
Abstract
Hepatic oval cells are thought to represent facultative hepatic epithelial stem cells in liver in which damage inhibits hepatocyte proliferation and liver regeneration. The LE/6 hepatic stem cell line was derived from the liver of male Sprague-Dawley rats fed a choline-deficient diet containing 0.1% ethionine. They are histochemically characterized by their expression of hepatocytic (hepPar1), cholangiocytic cytokeratin (CK19), hepatic progenitor cell (OV-6), and hematopoietic stem cell (c-kit) markers. In this study, we transplanted LE/6 cells by subcutaneous injection into adult female nude mice, and examined their engraftment and differentiation potential in the subcutaneous microenvironment in vivo. Our results demonstrated that following subcutaneous transplantation, differentiation of LE/6 cells into mesenchymal tumor tissue (MTT) was associated with reduced E-cadherin expression, upregulation of E-cadherin repressor molecules (Snail proteins), and increased expression of vimentin and N-cadherin, all of these events are characteristic of the epithelial–mesenchymal transition (EMT).
Introduction
S
The epithelial–mesenchymal transition (EMT) is a key developmental program that is often activated during early embryonic development, or during cancer invasion and metastasis. During EMT, E-cadherin is downregulated and this correlates with increased motility and invasion of cells. EMT is one of the mechanisms by which epithelial cells acquire the motile properties required for invasion [11,12]. Acquisition of the mesenchymal state, such as the fibroblastic phenotype, is accompanied by downregulation of E-cadherin, and upregulation of mesenchymal markers such as vimentin, smooth muscle actin (SMA), desmin, S-100, and N-cadherin, thus enabling cells to dissociate from the epithelial tissue and migrate.
EMT is mainly regulated by members of transcription inhibitor superfamily, particularly the Snail family that directly induces EMT through the inhibition of E-cadherin gene transcription. The Snail family of transcription factors (Snail, Snailp, Slug, Scratch1, and Scratch2) comprises several proteins that contain 4–6 zinc finger domains and share a high degree of homology across several species. In Drosophila, these proteins mediate the development of the mesoderm, neuroectoderm, and other organs [13]. Previous studies have documented the role of the Snail family in mediating the loss of epithelial adhesion molecules [2 –6]. Snail binds to E-boxes present in the E-cadherin promoter, consequently repressing E-cadherin transcription [14,15]. In some human cancer–derived cell lines, Snail-mediated EMT promotes cell motility and invasion [13]; for example, an inverse correlation between E-cadherin and Snail expression has been noted in cultured lines established from oral squamous cell carcinoma [16], melanoma [17], pancreatic carcinoma [18], and hepatocellular carcinoma and cholangiocarcinoma [19]. Recently, histological evidence of EMT in human tissue samples has also been reported; in gastric cancer and hepatocellular carcinoma, EMT regulator-positive tumors exhibited low levels of E-cadherin expression [17,20].
In this study, we present evidence that LE/6 cells modulate their lineage commitment by EMT following subcutaneous transplantation, when they possess an apparent broader capacity for cellular differentiation that is dependent on, and responsive to, the specific signals present in the microenvironment of the transplantation site.
Materials and Methods
Cell culture and transplantation
LE/6 cells were a gift from Prof. Nelson Fausto. Hepatic oval cells from the livers of 6- to 8-week-old male rats fed a choline-deficient diet for 6 weeks were isolated as described previously by Yaswen et al. [21]. LE/6 cells were cultured in 10-mm dishes in Dulbecco’s minimal essential medium: Ham’s F-10 (1:1) (Invitrogen Co., Carlsbad, CA), supplemented with 10% fetal calf serum (Gibco Laboratories, North Andover, MA), 1 µg/mL insulin (Sigma Chemical Co., St. Louis, MO), 0.5 µg/mL hydrocortisone (Sigma Chemical Co.), and 25 µg/mL gentamicin at 37°C in a 5% CO2 atmosphere. Cells intended for transplantation were harvested with trypsin containing 0.02% EDTA, washed in cell culture medium, and resuspended in serum-free medium. Cells were injected into both sides of the subcutaneous space of the armpits of female nude mice (BALB/cA nu/nu). Each side was injected with 5 × 106 cells in a volume of 0.1 mL, using a 1-mL syringe fitted with a needle. Control animals were handled identically, and received sham injections of serum-free medium. We also included a positive control group in which HepG2 hepatoma cells were used instead of LE/6 cells.
Histochemistry and immunohistochemistry
Five months after donor cell engraftment, recipient nude mice were killed. The transplanted tumors were dissected and sliced coronally into three blocks, one of which was fixed in formalin and embedded in paraffin, while the others were immediately frozen and stored in liquid nitrogen. Sections of paraffin-embedded tissues (5 µm in thickness) were used for immunostaining. Hematoxylin and eosin (H&E) staining was performed routinely on the immunostained sections. Tissue sections were deparaffinized in xylene and rehydrated through a graded series of alcohols. Endogenous peroxidases were inactivated by immersing sections in 3% H2O2 in methanol for 20 min. Antigen retrieval was performed in 10 mmol/L Tris buffer, and 1 mmol/L EDTA (pH 9.0) for 20 min in a microwave oven at 98.5°C. The sections were then incubated with primary antibody for 60 min at room temperature, and subsequently with goat anti-mouse EnVision (Dako Corp., Carpinteria, CA) for 30 min at room temperature. Both incubation steps were followed by a wash and were developed with 3-amino-9-ethylcarbazole (DAB) and counterstained with hematoxylin.
Immunohistochemical staining was performed on all tissue sections with the following antibodies: hepPar1 (monoclonal antibody specific for hepatocytes; 1:50; Dako Corp.); CK19 (monoclonal antibody specific for cholangiocytes and oval cells; 1:100; Dako Corp.); PCK (broad-spectrum anti-keratin polyclonal antibody; 1:50; Dako Corp.); OV-6 (monoclonal antibody specific for oval cells; 1:10; R&D Systems, Minneapolis, MN); c-kit (polyclonal antibody positive in hematopoietic stem cells and oval cells; 1:200; Santa Cruz Corp., Baltimore, MD); vimentin (monoclonal antibody specific for mesenchymal origin in normal and neoplastic tissues; 1:100; Dako Corp.); SMA (monoclonal antibody, identification of leiomyosarcomas; 1:50; Dako Corp.); desmin (monoclonal antibody specific for smooth muscle and mesothelial cells; 1:50; Dako Corp.); and anti-S100 (Polyclonal Rabbit, positive in myocytes; Dako Corp.).
Transmission electron microscopy
Fresh tumor sections were fixed in 2.5% (w/v) glutaraldehyde in 0.1 M cacodylate buffer (pH 7.4). After extensive washing in phosphate-buffered saline (PBS), the sections were post-fixed in 1% (w/v) osmium tetroxide (OsO4) for 30 min, dehydrated in a graded series of ethanol solutions, and embedded in Epon. Ultrathin sections were cut and examined under a Zeiss EM10C electron microscope after staining with uranyl acetate and lead citrate.
Fluorescence in situ hybridization
Paraffin-embedded tissue sections were deparaffinized, rehydrated through graded alcohols to water, incubated with sodium thiocyanate solution for 10 min at 80°C, and then rinsed in PBS. The slides were digested in pepsin solution (4 mg/mL in 0.1 M HCl) for 15 min at 37°C, following which the pepsin was quenched by incubation in glycine solution. After rinsing the slides in PBS, they were post-fixed in paraformaldehyde solution for 2 min at room temperature, washed well in PBS (three changes over 15 min), dehydrated through graded alcohols and air dried. A commercial cyanine-3 (Cy3)-labeled probe (Cambio Corp., Cambridge, UK) was used for the direct visualization of rat Y-chromosomes (red signal). The probe was denatured in advance at 65°C for 10 min and kept at 37°C for 30–60 min before use, then applied to the center of the slide, covered with a glass coverslip, and sealed with rubber cement. The probe and target DNA were denatured simultaneously at 72°C for 10 min, followed by overnight incubation in a humid chamber at 37°C. Slides were washed at high stringency, counterstained with 4,6-diamidino-2-phenylindole (DAPI) and mounted in antifade. The slide was evaluated on a Nikon TE2000-U fluorescence microscope (Nikon Corp., Japan) fitted with red and blue filters (Cy3/DAPI). Images were collected using a CytoVision system (NIS-Elements F 3.0; Nikon Corp., Japan).
Double-fluorescence immunostaining
Deparaffinization, rehydration, and heat-induced epitope retrieval of the transplanted tumor tissue sections were performed as described above. The primary antibody combinations were Snail plus N-cadherin and Snail plus vimentin. Secondary antibodies (goat anti-mouse IgG or goat anti-rabbit IgG) conjugated with Cy3 or fluorescein isothiocyanate were used. Double labeling using both primary antibodies on the same section was performed. The primary and secondary antibody staining procedures were performed according to the instructions from the Vector M.O.M™ Fluorescein Immunodetection kit (Vector Laboratories, UK) and Avidin/Biotin blocking kit (Vector Laboratories, UK). Controls in which the primary antibodies were omitted were consistently negative. Immunofluorescence was observed using a Nikon Digital ECLIPSE C1 microscope (Nikon Corp., Japan). An argon laser at 488 nm in combination with a helium neon laser at 543 nm were used to excite the green and red fluorochromes simultaneously.
Western blot analysis
Fresh frozen transplanted tumor tissues were lysed in RIPA buffer using a tissue homogenizer. Equal amounts of protein (16 µg) were separated on 10% SDS-polyacrylamide gels and transferred on to PVDF membranes. The blots were probed with antibodies against Snail (1:200; Santa Cruz Biotechnology Inc., Santa Cruz, CA), E-cadherin (1:500; BD Biosciences Pharmingen, San Diego, CA), vimentin (1:300; Sigma, St. Louis, MO), N-cadherin (1:2,000; Zymed Laboratories, South San Francisco, CA), and β-actin (1:3,000; Santa Cruz Biotechnology Inc.).
Results
Transplantation results
From a total of 50 nude mice inoculated with LE/6 cells, 22 developed transplanted tumors (44% positive rate). Visible nodules formed at the site of inoculation within 3 months, and grew slowly at a rate of 0.5–1 mm/month. The size of the tumors increased with time, and no tumors were observed to disappear spontaneously. The largest tumors reached an average diameter of 1 cm (Fig. 1). In contrast, the five nude mice in the control group did not develop tumors, while all five nude mice in the positive control developed tumors. The growth of tumor tissues in the positive control group was different from the LE/6 cell tumors, with visible nodules appearing after about 2 weeks, and tumors reaching an average diameter of 2 cm within 4–5 weeks.
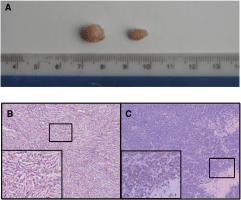
(
Histological features
The expression of hepPar1, CK19, PCK, OV-6, or c-kit was undetectable in sections prepared from LE/6 tumors. The expression of all of these markers was confirmed in LE/6 cells by immunofluorescence microscopy. All tumor sections were immunopositive for vimentin, SMA, desmin, and S-100 (Fig. 2).
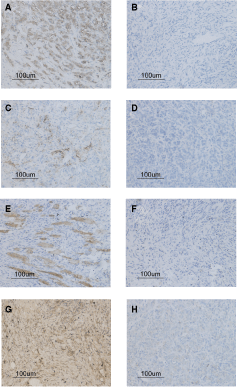
Immunohistochemical features of vimentin, smooth muscle actin (SMA), desmin, and S-100 in mesenchymal tumor tissues (MTTs). The MTTs were immunopositive for vimentin (
Together with the immunohistochemistry data, H&E staining of tumor tissue sections from the 22 mice that developed tumors following LE/6 cell transplantation confirmed that the LE/6 cells had transdifferentiated into mesenchymal tissue (Fig. 2). Histopathologically, the tumors were separated into nodules by septa. The margins of the tumors appeared to be clearly delimited from the surrounding muscle, fat, and fibrous tissues. The cells were loosely packed and unpolarized with morphologies reminiscent of spindle-shaped stromal or mesenchymal cells. The cells were arranged in an irregular, whirlpool-like formation, and possessed an eosinophilic cytoplasm and slender, rod-shaped nuclei. Only a few mitotic cells were present, indicating that these subcutaneous tumors were mainly highly differentiated mesenchymal tissue tumors. This was confirmed by transmission electron microscopy, which revealed the presence of the dense cytoplasmic bodies that are a specific marker of mesenchymal tissue. Transmission electron microscopy also revealed that the cells possessed atypical nuclei and hyperchromasia. The cells were relatively small with a median diameter of 6–8 µm, less than half that of mature hepatocytes (Fig. 3).

(
Fluorescence in situ hybridization profiles
We confirmed that the tumor tissues were derived from LE/6 cells by demonstrating that they contained a rat Y-chromosome by FISH analysis with a rat Y-chromosome-specific repetitive DNA sequence (Fig. 4). As a positive control, we harvested cultured LE/6 cells and confirmed that they contained this sequence by karyotype analysis. We also used adjacent 5-µm-thick serial sections to demonstrate the histopathologic features of the tumor tissues (Fig. 2), and observed the presence of a Cy3-labeled Y-chromosome in the nucleus of almost all cells.

Fluorescence in situ hybridization showing the presence of red Y-chromosomes and blue nuclei in the same sections. The nuclei exhibit the different sizes and shapes also observed by hematoxylin and eosin staining. Original magnification, ×1,000.
Double-fluorescence immunostaining profiles
The cells of the tumor tissues exhibited simultaneous cytoplasmic coexpression of Snail with vimentin, and Snail with N-cadherin, by confocal double-fluorescence immunostaining (Fig. 5).
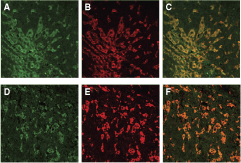
Double-immunofluorescence staining showing the expression of Snail (fluorescein isothiocyanate, green;
Western blot profiles
The cells of the tumor tissues expressed Snail, vimentin, and N-cadherin, but only low levels of E-cadherin were detected. However, HepG2 tumor and LE/6 cells exhibited low levels of Snail, vimentin, and N-cadherin protein, but expressed E-cadherin at extremely high levels. LE/6 cells expressed E-cadherin and vimentin, but exhibited low levels of Snail and N-cadherin (Fig. 6).
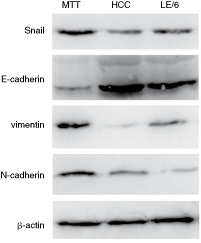
Western blot analysis of Snail, E-cadherin, vimentin, N-cadherin, and β-actin. Abbreviations: MTT, mesenchymal tumor tissue; HCC, tumor from HepG2 cells; LE/6, LE/6 cells.
Discussion
The aim of this study was to test whether the subcutaneous microenvironment can induce the differentiation of HOV cells into other cell phenotypes in vivo. In vitro and in vivo experiments confirmed that HOV cells can differentiate into hepatic cells and biliary epithelial cells, cardiac muscle cells, pancreatic cells, glial cells, and prolactin cells [1,7,9,22,23]. Because we know that HOV cells are an epithelial cell type, our results provide evidence that HOV cells can differentiate into mesenchymal cells by EMT. These findings are consistent with the hypothesis that reactivation of genetic programs in adult stem cells is possible in response to appropriate stimulation. These results indicate that the microenvironment and cell extrinsic factors are likely to play a role in stem cell plasticity, although the transcription factors are a key regulator of cell fate.
EMT is defined as the transdifferentiation of epithelial cells to mesenchymal cells, and has emerged as a central process contributing to embryonic development, chronic inflammation, fibrosis, and cancer progression [12,13]. Loss of cell adhesion, increased cell motility, and reversible dedifferentiation are even more marked in the process of wound healing, in embryogenesis during the migration of neural crest cells, and in gastrulation [24] when the mesoderm is formed. A characteristic feature of EMT is the disruption of normal epithelial cell–cell contacts, which is achieved by a reduction in E-cadherin levels [25]. The loss of E-cadherin expression is a critical step for EMT. EMT results in the complete loss of epithelial traits, including the loss of E-cadherin and cytokeratins, accompanied by the total acquisition of mesenchymal characteristics, including the expression of vimentin, SMA, and N-cadherin [24].
It has been documented that Snail functions as a direct repressor of E-cadherin to promote EMT in several cell types [14,15,26,27]. Snail family transcription factors bind to consensus E-box sequences in the E-cadherin gene promoter and downregulate E-cadherin transcription [28]. Transfection of Snail cDNA into MDCK cells induced epithelial to mesenchymal transitions [29], transfected cells acquired motile and invasive properties concurrent with transcriptional downregulation of E-cadherin, and the morphological changes induced by Snail expression suggested that adherens junctions, tight junctions, and desmosomes are all disrupted by Snail expression. These findings suggest that Snail also plays an important role in EMT through Snail-mediated transcriptional downregulation of E-cadherin.
In this study, differentiation of LE/6 cells into mesenchymal tumor tissue (MTT) occurred with a reduction in the expression of E-cadherin, while the endogenous expression levels of Snail, vimentin, SMA, and N-cadherin were characteristic of EMT. Evidence of Snail expression–associated EMT has not been well documented. To elucidate the existence of EMT in LE/6 cells, we performed double-fluorescence immunostaining analysis using Snail, vimentin, and N-cadherin antibodies. We found that Snail expression was significantly associated with both vimentin and N-cadherin expression.
In conclusion, our results suggest that undifferentiated HOV cells derived from liver tissues are not determined progenitor cells with limited differentiation potential. Rather, these cells seem to possess a much broader capacity for cellular differentiation that is dependent on, and responsive to, the specific signals present in the microenvironment of the inoculated site. We also provide evidence that EMT promoted HOV cell transdifferentiation into MTT. EMT provides HOV cells with migration and invasive activity, and this is indirect evidence that HOV cells can differentiate into liver cancer cells.
Footnotes
Acknowledgment
This work was supported by a grant from the Key Project of National Natural Science Foundation of China (Grant no. 30430670).
Author Disclosure Statement
No competing financial interests exist.