
Abstract
Pluripotency of stem cells refers to a stem cell that has the potential to differentiate into any of the three germ layers: endoderm, mesoderm, or ectoderm. Maintaining pluripotent stem cells in culture is a tedious and demanding task. Monitoring the changing pluripotency in live cells is essential for this task. Here, we report a pluripotency monitoring system in which the expression of green fluorescent protein (GFP) is under the control of the promoter of a pluripotency gene (Rex-1). The reporter system can be permanently integrated into the genome of live cells via lentiviral vectors. This pluripotency reporter system permits the long-term real-time monitoring of pluripotency changes in a live single cell and its progeny. Our data demonstrate that the BJ cell line (a normal human fibroblast cell line) that carries this hRex-GFP construct does not express GFP until it is reprogrammed to pluripotent stage. The GFP expression was progressively lost when these pluripotent hRex-GFP cells exposed to differentiation conditions. These results indicate that insertion of the hRex-GFP construct is stable in descendant cells, a finding that has particular value in tracking pluripotency of transplanted cells and their progenies in animal studies. With this hRex-GFP reporter, the pluripotency of cells can be monitored over long periods of time via the expression of GFP. Use of this reporter system will facilitate the study of stem cell pluripotency at the single-cell level, and sheds light on the molecular mechanisms of stem cell self-renewal and subsequent differentiation.
Introduction
P
Various experimental techniques have been developed for deriving pluripotent stem cells, including somatic cell nuclear transfer [3], cell fusion [4], and somatic cell reprogramming with defined transcription factors [5,6]. Many protocols have also been developed to maintain proliferation of pluripotent cell, the self-renewal of stem cells. However, maintaining the pluripotent cells in tissue culture is a tedious and expensive task. Spontaneous differentiation is always observed in stem cell cultures, and pluripotent cells are maintained by removing such differentiated cells during serial passage. In addition, recent single-cell analysis of human embryonic stem cell (hESC) colonies indicated that a pluripotent hESC colony is a heterogeneous cells population [7]. Therefore, close monitoring of the changing pluripotency of cells in culture is essential for many studies.
Pluripotency of stem cells can be identified by RT-PCR of embryonic stem cell (ESC) markers (ie, Oct-3/4, Sox2, Nanog, Rex-1), immunochemistry with stage-specific embryonic antigens (SSEA-3, SSEA-4, TRA-l-60, TRA-1-81), alkaline phosphatase activity, and methylation profiles of promoters of pluripotent-associated genes [2,5,6]. However, these methods often involve destruction or fixation of cells during analysis, and cannot be used to monitor the pluripotency of live cells. Although immunoassaying with antibodies recognizing stage-specific embryonic antigens can be used with live cells, degradation or endocytosis of antibodies prevent long-term monitoring. Here we describe the development of a pluripotency reporter for monitoring any changes in the pluripotency of live cells in real time and at single-cell resolution.
We constructed a pluripotency reporter that contains a green fluorescent protein (GFP) gene under the control of the human Rex-1 promoter. The reporter can be permanently integrated into the genome of live cells via lentiviral vectors. Rex-1 (Zfp42) was first identified in murine F9 embryonal carcinoma cells, and is down-regulated during F9 differentiation [8]. Human Rex-1 encodes a C2H2 zinc finger protein, and is a member of the YY1 (Yin-Yang 1) subfamily of transcription factors. It contains four cysteine–histidine zinc finger domains [9]. The human and mouse Rex-1 proteins display 57% amino acid sequence identity, and the mouse Rex-1 promoter region is well defined [10]. Several “pluripotency-specific” proteins (Nanog, Sox2, and Oct-3/4) co-occupy common sites on the mouse Rex-1 promoter region [11,12]. However, the promoter region of human Rex-1 is less clear. Employing bioinformatics approaches, we identified the human Rex-1 (hRex-1) promoter sequence, and confirmed the function of the hRex-1 promoter in human hESC and fibroblasts. Our data demonstrate that the hRex-GFP reporter is completely silent in differentiated cells, and only expresses GFP in pluripotent cells.
We have generated lentiviral vectors for stable integration of this hRex-GFP construct into human genomes. The lentiviral vectors can permanently integrate the hRex-GFP pluripotency reporter into the genomes of dividing and nondividing human cells. This pluripotency reporter system permits the long-term real-time monitoring of pluripotency changes in a live single cell and its progeny. Use of this reporter system facilitates the study of stem cell pluripotency at the single-cell level, and sheds light on the molecular mechanisms of stem cell self-renewal and subsequent differentiation.
Materials and Methods
Identification of human Rex-1 promoter
The mouse Rex-1 promoter region is well defined and studied, but the corresponding human Rex-1 promoter is less well known. Aligning the mouse promoter sequence to human sequence failed to identify significant homology. Therefore, we located a 5 kb region upstream of the star codon of the human Rex-1 gene from NCBI database, and then searched for sequences similar to the Oct-3/4-binding octamer motif (ATTTGCAT) and GC boxes (CCCGCCC), which are reported in mouse Rex-1 promoter [10], using pattern search software as described (http://bioweb.pasteur.fr/seqanal/interfaces/scan_for_matches.html). Patterns with significant matches were scored for regions of ∼800 bp. Five potential human Rex-1 regions were selected for producing the hRex-GFP constructs that were then tested in human BJ cells (normal human fibroblasts).
Construction of Rex-GFP lentiviral vectors
A lentiviral construct pL_UGIP in which GFP expression is under the control of Ubi promoter was purchased from ATCC (ATCC #10326372). The Ubi promoter was removed by digesting the vector with AgeI and PacI (New England Biolab, Ipswich, MA, USA). Mouse rex-1 promoter and the five potential human Rex-1 promoters were amplified from human and mouse genomic DNA, respectively (Clontech, Mountain View, CA, USA), with corresponding PCR primers containing restriction sites. The PCR products were first cloned into pCR2.1 vector (Invitrogen, Carlsbad, CA, USA) and digested with AgeI and PacI (New England Biolab, USA) to obtain the respective promoter regions. After electrophoresis, different promoter fragments were purified using an agarose gel extraction kit (Qiagen, USA). The purified promoter fragments were subcloned into the pL-UGIP backbone.
The sequence and orientation of mouse and human Rex 1 promoters were confirmed by DNA sequencing. The resulting mouse and human Rex-GFP constructs were then tested in human ESC and BJ cells to identify the human Rex-GFP construct that best served as a pluripotency reporter.
Primers for amplifying mouse Rex-1 promoter are: CATATCCTTAATTAAACCGATTCCTC (forward) and CTATCACCGGTTCTAGGCGGCTAGGAG (reverse); primers for amplifying human Rex-1 promoter are: CATATC CTTAATTAATGCAGAAAAGC (forward) and CTATCA CCGGTACAAATACGCGTGGGTGTAA (reverse).
Cell culture
BJ cells were purchased from ATCC (CRL-2522) and maintained as instructed in Eagle’s minimum essential medium (ATCC) with 10% fetal bovine serum (FBS). The HES3 were obtained from the stem cell core facility at the University of Southern California. The HES3 cells were cultured on mouse embryonic fibroblast (MEF) feeders with hESC medium: DMEM supplemented with 5% FBS (Hyclone, USA), 1% nonessential amino acid, 1% Insulin–Transferrin–Selenium-G Supplement, and for each 100 mL medium 0.5 mL Pen/Strep and 0.18 mL of 2-mercaptoethanol were added (except FBS, all components were purchased from Invitrogen). Cell passages were performed with microdissection of colonies. A sterile 25-G needle was used to remove the center of each colony for re-plating into fresh MEF feeder each week. The hESC medium was changed everyday. The H9 line was purchased from WiCell and maintained as instructed.
Transduction of human ESCs and fibroblasts
Retroviral vectors were produced with three-plasmid transient transfection using a 10-cm plate of 293T cells. The 293T cells were split the previous day with a 1:5 ratio. The calcium 6-phosphate precipitation method was performed with 2 µg vesicular stomatitis virus glycoprotein (VSVG) envelope expression plasmid, 10 µg of pCMV delta 8.91 (the gag, pol plasmid), and 10 µg of the lentiviral expression vector plasmid. The medium was changed the following morning and the respective lentiviral vectors were collected 24 h later by filtration through a 0.2-µm filter.
One milliliter of the vector supernatant plus 8 µg/mL polybrene were added to each well of a 24-well dish containing 2 × 104 fibroblasts or HES3 on MEF feeders. After incubation with the respective vector for 2–4 h, transduction medium was replaced with 2 mL growth medium (EMEM with 10% FBS) for fibroblasts, or 2 mL of hESC medium for transduced HES3.
Clonal selection of BJ cells with hRex-GFP insertion
The hRex-GFP construct did not express GFP in somatic BJ cells. After transduction with hRex-GFP lentiviral vectors, BJ cells were prepared as single-cell suspensions using trypsin. A serial dilution was made to plate single BJ cells into 96-well plates. After expansion to ∼500 cells in wells, half the cells were removed mechanically with a pipette tip. Genomic DNA was extracted with DNeasy Kit (Qiagene, USA) and subjected to PCR analysis with GFP-specific primers: GACGTAAACGGCCACAAGTT (forward) and GAACTCCAGCAGGACCATGT (reverse).
Reprogramming and differentiation of hRex-GFP fibroblasts
Cell fusion was performed to reprogram BJ cells carrying the Rex-GFP construct. To induce cell fusion, 5 × 107 hESC (H9) and 5 × 107 transduced fibroblasts were mixed in the presence of polyethylene glycol (PEG; Roche, Nutley, NJ) as instructed by the manufacturer. The cell fusion mixtures were plated on MEF feeder under standard conditions for the maintenance of hESCs as described [2]. After 5 days, both GFP-positive colonies (reprogrammed hESC) and GFP-negative colonies (not reprogrammed) were observed. Both types of colonies were subjected to alkaline phosphatase assays (VECTOR Blue Substrate Kit, VECTOR Labs). The GFP-positive colonies were picked mechanically. To differentiate the reprogrammed cells, GFP-positive colonies were plated on a regular Petri dish with DMEM supplemented with 10% FBS.
Results
Silence of hRex-GFP in fibroblasts
Rex-1 was found to be highly expressed in pluripotent human and murine cells, and its expression decreased as the cells differentiated [9,10,13]. Although the regulatory role of Rex-1 in stem cell pluripotency is unknown, it was reported that it is only expressed in pluripotent cells [13,14]. We transduced BJ cells with lentiviral vectors containing Ubi-GFP and human Rex-GFP constructs. Among six constructs, the mouse Rex-GFP and three human Rex-GFP constructs showed low GFP expression in BJ cells. Therefore, we selected one human Rex-GFP construct with no detectable GFP expression in five independent transduction experiments, and designated this construct as hRex-GFP. As shown in Figure 1, BJ cells incorporating the Ubi-GFP construct express GFP in transduced cells (Fig. 1). However, BJ cells transduced with hRex-GFP vectors did not express GFP (Fig. 2).
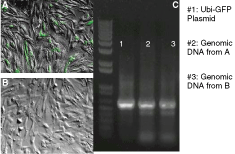
Fibroblasts with hRex-GFP construct do not express GFP. (
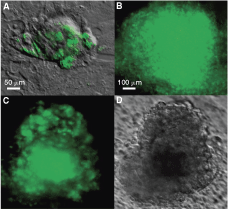
Expression of GFP from the hRex-GFP construct is stable in pluripotent human embryonic stem cells (hESCs) for at least eight passages. HES3 is maintained in pluripotent status. (
For verifying the incorporation of vector constructs into the genomes, genomic DNA was extracted from transduced BJ cells and subjected to PCR with GFP-specific primers. To rule out the possibility of vector DNA contamination, genomic DNA was prepared from transduced BJ cells that were at least three passages post-transduction. Electrophoresis of PCR products indicated that GFP sequence is present in genomic DNA of BJ cells transduced with Ubi-GFP or hRex-GFP vectors. The lack of GFP expression in BJ cells and the confirmation of GFP sequence incorporation show that the hRex-GFP construct is silent in BJ cells. This result agreed with previous reports that BJ cells do not express Rex-1 [4,15].
Stable expression of hRex-GFP in hESCs
In order to use the hRex-GFP construct as a real-time pluripotency reporter, the construct must be silent in somatic cells and have stable GFP expression in pluripotent cells, such as hESC. After transducing hESC (HES3) on MEF feeders with the hRex-GFP vector, patches of green cells appear (Fig. 2A) inside the pluripotent HES3 colonies ∼7 days post-transduction. The GFP-positive cell patches were dissected and re-plated onto MEF feeder to obtain GFP-positive hESC colonies (Fig. 2B). These GFP-positive hESC colonies were passaged by dissecting out the center of the colonies and re-plated onto MEF feeders. After eight passages, the colonies continued to express GFP (Fig. 2C). These data indicate the stable expression of GFP in hESC. We also observed that GFP expression is gradually lost in the cells located at the edge of the hESC colonies, suggestive of spontaneously differentiation at the colony periphery. Therefore, we further investigated whether the loss of GFP expression in these peripherally located cells is reflective of cell differentiation.
Lost of GFP expression in differentiated hRex-GFP cells
To initiate differentiation of HES3 colonies, hESC medium was replaced with DMEM medium supplemented with 10% FBS. Cells in the edge of the GFP-positive colonies rapidly lost GFP expression, while cells in the center of the colonies continued to express GFP (Fig. 3A). After exposure to differentiation conditions for 4 days, the cellular morphology of the colonies also changed from round to irregular in shape. Eventually most of the cells lost GFP expression, but the cells located in the center continued to express GFP. During HES3 culture maintenance, we observed that differentiation of a pluripotent colony often progressing from periphery toward the colony center. Therefore, these data suggest that only pluripotent cells with the hRex-GFP insertion express GFP. Differentiated cells with hRex-GFP insertion do not express GFP.
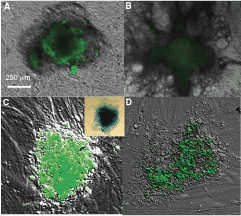
GFP expression from hRex-GFP construct is reduced during differentiation of pluripotent cells. To induce differentiation, human embryonic stem cell (hESC) medium was replaced by DMEM with 10% FBS. (
To investigate further the specificity of hRex-GFP as a pluripotency reporter, we reprogrammed BJ cells that have the hRex-GFP insertion by a cell fusion technique [4]. These BJ cells do not express GFP when they are cultured as fibroblasts (Fig. 1B). After fusing hRex-GFP BJ cells with H9 cells, the hybrid cells are plated on MEF feeder following which GFP-positive colonies appeared ∼5 days after fusion (Fig. 3C). We further isolated and re-plated GFP-positive colonies on MEF feeders in chamber slides. Alkaline phosphatase (AP) staining of GFP-positive colonies indicates that these colonies have strong AK activity and are pluripotent (Fig. 3, inset). The reprogrammed hybrid cells were then exposed to differentiation medium to induce cellular differentiation. As expected, when the GFP-positive hybrid cells underwent differentiation, they lost GFP expression. The lost of GFP expression began ∼48 h after exposing to differentiation medium, and progressed from the edge of the colonies toward the center (Fig. 3B and D). These data indicate that the hRex-GFP construct can be inserted into the genome of human fibroblasts via lentiviral vectors, and the insertion is stable and functional in progeny of these fibroblasts. Therefore, the hRex-GFP construct can be used for long-term and real-time monitoring of pluripotency in live cells.
Discussion
Pluripotent stem cells are cells that can differentiate into any type of tissue in the human body. These pluripotent cells not only have great potential in regenerative medicine, but also provide the opportunity to study and understand human development. Developing an understanding of the underlying molecular mechanisms of pluripotency and differentiation is particularly important, because it may provide knowledge that will allow specific manipulation of the development of human cells, to direct stem cells for generating specific cell types, and to understand abnormal cell development in conditions such as cancer.
However, a simple assay of pluripotency is not yet available. Current assays for pluripotency such as teratoma assay, immunochemical profile, or gene expression assay with RT-PCR cannot monitor the pluripotency in live cells in real time. Attempts to use gene manipulation for monitoring pluripotency have been made. Genetic manipulation methods in transgenic mice have successfully introduced GFP into mouse genomes under the control of pluripotency-related genes such as Oct-3/4 [16] and Nanog [16 –18]. However, such transgenic techniques cannot be applied to humans. Transient transfection of a plasmid with expression of GFP under the control of mouse Rex-1 promoter has also been used to monitor the pluripotency of cells [4,19]. Unfortunately, the transfected plasmid cannot be passed on with cell division, and monitoring pluripotency in progeny cells as they undergo differentiation is not possible.
Here, we report a lentiviral deliverable system for monitoring pluripotency of human cells in real time and in multiple-cell passages. The system is based on GFP expression under the control of human Rex-1 promoter. The human Rex-1 promoter did not show significant sequence homology to its previously reported mouse counterpart [10]. However, with bioinformatics approaches, we found five potential human Rex-1 promoter regions, that were then cloned into lentiviral expression vectors, to control the expression of GFP. These were functionally tested in human fibroblasts, BJ cells. One promoter, hRex-GFP, was identified as the best functional human Rex-1 promoter. We further tested the stability and specificity of the hRex-GFP construct in pluripotent hESC and in reprogrammed human fibroblasts. Our data demonstrate that the hRex-GFP constructs can be integrated into genome of human fibroblasts via lentiviral vectors. The construct is stable and can be passed to progeny cells. The expression of GFP is specific for pluripotent cells.
The hRex-GFP lentiviral vector reported here provides a convenient method to monitor the pluripotency in live cells in real time with single-cell resolution. Rex-1 is the direct target downstream of Oct-3/4 and has Oct-3/4-binding site in its promoter sequence [20,21]. Study of Oct-3/4 indicated that the expression of Oct-3/4 is not directly related to pluripotency. Stem cell differentiation was observed in both overexpression and suppression of Oct-3/4 [22]. The level of Oct-3/4 expressions may be associated with activation of different gene networks. Overexpression or suppression of Oct-3/4 may lead to activation of differentiation gene networks, while the appropriate level of Oct-3/4 associates with pluripotent networks including the Rex gene network. This could make Rex-1 expression a good marker of pluripotency [9]. In addition, the cellular function of Rex-1 is less studied than Oct-3/4, and how Rex-1 affects pluripotency in human cells is not clear. It was reported that mouse Rex-1 is dispensable for pluripotency in mouse ES cells [23]. Therefore, our study provides an additional marker to stem cell researchers, and a new tool for studying the Rex-1 cellular function in stem cells.
Because the hRex-GFP construct can be stably integrated into human genomes and is inherited by progeny cells, stable cell lines with this pluripotency reporter can be generated for tracking and studying transplanted cells in animals. This approach is not possible for transient transfection with mouse Rex-GFP plasmid [19], because changes in GFP expression may occur due to the degradation or loss of the plasmid during cell division. Our data show that BJ cells that carry this hRex-GFP construct do not express GFP, until they are reprogrammed to pluripotent stage. These results demonstrate that the insertion of hRex-GFP construct is stable and functional in descendant cells, a finding that has particular value in animal studies in which transplanted cells are migrated into various tissues, but typically only small numbers of cells can be recovered for analysis. With this hRex-GFP reporter, the pluripotency of just a single recovered cell can be monitored over long periods of time via the expression of GFP. In addition to stability, the integration site of the hRex-GFP can be used specifically to identify the origin of the cells that carried this construct. This pluripotency reporter construct will facilitate stem cell research by providing a convenient method to monitor changes of pluripotency in live cells in real time.
Footnotes
Acknowledgment
This work was supported by a grant from the National Multiple Sclerosis Society (NMSS PP1473).