF expression after exposure to 1 mT EMFs. In contrast less expression of VEGF was found at exposure to 10 mT EMFs (Fig. 3C and D, n = 4). Treatment of embryoid bodies with trolox (100 μM) or DPI (10 μM) resulted in responses similar to those obtained by immunohistochemistry and western blot, which clearly indicates that ROS modulate the level of VEGF expression.
Regulation of VEGF expression upon static EMF treatment by ROS and MAPK pathways
To elucidate the signaling pathways involved in the stimulation of VEGF expression upon treatment of ES cells with static EMFs, the activation of ERK1/2, p38, as well as JNK signaling pathways was determined by using phospho-specific antibodies. It was found that exposure to 1 mT EMFs resulted in a significant activation of ERK1/2, p38, as well as JNK in 5-day-old embryoid bodies after 8 h of EMF exposure (Fig. 4A and B, n = 3). The activation of different MAPK members was not significantly different when embryoid bodies were treated with either 1 or 10 mT (see Fig. 4A and B). The effect of EMF treatment on MAPK activation was totally abolished in the presence of the NAD(P)H oxidase inhibitor DPI (10 μM), indicating an involvement of ROS and NAD(P)H oxidase activity in MAPK activation upon EMF treatment.

Activation of the MAPKs ERK1/2, p38, and JNK upon treatment with electromagnetic fields (EMFs) in embryonic stem (ES) cell-derived embryoid bodies. Immunohistochemical analysis (
To determine the specific MAPK crucial for VEGF expression upon treatment of embryoid bodies with EMFs, we investigated VEGF expression following EMF exposure in the presence of the ERK1/2 inhibitor U0126 (10 μM), the p38 inhibitor SB203580 (1 μM), or the JNK inhibitor SP600125 (1 μM). It was apparent that ERK1/2 inhibition by U0126 reduced VEGF expression in the control, as well as in the sample exposed to a 1 mT EMF. In contrast, no significant change in VEGF expression was detected by immunofluorescence and western blotting in the presence of SB203580 or SP600125 upon EMF exposure, excluding involvement of the JNK or p38 MAPK signaling cascade in the regulation of VEGF expression in embryoid bodies (Fig. 5A and B, n = 5).
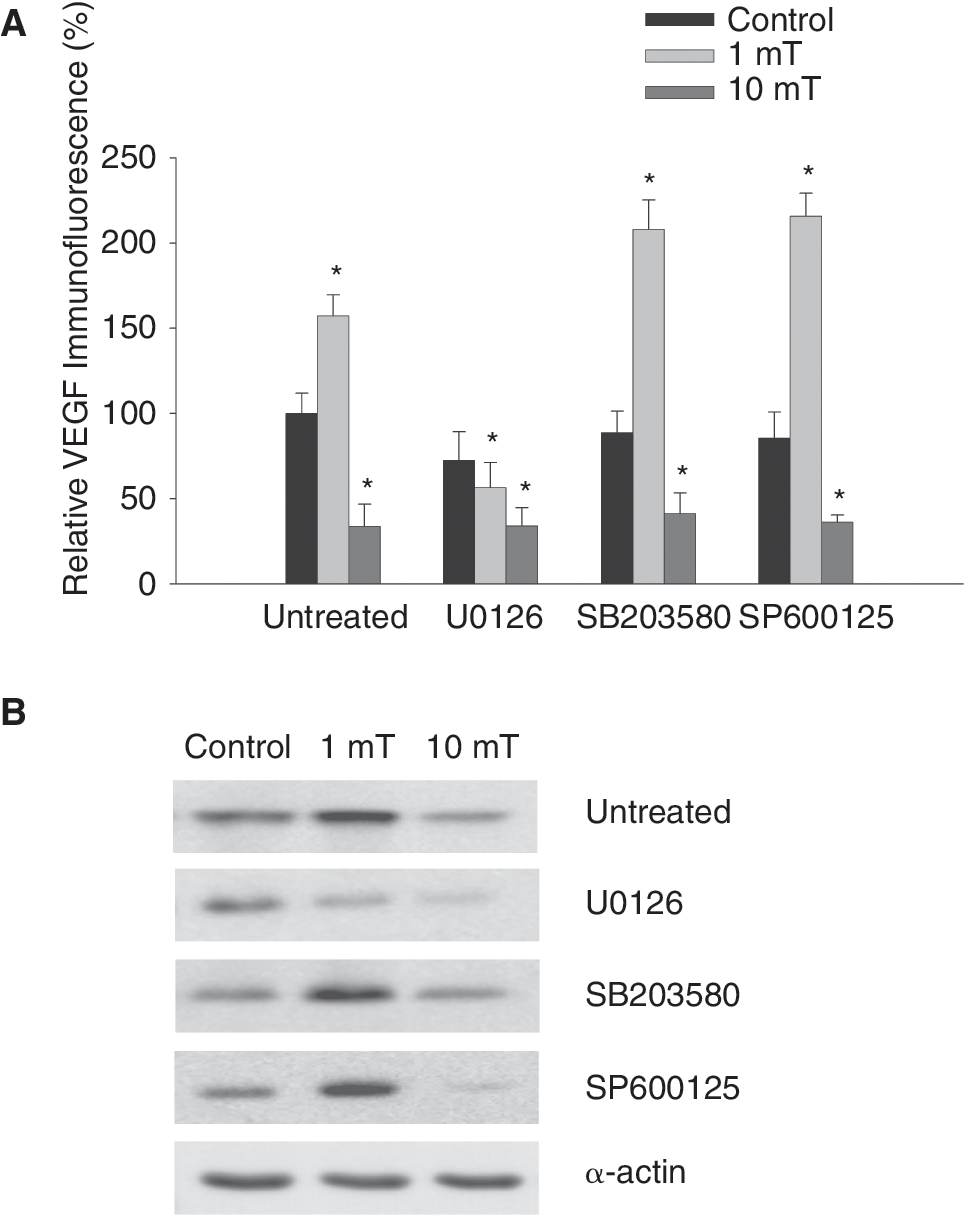
Effects of MAPK inhibitors on electromagnetic fields (EMFs)-induced vascular endothelial growth factor (VEGF) expression. (
Role of ROS and VEGF in chondro-osteogenic differentiation of ES cells
The goal of the present study was to investigate whether EMFs would stimulate chondro-osteogenesis dependent on vasculogenesis and generation of ROS. To investigate whether ROS are involved in chondro-osteogenesis processes, embryoid bodies were incubated with either DPI (10 μM) or trolox (100 μM) during the exposure to static EMFs (1, 10 μT). Following 20 days of cell culture (17 days exposure to EMFs), the number of plated embryoid bodies showing Alcian blue-positive cartilage nodules and the number of nodules/plated embryoid body were counted. In parallel the areas of nodules were measured. To correlate chondrogenesis to osteogenesis and vasculogenesis, embryoid bodies were stained with antibodies for endothelial cells (PECAM-1) and osteoblast cells (BGLAP). It was apparent that an EMF strength of 1 mT significantly raised numbers of embryoid bodies differentiating to chondrocyte nodules and increased the numbers of chondrocyte nodules per embyoid body as well as the nodule areas (Fig. 6A–C, n = 6). Moreover, an increase in capillary structures indicated by PECAM-1 expression (Fig. 7A and B, n = 4) and also osteocyte progenitor cells indicated by BGLAP expression (Fig. 8A and B, n = 4) was observed in embryoid bodies treated with 1 mT EMFs. Interestingly, an enhanced invasion of capillary-like structures into the hypertrophic zone of the cartilage nodules was observed (see Fig. 9B), as compared to the untreated control (see Fig. 9A), supporting the notion of a correlation between vasculogenesis and chondro-osteogenic differentiation. In contrast, 10 mT static EMFs showed a significant decrease in the number of embryoid bodies differentiating to cartilage nodules as well as the number of nodules per embryoid body and the nodule area (Fig. 6A–C). Moreover, significant decrease in endothelial cell (Fig. 7A and B) and osteoblast differentiation (Fig. 8A and B) accompanied by a complete inhibition of blood vessel invasion into the hypertrophic zone of cartilage nodules and decrease in osteoblast differentiation in nodules (see Fig. 9C) was found at 10 mT EMF exposure. These results suggest that angiogenesis adjacent to the cartilage nodules may be a prerequisite for further differentiation into bone.
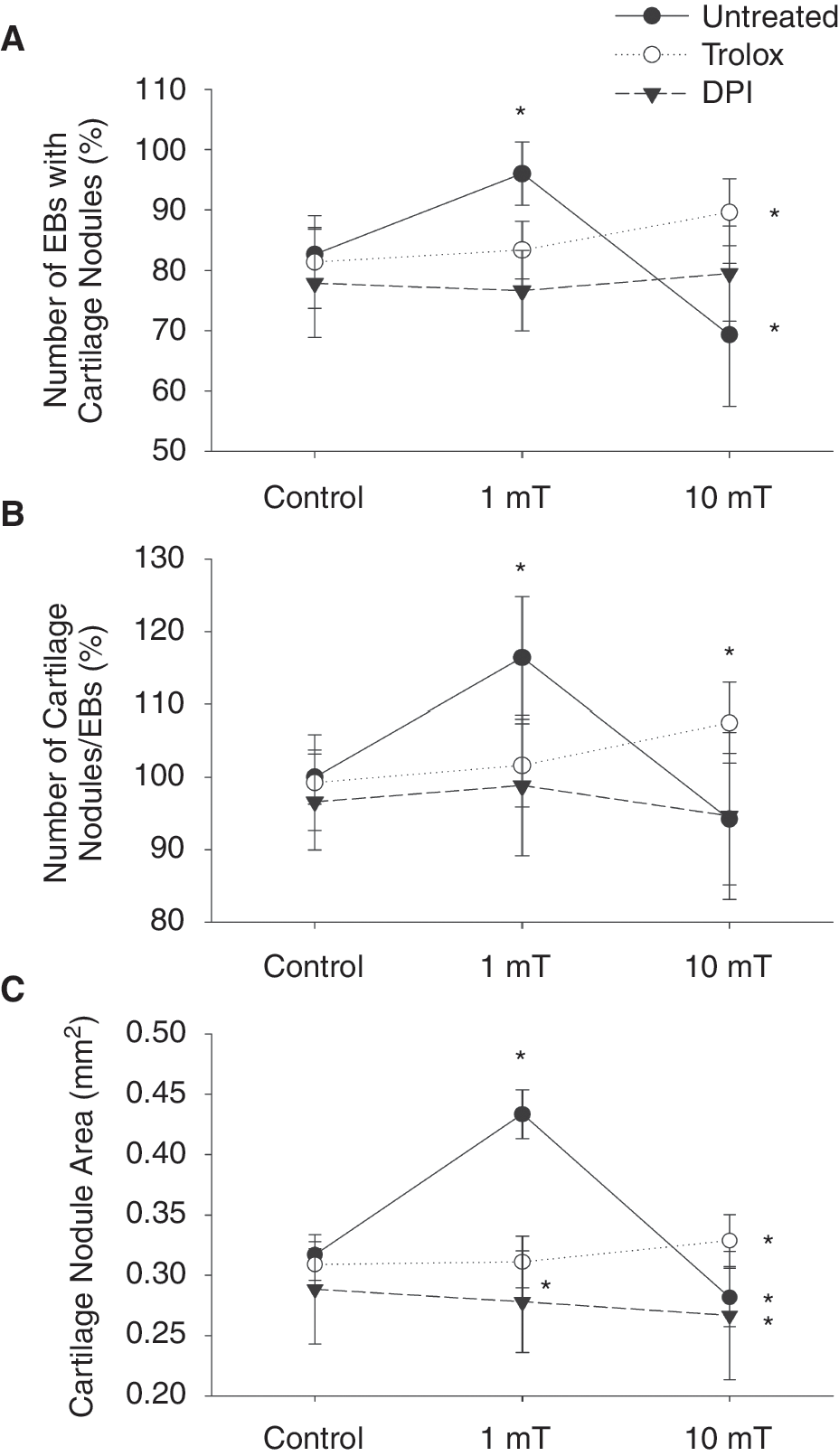
Stimulation of chondro-osteogenic differentiation after application of electromagnetic fields (EMFs) to embryonic stem (ES) cell-derived embryoid bodies (EBs). Embryoid bodies were treated on day 3 for 8 h/day with either 1 or 10 mT EMFs. On day 20 of cell culture, the number of embryoid bodies differentiating cartilage nodules (

Stimulation of vasculogenesis of embryonic stem (ES) cell-derived embryoid bodies by electromagnetic fields (EMFs). (

Stimulation of osteoblast differentiation of embryonic stem (ES) cell-derived embryoid bodies by electromagnetic fields (EMFs). (

Vascularization of chondro-osteogenic nodules in embryonic stem (ES) cell-derived embryoid bodies upon treatment with electromagnetic fields (EMFs). Nodules showing osteogenic differentiation as indicated by immunohistochemical staining against the osteoblast marker BGLAP (green) were analyzed for endothelial cell invasion as indicated by PECAM-1 staining (red); blue: transmission image overlay. Treatment with 1 mT EMFs resulted in increased BGLAP expression and endothelial cell invasion into the hypertrophic zone of nodules (
The role of ROS in chondro-osteogenesis and vasculogenesis upon treatment of embryoid bodies with EMFs was investigated following incubation of embryoid bodies with either trolox (100 μM) or DPI (10 μM) during EMF exposure. As shown in Figure 6A–C, pretreatment with both trolox (100 μM) and DPI (10 μM) resulted in complete inhibition of chondro-osteogenesis upon exposure to 1 mT EMFs as evaluated by determination of the number of embryoid bodies displaying Alcian blue-positive cartilage nodules, the number of nodules/embryoid body, and the area of these nodules (Fig. 6A–C). Moreover, we noticed significant decrease in the areas covered by capillary-like structures stained by PECAM-1 (Fig. 7A, B and Fig. 9D, E) and BGLAP expression (Fig. 8A and B), indicating a decrease of osteoblast differentiation in nodules. In contrast, the number of embryoid bodies differentiating to cartilage nodules, the number of nodules per embryoid body, and area of nodules were significantly increased, when embryoid bodies were exposed to the 10 mT EMFs under conditions of co-treatment with trolox (100 μM). Furthermore, under high-field (10 mT) exposure and trolox treatment a significant increase in blood vessel invasion into the hypertrophic zone of nodules and an increase of BGLAP expression were observed (Fig. 9F). In contrast embryoid bodies preincubated with DPI (10 μM) and exposed to either 1 or 10 mT EMFs showed a significant decrease in the number of embryoid bodies displaying cartilage nodules, the number of nodules per embryoid body, and the area of nodules (Fig. 6A–C). Moreover, under these experimental conditions inhibition of blood vessel differentiation and endothelial cell invasion into nodules (Fig. 7A, B and Fig. 9G–I) as well as inhibition of osteoblast differentiation (Fig. 8A, B and Fig. 9G–I) was observed.
Chondrocyte differentiation may be influenced by paracrine mechanisms, for example, by growth factors released to the cell culture medium during exposure of ES cells with EMFs. To exclude the influence of paracrine mechanisms differentiating embryoid bodies were treated in the absence of an external field with conditioned cell culture medium obtained from cells treated with 1 mT EMFs. Under these experimental conditions no difference in the number of embryoid bodies differentiating cartilage nodules, the number of cartilage nodules per embryoid body, and the cartilage nodule area was observed (data not shown). These data indicate that chondrocyte differentiation was not influenced by factors released to the cell culture medium upon EMF treatment. However, our data do not exclude that unstable mediators only active over a short distance could account for the observed effects.
Inhibition of chondro-osteogenic differentiation of ES cells upon interference with VEGF-R2 signaling
It is well established that VEGF elicits downstream signaling pathways via binding and activation to VEGF-R2 (flk-1). To further substantiate our observations of a relationship between vasculogenesis and chondro-osteogenesis embryoid bodies were treated with the VEGF-R2 inhibitor SU5614 (1 μM) during the time of exposure with EMFs. Preincubation of embryoid bodies with SU5614 inhibited angiogenesis (Fig. 7A and B) as well as osteogenesis (Fig. 8A and B) in the control samples as well as in the samples treated with EMFs. The number and size of cartilage nodules were not affected (data not shown). Moreover, upon treatment with SU5614 a significant inhibition of blood vessel invasion into cartilage nodules accompanied by a decrease in osteoblast differentiation (Fig. 9J–L) was observed, thus suggesting that VEGF induces bone development in differentiating ES cells through induction of blood vessel formation and osteoblasts differentiation.
Effects of static EMFs on chondro-osteogenesis and bone mineralization of mouse embryos
To confirm our in vitro results, we analyzed the role of EMFs on chondro-osteogenesis in vivo by treating pregnant mice from the third day of gestation till day 20 either with 1 mT or 10 mT EMFs for 8 h/day (Fig. 10, n = 10). Analysis of 20th day skeletons of mouse embryos showed that the bone mineral density zones were increased upon treatment with 1 mT EMFs (Fig. 10B) but they were strongly reduced following 10 mT EMF treatment (Fig. 10C) as compared to the untreated control (Fig. 10A), indicating a reduction in osteoblast differentiation. Not only limb bones were affected in the exposed mouse embryos; retardation/improvement in mineralization zones could also be observed in the skull components, for example, in tympanic, nasal, squamosal, parietal, interparietal, and exocciptal regions. To evaluate the role of ROS during EMF treatment pregnant mice were daily i.p. injected with trolox during the treatment with EMFs. This treatment resulted in reduced bone mineralization in untreated controls (Fig. 9D) and mouse embryos treated with 1 mT EMFs (Fig. 10E). In contrast, there was apparent improvement in embryonic bone mineralization in embryos of mothers i.p. injected with trolox and in parallel exposed to 10 mT SMF during the pregnancy (Fig. 10F) as compared embryos of mice treated with 10 mT EMFs in the absence of trolox (see Fig. 10C). This increase in bone mineralization was observed in long bones in the limbs, in ribs, in sternum, and in the bones of the vertebral column. Our data therefore suggest that ROS are important for the process of endochondral bone formation and bone mineral density not only in the in vitro system of ES cell-derived embryoid bodies but also in vivo in mouse embryos.
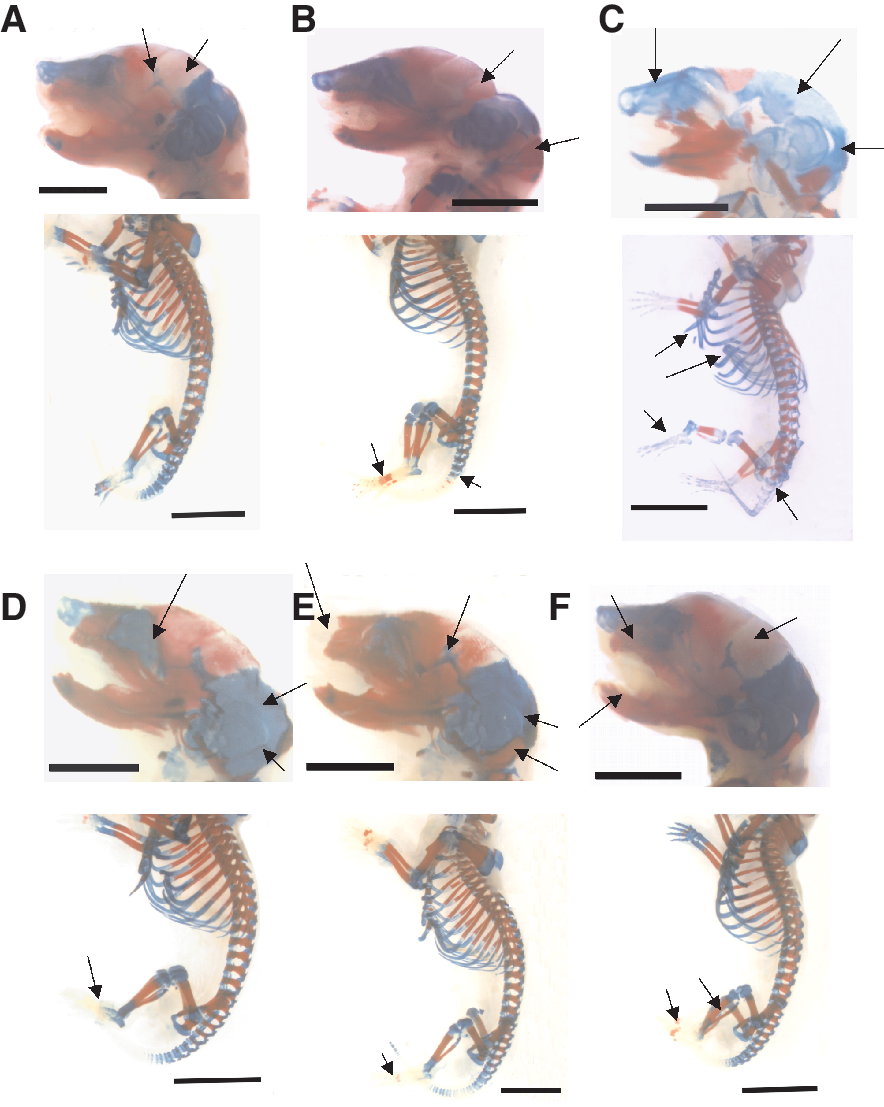
Bone mineralization of skeletons of mouse embryos treated with electromagnetic fields (EMFs). Pregnant mice remained either untreated (
Discussion
ES cells have the unlimited capacity to self-renew and maintain their pluripotency in culture. Upon induction of various signals they differentiate into distinctive cell types such as neurons, cardiomyocytes, endothelial cells, chondrocytes, and osteoblasts [32]. Our results indicate that ROS stimulated by static EMFs play an essential role as signal transducers for chondrocyte and osteocyte differentiation from ES cells through induction of hypertrophic chondrocytes. In pilot studies we furthermore investigated whether 1 mT and 10 mT 50 Hz alternating current (AC) fields would increase ROS and enhance chondro-osteogenesis of ES cells. These experiments revealed that AC fields increased ROS in differentiating embryoid bodies comparably to direct current (DC) fields. Likewise 1 mT AC fields stimulated chondro-osteogenesis, whereas an inhibitory effect was observed at 10 mT field strength (data not shown). It is well known that chondrocyte hypertrophy is associated with apoptosis, angiogenesis, and mineralization. These processes may be regulated by intracellular ROS levels acting within different signal transduction pathways. Hence we assumed that EMF-induced ROS may regulate the interaction between vasculogenesis and chondro-osteogenesis during the differentiation of ES cells.
Comparing the skeletal development of mice embryos in vivo with chondro-osteocyte differentiation in ES cells in vitro revealed a broad similarity in chondro-osteogenic differentiation. In this study we have provided in vivo evidence that ROS are required not only for chondro-osteogenesis of ES cells but likewise for embryonic chondro-osteogenesis and bone mineral density in developing bones in vivo. At day 20, mouse embryos treated with trolox showed a decrease in the hypertrophic cartilage and a decrease in bone mineral density as compared with untreated mice. Moreover, trolox scavenged the excess of ROS generated from NAD(P)H oxidase if embryos were exposed to 10 mT. This led to improvement of chondro-osteocyte differentiation and endochondral ossification and antagonized the detrimental effects of EMFs on chondro-osteogenesis at high EMF strength. These data clearly suggest that ROS are critical for chondrocyte and bone formation in vivo.
The actual mechanisms of EMF action on bone development are not known, but effects on signal transduction at the receptor and at the intracellular level have been suggested. The effect of EMF was hypothesized to be propagated and amplified by signal transduction pathways, ultimately leading to modification of cell behavior [33 –35]. EMFs may interact with ion channels or voltage-dependent receptors. Furthermore, EMFs may interact within signal transduction pathways; for example, ligand (hormone or growth factor) binding, conformational changes of receptors, membrane fluidity, G-protein kinetics, its affector enzyme activity (adenylate cyclase), and phosphorylation/dephosphorylation of receptors, enzymes, or proteins [34]. All these interaction sites of EMFs may utilize ROS in their specific molecular function. In ES cells significant amounts of ROS are generated by an NAD(P)H oxidase that has been demonstrated in the present study to be activated by static EMFs. ROS generation following EMF exposure has previously been reported in differentiating ES cells [29,31,36], in human monocytes and lymphocytes [37 –39], in experimental animals [40], and in oral mucosa cancer cells [41].
Members of the MAPK family, ERK1/2, JNK, and p38, are central elements that transduce the signal generated by growth factors, cytokines, and stress [42]. Our findings show that static EMFs elevated intracellular ROS levels, which in turn activated the ERK1/2 pathway. ROS were apparently generated through the activation of NAD(P)H oxidase, since preincubation with the NAD(P)H oxidase inhibitor DPI abolished the observed effect. Previously it has been shown by us that differentiating ES cells express Nox-1, Nox-2, and Nox-4. These Nox enzymes endogenously generate ROS, which are involved in cardiovascular differentiation [30]. The elevation in ERK1/2 activity after exposure to static EMFs may be due to inhibition of phosphatases that normally counteract ERK activation [43]. Such inhibition of phosphatases could be due to the production of ROS, which have previously shown to counteract phosphatases of the MAPK cascades [44]. Changes in activation of MAPK observed in the present study agree with earlier studies of others. It has been shown that prolonged exposure to EMFs can activate the p38 MAPK, JNK, and ERK1/2 cascades [33,45,46]. Exposure to EMF rapidly activated ERK1/2, p38, and JNK MAPK by generating ROS from NAD(P)H oxidase [47], although a reduction in p38 MAPK levels has also been reported [48].
Using signaling inhibitors the results of the present study suggest that the ERK1/2 pathway is involved in the activation of VEGF following EMF treatment. We demonstrated increased VEGF mRNA and protein expression upon treatment of ES cells with 1 mT EMFs during chondro-osteogenic differentiation. ROS-induced VEGF expression was apparently crucial for chondro-osteogenic differentiation in ES cells. In this respect inhibition of NAD(P)H oxidase as well as interfering with VEGF-R2 signaling by treatment of differentiating ES cells with SU5614 abolished the observed effects. The involvement of VEGF in angiogenesis processes and osteoblastic differentiation or function during bone development was previously reported by several authors [12,13,20,49]. Their data suggest that blood vessel invasion into the primary ossification center is a key step in bone development during in vivo embryogenesis. Interestingly high EMF strength of 10 mT exerted adverse effects by inhibiting blood vessel invasion into the hypertrophic zone of cartilage nodules thus resulting in decrease of bone formation and mineralization. The adverse effects of high EMF field strength could be explained by inhibition of VEGF expression under these conditions, which resulted in an antiangiogenic effect of EMFs. This may be caused by an overrun of the antioxidative capacity of the cells by the oxidative stress exerted by 10 mT EMF strength. Consequently, preincubation of embryoid bodies with trolox prior to 10 mT EMF treatment resulted in a significantly increase in the expression level of VEGF even above the expression level of the untreated control. Hence our data support the notion that ROS accumulation beyond the antioxidant capacities of the cells may lead to inhibition of VEGF expression and production. Therefore the results of the present study agree with several authors who previously showed that oxidative stress could be involved in VEGF inhibition [50 –52].
Our observations on the modulation of VEGF expression by static EMFs through mechanisms involving ROS strengthen the hypothesis that VEGF is a principal regulator for many processes that normally occur during bone development. VEGF thereby not only mediates bone vascularization but also allows normal differentiation of hypertrophic chondrocytes, osteoblasts, endothelial cells, and osteoclasts. These observations are in conformity with those of Xu et al. [53], who found that the enhancement of femoral bone formation in the ischemic rat model by static magnetic fields seems to be due to the improved blood circulation in the femur. Moreover, Deckers et al. [54] confirmed the role of VEGF in differentiation of osteoblasts. They suggested that inactivation of VEGF inhibited endochondral bone formation via inhibition of angiogenesis. VEGF controls at least three aspects of bone development [54]. First, it induces angiogenesis in regions of intramembranous bone formation and in the perichondral regions of cartilage templates in the endochondral skeleton. Second, it stimulates angiogenesis and chemotactic migration of osteoclasts into hypertrophic cartilage. Finally, VEGF stimulates bone formation by increasing the activity of osteoblasts in intramembranous as well as endochondral bones. This notion was substantiated by in vivo studies that provided evidence for the important role of VEGF in blood vessel invasion into hypertrophic cartilage during bone development [20]. Furthermore it was shown that inhibition of all VEGF isoforms in cultures of embryonic cartilaginous metatarsals through the administration of a soluble receptor chimeric protein (mFlt-1/Fc) retarded the onset and progression of ossification, which was probably mediated by impairment of osteoblast and/or hypertrophic chondrocyte development [19]. There is also suggestive evidence from in vitro experiments that VEGF may regulate bone formation through a direct effect on osteoblasts. It has been reported that human VEGF165 binds to osteoblasts in culture and is capable of inducing migration and alkaline phosphatase activity but does not affect proliferation of these cells [16]. Indeed, VEGF-R1 and VEGF-R2 have been reported to be expressed on osteoblasts [54]. Not only does VEGF act upon osteoblasts in culture but osteoblasts also produce VEGF when stimulated by vitamin D3 [55]. Furthermore, VEGF seems to be a chemoattractant for osteoclasts [56].
The data of the present study conclusively demonstrate that EMF-induced ROS modulate VEGF expression, which is crucial for hypertrophic cartilage neovascularization in differentiating ES cells, with invading endothelial cells as a target. Furthermore, the previously described VEGF receptor localization in chondrocytes strongly supports the hypothesis of the role of VEGF activity in chondro-osteogenic differentiation in ES cells under static EMF conditions. Our data provide further support for clinical applications of EMFs to improve bone healing and give insight into the so far not well investigated molecular mechanisms. However, our data additionally provide evidence that in vivo EMFs of high field strength may give rise to bone defects due to the generation of oxidative stress. Oxidative stress may interfere with VEGF expression levels that are not only required during embryogenesis for the formation of the cardiovascular system but also for the proper formation of a mineralized skeleton.
Footnotes
Acknowledgments
This work was supported by the SFB 604 “Multifunctional Signalling Proteins,” the excellence cluster “Cardiopulmonary System” (ECCPS) of the German Research Foundation (DFG) and the German Foundation for Heart Research.
Author Disclosure Statement
No competing financial interests exist.