
Abstract
We have shown that bone marrow (BM) from untreated advanced lung and breast cancer patients (LCP and BCP) have a reduced number of colony-forming unit fibroblasts (CFU-Fs) or mesenchymal stem cells (MSCs). Factors that regulate the proliferation and differentiation of CFU-F are produced by the patients’ BM microenvironment. We have now examined whether conditioned media (CM) from patients’ CFU-F-derived stromal cells also inhibits the colony-forming efficiency (CFE) of CFU-F in primary cultures from healthy volunteers (HV)-BM. Thus the number and proliferation potential of HV-CFU-F were also found to be decreased and similar to colony numbers and colony size of patients’ CFU-F. Stromal cells from both of these types of colonies appeared relatively larger and lacked the characteristic spindle morphology typically seen in healthy stromal cells. We developed an arbitrary mesenchymal stromal cell maturational index by taking three measures consisting of stromal cell surface area, longitudinal and horizontal axis. All stromal indices derived from HV-CFU-F grown in patients’ CM were similar to those from stromal elements derived from patients’ CFU-F. These indices were markedly higher than stromal indices typical of HV-CFU-F cultured in healthy CM or standard medium [α-medium plus 20% heat-inactivated fetal bovine serum (FBS)]. Patients’ CM had increased concentrations of the CFU-F inhibitor, GM-CSF, and low levels of bFGF and Dkk-1, strong promoters of self-renewal of MSCs, compared to the levels quantified in CM from HV-CFU-F. Moreover, the majority of patients’ MSCs were unresponsive in standard medium and healthy CM to give CFU-F, indicating that the majority of mesenchymal stromal cells from patients’ CFU-F are locked in maturational arrest. These results show that alterations of GM-CSF, bFGF, and Dkk-1 are associated with deficient cloning and maturation arrest of CFU-F. Defective autocrine and paracrine mechanisms may be involved in the BM microenvironments of LCP and BCP.
Introduction
T
The functional integrity of the BM microenvironment during solid tumor growth in in vitro and in vivo systems is not completely understood. Abnormalities in the function of CFU-F may be one of the possible mechanisms underlying metastatic processes [16 –19]. In previous studies, we have found that BM from untreated advanced lung cancer patients (LCP) and breast cancer patients (BCP), without prior irradiation and chemotherapy treatments and without bone and BM metastases, had defective CFU-F as reflected by their colony-forming efficiency (CFE) reduction and their deficient capacity to form confluent layers of stromal cells [20 –23]. We now hypothesize that CFU-F in LCP and BCP are under the regulation of a defective microenvironment. CFU-F in turn also produces defective stromal cells as part of the microenvironment that alter the proliferation and differentiation of other CFU-F in a paracrine and autocrine way. In these studies we established in vitro models to examine the function of LCP and BCP-CFU-F in their BM microenvironment. Regulatory factor activities from patients’ stromal cell conditioned media (CM) were measured by its ability to regulate CFE of CFU-F as well as the maturation of stromal cells in primary cultures from healthy volunteers (HV)-BM.
Materials and Methods
Patients
Bone marrow (BM) samples were obtained from 14 HV, 14 consecutive untreated patients with non-small cell (squamous cell) LC, Stages III A, III B, and IV, and 14 consecutive untreated patients with infiltrative ductal BC, Stages III and IV. All the BM aspirates were studied in patients without BM and bone metastases, before surgery, irradiation, and chemotherapy protocols. The International Union Against Cancer TNM classification system was used. Bone metastases were detected by X-ray and bone scintigraphy. HV were donors for allogeneic BM transplantation. LCP, BCP, and HV were age- (median age = 50, range = 30–67) and sex-matched. Informed consent was obtained from all individuals. Investigations were approved by the British and I. Iriarte Hospital Ethical Committees and were performed in accordance with the principles of the Declaration of Helsinki.
BM micrometastases
Bone marrow (BM) infiltration with neoplastic cells was detected by immunocytochemistry staining (Universal Dako LSAB System, Biotin–Streptavidin–Peroxidase complex/diaminobenzidine (DAB) chromogen, Carpinteria, CA) and analysis of cell morphology was done by the Pappenheim technique. BM aspirates were stained with monoclonal antibodies against epithelial membrane antigen (EMA; Dako), cytokeratin AE1–AE3 (Dako), cytokeratin 7 (CK7; Dako), and cytokeratin 20 (CK20; Dako). Patients’ BM were considered positive for micrometastases only if cells expressed EMA (breast cancer), cytokeratin AE1–AE3 (lung and breast cancers), CK7 (lung cancer), and CK20 (lung cancer) and if cells were morphologically malignant.
Collection and preparation of BM cells
Bone marrow (BM) samples were collected under local anesthesia from posterior iliac crest into heparinized saline without preservatives (25 units/mL, Gibco Life Technologies, Gaithersburg, MD). Aspirates were diluted ½ times with phosphate-buffered saline (PBS, pH = 7.5) and were layered on Ficoll-Hypaque (density = 1,075 g/cm3; Sigma, St. Louis, MO). After being centrifuged for 25 min at 340g, mononuclear cells (MNC) were harvested from the interface, washed twice with PBS, and resuspended in α-medium (Gibco) containing 100 IU/mL penicillin (Gibco), 100 µg/mL streptomycin (Gibco), and 25 µg/mL amphotericin B (Gibco). The cellular suspension was counted with 3% acetic acid solution (Sigma) and cell viability was determined by 0.04% Trypan blue (Sigma) dye exclusion.
Fibroblast colony-forming unit (CFU-F) assay
The 2 × 106 viable light density MNC, from 14 HV, 14 LCP, and 14 BCP, were placed in 25-cm2 tissue culture flasks (Corning Glass Works, NY) containing 10 mL of previously described supplemented α-medium (Gibco) plus 20% heat-inactivated fetal bovine serum (FBS) (cat number 16,000-044; Gibco). This supplemented medium with 20% FBS (standard medium) is known to be blastogenic for MSCs and fibroblast precursors because it contains optimal concentration of PDGF, bFGF, EGF, and TGF-β [24,25]. FBS from the same batch was used for all experiments. Cells were incubated in a 37°C, 5% CO2 humidified environment for 7 days. After this period, nonadherent cells were removed, the 7-day CM was harvested, and the medium was renewed. The primary cultures were incubated for an additional 7 days. At the end of this period, the 14-day CM was harvested and the adherent cells were washed twice with PBS, fixed with 100% methanol (Merck, 15 min), and stained with pure Giemsa (Sigma, St. Louis, MO) for 5 min. Clones of >50 cells were scored as CFU-F under a light microscope. The frequency of CFU-F is indicated by CFE defined as the ratio of the number of colonies to the number of cells seeded. For the evaluation of the CFE we read two flasks of each sample.
Morphological changes in these LCP, BCP, and HV adherent MNC cultures (6/14 samples of each group) were evaluated with Scion Image Software. Analysis of area, ellipse longitudinal/major and horizontal/minor axis values was carried out using two pictures obtained from two typical regions (two optical fields, 200×) of each CFU-F culture, evaluating 10 cells per photo. A total of 72 photos (24 per group) with a total of 720 cells were examined considering the duplicate cultures. The number of stromal adherent cells (majority fibroblasts, 85%–95%) per microscope optical field (100×) of CFU-F (stromal cell density, SCD) was also determined. For this purpose, 10 photos of different optical fields of each CFU-F culture were taken and analyzed with Photo Impact SE Software. A total of 360 photos (120 per group) were evaluated considering the duplicate cultures.
The fibroblastic nature of the mesenchymal stromal cells that composed the CFU-F cultures was demonstrated by immunocytochemistry using a monoclonal antibody against human β-subunit of prolyl-4-hydroxylase (Dako) that reacts only with fibroblasts and fibroblastic progenitor cells. Cell samples were fixed with 50% methanol, and sequentially incubated with biotin-labeled secondary antibody and streptavidine–peroxidase, following manufactures’ instructions (Dako, Kit of Biotin–Streptavidin–Peroxidase complex/DAB chromogen).
In parallel, we studied the presence of tumor cells in these CFU-F cultures from LCP and BCP by immunocytochemistry staining as previously described.
Preparation of CM from CFU-F cultures
CM (7 and 14 days) were obtained by centrifugation at 250g for 10 min and frozen at −20°C and −70°C until use.
Determination of the levels of Dkk-1, IL-1β, TGF-β1, PDGF-AB, GM-CSF, basic FGF, soluble receptors of TNF-α, EGF, IL-17, PGE2, IL-4, TNF-α, INF-α and INF-γ, IL-2, and IL-10 in the CM of CFU-F cultures at days 7 and 14
Soluble factors were quantified by ELISA in CM of CFU-F cultures from BM of 14 HV, 14 LCP, and 14 BCP. INF-α and Dkk-1 were measured by an ELISA Kit from Endogen Co. (Woburn, MA) and an ELISA Kit developed in our laboratory, respectively. The rest of the soluble factors were measured by specific ELISA Kits from R&D System (Minneapolis, MN). We used as control samples of supplemented α-medium with 20% of FBS after 7 days of incubation at 37°C and the levels of these soluble factors in these samples fell below assay detection limits. All samples and standards were carried out in triplicate. TGF-β1 and PGE2 were measured in CM frozen at −70°C and the rest of the factors were quantified in CM frozen at −20°C.
Mainly because of their hematopoietic origin PDGF-AB, IL-17, IL-4, INF-γ, and IL-2 were quantified only in CM at day 7.
Dkk-1 ELISA development kit
Microwell plates (Costar, catalog number 2592) were coated with 8 µg/mL PBS capture antibody (anti-human Dkk-1 antibody, goat IgG, R&D System, catalog number AF1096) and incubated overnight at room temperature.
Each well was aspirated and washed thrice with washing buffer (0.05% Tween 20 [USB, catalog number 20605] in PBS) and then blocked with reagent diluent (1% BSA [Sigma; catalog number A2153] in PBS) for 1 h at room temperature. The microplates were washed again and incubated with samples (CM, 7 and 14 days) or standards in reagent diluent (recombinant human Dkk-1 = rhDkk-1, R&D System, catalog number 1096-DK) (in triplicate). Standard curve of rhDkk-1 was prepared in reagent diluent, and the concentration range was between 16 and 0.250 ng/mL. The microplates were incubated 2 h at room temperature. After this period, the microplates were washed thrice with washing buffer and incubated with detection antibody (biotinylated goat anti-human Dkk-1 antibody, goat IgG, R&D System, catalog number BAF1096) for 2 h at room temperature. The working concentration of detection antibody was 50 ng/mL in reagent diluent. Then the microplates were aspirated and washed again. Substrate solution (streptavidin–peroxidase from Streptomyces avidinii [Sigma; catalog number S5512]) was added and the microplates were incubated for 20 min at room temperature. After this period, the microplates were washed again. Enzyme activity was revealed with 100 µL of substrate solution (8 µL H2O2–tetramethylbenzidine [3 mg, Sigma T8767] in 10 mL citrate buffer [0.1 M, pH = 5.5, Mallinckrodt]) added to each well and the microplates were incubated 20 min at room temperature. Finally, 50 µL of stop solution (1 N HCl, Merck) was added per well and the optical density of each well was determined immediately using a microplate reader set to 450 nm.
Evaluation of the effects of CM (100%, 7 and 14 days) from CFU-F cultures of LCP and BCP on HV-BM fibroblast colony formation
The 2 × 106 viable light density MNC from healthy BM were placed in 25-cm2 tissue culture flasks (Corning) containing 10 mL of standard medium or 10 mL of 100% CM from LCP, BCP, or HV-CFU-F cultures. Each CM from CFU-F cultures of 6 LCP, 6 BCP, and 6 HV harvested at days 7 and 14 was tested against six different healthy BM. Each culture was assayed in duplicate.
After 7 days nonadherent cells were removed, and each culture was incubated in the respective medium for additional 7 days. At the end of this period, the adherent cells were washed twice with PBS, fixed with 100% methanol, and stained with 100% Giemsa. Clones of >50 cells were scored as CFU-F under a light microscope.
Morphological changes in these HV adherent MNC cultures treated with standard medium or LCP, BCP, and HV-CM (7 and 14 days) were evaluated with Scion Image Software. Analysis of area, ellipse longitudinal and horizontal axis values was carried out using two pictures obtained from two typical regions (two optical fields, 200×) of each CFU-F culture, evaluating 10 cells per photo. A total of 456 photos (144 photos per CM treatment [LCP, BCP, and HV] and 24 photos of those HV cultures incubated in standard medium) with a total of 4,560 cells were evaluated considering the duplicate cultures. The number of stromal adherent cells (majority fibroblasts) per microscope optical field (100×) of CFU-F (SCD) was also determined. For this purpose, 10 photos of different optical fields of each CFU-F culture were taken and analyzed with Photo Impact SE Software. A total of 2,280 photos (240 per CM/healthy BM and 60 per treatment with standard medium) were examined considering the duplicate cultures.
Additionally, three of the healthy CM (days 7 and 14) of CFU-F cultures (at 20% and 100%) were tested in duplicate against 3 LCP and 3 BCP-MNC cultures for the purpose of evaluating its effects on CFE of patients’ CFU-F. During all the period of incubation, each culture was assayed in the respective medium.
Evaluation of the effects of CM (20%, 7 and 14 days) from CFU-F cultures of LCP and BCP on HV-BM fibroblast colony formation
The 2 × 106 viable light density MNC from healthy BM were placed in 25-cm2 tissue culture flasks containing 10 mL of standard medium or standard medium plus 20% CM from LCP, BCP, or HV-CFU-F cultures. Each CM from CFU-F cultures of 8 LCP, 7 BCP, and 6 HV harvested at days 7 and 14 was tested against six different healthy BM. Each culture was assayed in duplicate.
After 7 days nonadherent cells were removed, and each culture was incubated in α-medium plus 20% FBS for additional 7 days. At the end of this period, the adherent cells were washed twice with PBS, fixed with 100% methanol, and stained with 100% Giemsa. Clones of >50 cells were scored as CFU-F under a light microscope.
Statistics
Statistical analysis was performed using parametric and non-parametric tests as needed. Differences were considered statistically significant when P < 0.05.
Results
BM micrometastases
Non-evidence of BM infiltration with neoplastic cells was observed in any of the cancer patients studied. Moreover, neoplastic cells were not found in patients’ CFU-F cultures indicating that all the soluble factors were released by stromal and hematopoietic cells in the CM of these cultures (data not shown).
Effects of conditioned media from patients’ CFU-F cultures on healthy volunteers’ BM fibroblast colony formation
As we have previously described [22,23], these BM from untreated advanced LCP and BCP have significantly lower CFE of CFU-F than healthy BM when grown under standard culture conditions. CFE = # of CFU-F/2 × 106 MNC were (Mean ± SE); LCP = 5.36 ± 1.38 (n = 14)a; BCP = 5.86 ± 2.51 (n = 14)b; and HV = 38.33 ± 3.98 (n = 14)a,b (a,b P < 0.001; non-parametric Kruskal–Wallis test). The CFE in patients’ BM is approximately seven times less. We hypothesized that the patient’s microenvironmental factors may be inhibiting the self-renewel processes of MSCs, CFU-F, in vivo. Possible mechanisms may involve a direct effect on the CFU-F inhibiting its ability to proliferate and that patients’ CFU-F may also be defective and unresponsive to positive signals from the microenvironment in vivo. To explore these possibilities, we first tested whether CM from patients’ CFU-F-derived stromal cell colonies has inhibitory activities that decrease the CFE of HV-CFU-F. CM from healthy CFU-F-derived colonies stimulated the CFE of HV-CFU-F equal to standard medium, indicating that HV-CFU-F are equally responsive as shown in Table 1. In contrast, CM from healthy CFU-F-derived stromal cells could not reciprocate to further enhance the CFE of patients’ CFU-F beyond standard medium indicating that the patients’ CFU-F are unresponsive to optimal concentrations of growth factors found in both healthy CM or standard medium (Table 2), while CM from patients’ stromal cells markedly reduced the CFE of HV-CFU-F by 5-fold (Table 1). These results indicate that the patients’ stromal microenvironment does not produce enough levels of growth factors or increased levels of one or more inhibitory factors that can decrease the CFE of HV-CFU-F in vitro to the same extent as the patients’ CFU-F in vivo as shown in Tables 1 and 2.
E
Values expressed as Mean ± SE. Statistical analysis: non-parametric Mann–Whitney’s test. a to h = P < 0.0022.
Each culture was assayed in duplicate. Six CM were evaluated from each group, testing their activity at 100% in six different HV-bone marrow (BM). To quantify the colony-forming efficiency (CFE) = # of CFU-F/2 × 106 mononuclear cells (MNC) from HV-BM, the effects of each CM on six HV-BM were measured, and the X-value from each CM was used to obtain the final Mean-value.
E
Values expressed as Mean ± SE. Each culture was assayed in duplicate. Three HV-CM were tested against 3 LCP and 3BCP-bone marrow (BM). Mononuclear cells (MNC, 2 × 106) from LCP or BCP were incubated with standard medium, and with this same standard medium further enriched with 20% of a CM from HV-CFU-F. Also, these MNC were cultured in 100% of such CM. During all the period of incubation, each culture was assayed in the respective medium. To quantify the colony-forming efficiency (CFE) = # of CFU-F/2 × 106 MNC from patients’ BM, the effects of each HV-CM on 3 LCP-BM and 3 BCP-BM were measured, and the Mean-value from each CM was used to obtain the final Mean-value.
Furthermore, results indicate an inhibitory effect of these patients’ CM (20%) on the CFE of HV-CFU-F, irrespective of the day when the CM was harvested. A significant decrease in fibroblast colonies number was found when comparing values of colonies obtained when HV-MNC were cultured in patients’ CM with those cultured in standard medium or CM from healthy CFU-F-derived stromal cell colonies (20%, 7 and 14 days) (Fig. 1). As we described in Material and Methods in these assays, HV-MNC were treated with standard medium plus 20% of patients’ CM for the first 7 days (7 and 14 days) and the last 7 days with standard medium to evaluate if the inhibitor effect of patients’ CM was suppressed with the FBS (20%) contained in the standard medium. Thus, results showed that 20% of FBS did not normalize the CFE of HV-CFU-F cultured previously with patients’ CM. The effects of patients’ CM over HV colonies formation were independent of whether the patient’s CFU-F culture had a high or low number of fibroblast colonies and the clinical stage of the cancer (data not shown).

Evaluation of the effect of 20% conditioned media (CM) from lung and breast cancer patients (LCP and BCP)-colony-forming unit-fibroblast (CFU-F) cultures on healthy volunteers’ (HV) bone marrow (BM) fibroblast colony formation. Values expressed as Mean + SE. Mononuclear cells (MNC, 2 × 106) from HV-BM were treated the first 7 days with: α-medium + 20% FBS (standard medium); or standard medium plus 20% of CM from LCP, BCP, or HV-CFU-F cultures. CM from CFU-F cultures (LCP = 8, BCP = 7, and HV = 6) were harvested at days 7 and 14, and each one was tested against six different healthy BM. Finally, the last 7 days of the CFU-F assay, the MNC were cultured in standard medium. Each culture was assayed in duplicate. To quantify the colony-forming efficiency (CFE) = # of CFU-F/2 × 106 MNC from HV-BM, the effects of each CM on 6 HV-BM were measured, and the X-value from each CM was used to obtain the final Mean-value. Statistical analysis: parametric Student–Newman–Keuls multiple comparison’s test. a, b, c, and d = P < 0.001; e, f, g, and h = P < 0.05.
In addition to these patients’ CFU-F alterations we found that MSCs from 8 of 14 LCP and 10 of 14 BCP achieved confluence in primary cultures, and when these confluent cells were further induced to proliferate following four continuous subcultures, the growth diminished or stopped in 5 and 7 LCP and BCP cultures, respectively. In contrast, all HV (#14) cultures presented full confluent stromal layers in primary cultures and had the capacity to proliferate following four continuous subcultures. Additionally, the number of days that MSC took to achieve confluence in primary cultures were (Mean ± SE): HV = 20.39 ± 1.90 (#14)a,b; LCP = 36.18 ± 2.75 (#8)a; and BCP = 35.51 ± 2.98b (#10). Statistical analysis: non-parametric Mann–Whitney’s test, a P < 0.0014 and b0.0011.
Stromal cell density (SCD) patterns of healthy volunteers’ CFU-F-derived stromal cell colonies grown in lung cancer and breast cancer stromal cell-conditioned media
In these studies we measured and compared SCD in individual colonies as shown in Figures 2 and 3. HV-CFU-F were cultured with either standard medium, 7- and 14-day CM (100%) from patients’ and healthy stromal cell colonies. Similar SCD patterns are shown in Figure 2A, B, and C from standard medium, 7 and 14 days healthy stroma CM. In contrast, SCD of HV-CFU-F cultured with 7- or 14-day CM from LCP stroma, shown in Figure 2D and E; or 7- and 14-day CM from BCP stroma, shown in Figure 2F and G, indicate that there is a marked reduction of stromal cells growth in HV-CFU-F-derived colonies. The measures of the SCD per microscope optical field of fibroblast colonies are shown in Figure 3. About 90%–95% of these stromal cells were fibroblast-like cells with prolyl-4-hydroxylase positive, independent of the source of CM or MNC used (data not shown). The SCD for HV-CFU-F grown in standard medium or CM from healthy CFU-F-derived stromal cell colonies is more than two times than the SCD of HV-CFU-F grown in CM from LCP and BCP stromal cells. In agree with these data, similar SCD were found in patients’ CFU-F cultures. The number of stromal adherent cells/microscope optical field of each patient’s colony was decreased compared to the value observed in HV and the Mean ± SE values were: LCP = 209.60 ± 18.02a; BCP = 244.53 ± 16.64b; and HV = 565.27 ± 69. 73a,b (a P < 0.01 and b P < 0.05, non-parametric Dunn’s multiple comparisons test).
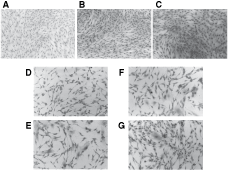
Distribution of adherent stromal cells from healthy bone marrow (BM) in colony-forming unit-fibroblast (CFU-F) cultures treated with: (

Evaluation of the effect of 100% conditioned media (CM) from lung and breast cancer patients’ (LCP and BCP) colony-forming unit-fibroblast (CFU-F) cultures on the number of stromal cells per microscope optical field (stroma cell density, SCD) in each CFU-F from healthy volunteers’ (HV) bone marrow (BM) cultures. Values expressed as Mean + SE. Mononuclear cells (MNC, 2 × 106) from HV-BM were treated with: α-medium + 20% FBS (standard medium) or 100% of CM from LCP, BCP, and HV-CFU-F cultures. CM from CFU-F cultures were harvested at days 7 and 14, and each one was tested against six different healthy BM. Six CM of each group were used. Each culture was assayed in duplicate. Statistical analysis: non-parametric Dunn’s multiple comparisons test. a, e, f = P < 0.001; b, c, d, h = P < 0.05 and g = P < 0.01.
Representative groupings of stromal elements from HV-CFU-F grown with standard medium, 7-day CM, and 14-day CM from healthy stromal cells, shown in Figure 4A, B, and C, respectively, have similar spindle-shaped morphology indicative of mature stromal elements. In contrast, stromal elements from HV-CFU-F cultured with 7- and 14-day stromal CM from LCP, shown in Figure 4D and E, appear relatively immature with a “blanket cell type” morphology. A similar pattern is also seen with those healthy CFU-F grown in BCP CM (Fig. 4F and G). Furthermore, similar type of stromal cell morphologies were found in patients’ CFU-F-derived stromal cells (Fig. 5).
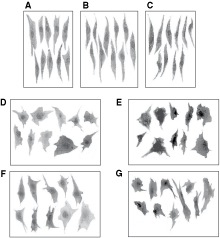
Pictures of 10 stromal cells in typical regions of colony-forming unit-fibroblast (CFU-F) cultures from healthy volunteers (HV)-bone marrow (BM) plated in different conditions: (

Pictures of 10 stromal cells in typical regions of colony-forming unit-fibroblast (CFU-F) cultures from (
We hypothesized that the relative size differences among HV individual stromal cells grown in healthy and patients’ CM or between HV and patients’ stromal cells may be used as a surrogate marker to predict individual stromal cell maturational arrest. Three different measures that included surface area, the longitudinal axis, and the horizontal axis were used to estimate the size of individual stromal cells (Fig. 6). Lower stromal index values were associated with mature stromal cells, while a high stromal index was associated with an inadequate maturation of stromal cells. All these three indices consistently showed that HV stromal cells cultured in standard medium (a) or with 7 (b) and 14 (c) days CM from healthy CFU-F (Fig. 6) had the lowest scores, indicating that they were relatively mature (spindle shape morphology, small cells), while stromal cells cultured in 7 and 14 days CM from LCP and BCP (Fig. 6D, E, F, G, respectively) had higher scores that were similar (“blanket cell type morphology,” large cells). These data suggest that the microenvironment from LCP and BCP produce inhibitory factors or not optimal concentration of growth factors that regulate the proliferation and differentiation of healthy CFU-F by rendering them unresponsive to growth factor signaling that keeps them in an inadequate maturation state. These last results obtained with the patients’ CM were in agreement with the high stromal index (area, longitudinal and horizontal axis) observed in the patients’ CFU-F cultures. The values (Mean ± SE) of the area were (mm2): LCP = 6.00 ± 1.20a; BCP = 3.82 ± 0.46b; and HV = 0.19 ± 0.02a,b (a P < 0.01 and b P < 0.05, non-parametric Dunn’s multiple comparisons test). The values (Mean ± SE) of the ellipse longitudinal axis were (mm): LCP = 4.12 ± 0.30a; BCP = 3.27 ± 0.23b; and HV = 1.05 ± 0.89a,b (a P < 0.01 and b P < 0.05, non-parametric Dunn’s multiple comparisons test). The values (Mean ± SE) of the ellipse horizontal axis were (mm): LCP = 1.75 ± 0.21a; BCP = 1.35 ± 0.09b; and HV = 0.23 ± 0.01a,b (a,b P < 0.01, non-parametric Dunn’s multiple comparisons test). The data showed that CFU-F-derived stromal cells from HV had lowest scores (spindle shape morphology, small cells with prolyl-4-hydroxylase positive, Fig. 7A) while the stromal cells from LCP and BCP had higher scores (blanket cell type morphology, large cells with prolyl-4-hydroxylase positive, Fig. 7B and C, respectively).
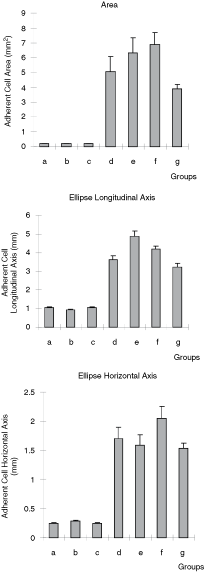
Cell area, ellipse longitudinal and horizontal axis of the 20 stroma cells in typical regions of each colony-forming unit-fibroblast (CFU-F) culture from healthy volunteers (HV)-bone marrow (BM) plated in different conditions: (

(
The BM microenvironment in LCP and BCP may keep CFU-F unresponsive to growth factors by unbalanced production of regulatory factors that promote and inhibit the proliferation and terminal maturation of stromal cells. There were no significant differences in the concentration (Mean ± SEM, pg/mL) of most growth factors in stromal CM from LCP, BCP, and HV-BM with the exception of three growth factors, GM-CSF, bFGF, and Dkk-1. GM-CSF concentrations from day 7 CM were 77.33 ± 33.64 and 98.18 ± 43.00 from LCP and BCP, respectively, and were markedly higher than healthy stroma ≤7.8. bFGF from day 7 CM were 6.60 ± 0.18 and 7.04 ± 0.88 from LCP and BCP, respectively, and were more than two times less than that of healthy stroma = 17.29 ± 2.25 (P < 0.05 and P < 0.01; non-parametric Dunn’s multiple comparisons test) while Dkk-1 from LCP = 2,015 ± 429 and BCP = 1,798 ± 469 was not different from healthy stroma = 1,602 ± 399. In contrast, 14-day conditioned media’s GM-CSF from patients was ≤7.8, and markedly reduced by >10 times the concentrations from the same patients’ 7-day CM, and similar to 7- and 14-day CM from healthy stroma. While 14-day conditioned media’s bFGF from LCP = 7.00 ± 0.39; and BCP = 7.83 ± 0.95 was not different from healthy stroma = 5.90 ± 0.25. Dkk-1 levels from 14-day CM from LCP = 4,276 ± 691 and BCP = 2,665 ± 611 were more than three times less than healthy stroma = 14,484 ± 4,034 (P < 0.0008, P < 0.0002; non-parametric Mann–Whitney’s test). These results indicate that concentration of Dkk-1 from 14-day CM from LCP stroma was more than two times greater than its 7-day CM; and 14-day CM from healthy stroma produced more than seven times greater than its 7-day (P < 0.01 and P < 0.0001, respectively, non-parametric Mann–Whitney’s test). Respect to the CM from BCP the increase of Dkk-1 levels between the days 7 and 14 was 18%. These data indicate that Dkk-1 production from CFU-F-derived stromal cells in cancer patients is markedly lower than healthy stroma by more than 4-fold.
There were no significant differences in the concentration of the evaluated inhibitory factors (PGE2, IL-4, TNF-α, INF-γ, INF-α, IL-2, and IL-10) in day 7 and day 14 CM from LCP, BCP, and healthy stroma.
Finally, studies were made to confirm if the increase of GM-CSF and the decrease of bFGF and Dkk-1 were the only factors responsible for the deficient cloning capacity of MSCs, CFU-F, from BM of LCP and BCP. For this purpose, we performed experiments by adding recombinant human bFGF (rhbFGF, 1–10 ng/mL; Sigma) and recombinant human Dkk-1 (rhDkk-1, 0.01–0.1 µg/mL; R&D system, Minneapolis, MN) to promote proliferation and anti-human GM-CSF antibody (different concentration starting on 0.01 µg/mL; R&D system) to neutralize inhibitory effects in standard medium used to culture CFU-F from patients’ BM. Results showed that adding growth factors bFGF, Dkk-1, and neutralizing anti-GM-CSF antibody did not alter the responsiveness of CFU-F from LCP and BCP (data not shown). These results suggest that the majority of CFU-F from BM of patients are defective and do not respond to regulatory signals as CFU-F from healthy BM.
Discussion
The stromal microenvironment of healthy BM produces an array of growth factors and components of the extracellular matrix that regulate the differentiation and terminal maturation of CFU-F. These mitogenic processes involve paracrine mechanisms between MSCs, CFU-F, and stromal elements of the microenvironment. The initial cell divisions involve limited self-renewal processes that ensure that at least one daughter cell remains in G0. The unique features of the mesenchymal system are that it also functions as an autocrine organ as it forms functionally mature stromal elements that promote further expansion of stromal elements within the same clone [24,26–28].
Ruling out neoplastic cells
Initial studies were essential to confirm that the BM microenvironment was not infiltrated with breast or lung cancer cells to eliminate the possibility that they both were involved to suppress patient’s fibroblast colonies formation. We did not observe any neoplastic infiltration in any of the cancer patients’ BM specimens studied. Moreover, neoplastic cells were not found in patients’ BM-CFU-F-extended stromal cell cultures, indicating that neoplastic progenitor cells were not present in the BM microenvironment of LCP and BCP.
Determining baseline estimates of CFU-F concentrations in patients’ bone marrow
It was also important for us to determine the ratio of CFU-F in patient’s BM in order to obtain a comparative baseline between both type of patients’ and healthy BM. It became apparent that the number of CFU-F and growth characteristics of stromal elements between LCP and BCP were similar and suggested that similar mechanisms may be involved. We hypothesized that patients’ microenvironments are functionally defective and disrupt the asymmetric self-renewal of MSC to fibroblast colony so that the majority of CFU-F progeny remain in G0, while the minor population of CFU-F progeny is partially responsive to regulatory factors. We thought that these defects were central to both patient populations and could account for both the low cloning efficiency of MSC, CFU-F, and the retarded maturation of their stromal progenies. Moreover, the defective stromal progenies were also hypothesized to produce regulatory factors that also maintain any secondary fibroblast colony generated by self-renewal in G0. Then these inhibitory activities produced by each colony should be detected and quantified in CM, biologically active, by inhibiting the growth of CFU-F from HV-BM.
Suppression of CFU-F from healthy volunteers’ bone marrow by conditioned media from patients’ CFU-F
CM from LCP and BCP induced morphological changes in HV stromal cells (majority fibroblasts), from small spindle-shaped to large polygonal-shaped. The block of maturational processes may also be involved in dedifferentiation or conversion of individual mature stromal elements to an immature state. We were able to measure individual representative stromal elements by their surface area and ellipse longitudinal and horizontal axis. All of these measures consistently correlated with stromal cells from both cancer patients having higher scores and healthy stromal cells having lower scores. These morphological changes of healthy stromal cells exposed to CM from cancer patients might be associated with an inappropriate response to a low level of bFGF (7 days CM) and suboptimal production of extracellular matrix components, which are essential to fibroblastic progenitor proliferation and confluence achievement [29,30]. Induction of mitosis by bFGF is accompanied by a persistent decrease in mean cellular volume and a decrease in collagen concentration [30]. Prockop et al. [13,28,31] described that the cells within the single cell-derived colonies are morphologically heterogeneous in that they contain both small, rapidly self-renewing cells, and larger, more slowly replicating cells. Moreover, cultures enriched for small cells have a greater potential to differentiate than cultures of the large cells [31]. These last observations are in agreement with our recent works where we have found a reduction of the osteogenic and adipogenic differentiation potential of patients’ MSCs compared to HV [32]. Moreover, the patients’ MSC cultures with osteoblastic/osteocytic cells had a scattered anarchic distribution as well as a poor mineralized matrix, while HV cultures showed homogeneous distribution of calcium as well as an adequate mineralization of the entire examined surface. So, these patients’ BM cultures had stromal cells with higher surface area, ellipse longitudinal and horizontal axis and less osteogenic and adipogenic differentiation capacity.
We also showed a significant increase in GM-CSF levels in patients’ stromal cells’ CM at day 7. Other studies [33,34] have also shown that GM-CSF exhibits a dose-dependent inhibition of healthy fibroblast colonies formation by interfering with the orientation of certain cytoskeletal structures. The increase in GM-CSF concentrations coupled with the relatively low concentrations of other growth factors might contribute to the deficient proliferation of CFU-F from cancer patients and with the inhibitory effect of patients’ CM on the CFE of HV-CFU-F.
Moreover, our analyses revealed that the 14-day CM from patients were not an effective mediator of HV fibroblast colonies formation probably because of the low concentration of Dkk-1. In relation with this observation, recent studies [31,35] suggest that Dkk-1 could hold the key to maintain stromal progenitor cells and MSC at an undifferentiated state during expansion. When early passage of these cells are plated or replated at low density, the cultures display a lag phase of 3–5 days, then a phase of rapid exponential growth, which is followed by a stationary phase without the cultures reaching confluence. Once the cells leave the lag phase and begin the early log phase, they synthesize and secrete considerable quantities of Dkk-1, an inhibitor of the canonical Wnt signaling pathway [35,36]. Moreover, the effect of Dkk-1 in medium supplemented with 10% FBS was biphasic during stromal cells/MSCs expansion. In high concentrations, as 0.5 µg/mL, Dkk-1 decrease the rate of proliferation of stromal cells/MSCs and reduce both the colony size and number while at lower concentrations, between 0.01 and 0.1 µg/mL, Dkk-1 increase proliferation of these stromal cells [35]. Therefore, the low Dkk-1 levels in patients’ CM (14 days) correlates with the low number of stromal cells per optical field in patients’ colonies and in HV-CFU-F-derived colonies cultured with patients’ CM.
The patients’ CM were inhibitory to healthy CFU-F, suggesting that the inhibitory activity in the patients’ microenvironment directly targets CFU-F progenitor cells.
Responsiveness of lung and breast cancer patients’ CFU-F to conditioned media from healthy CFU-F
Lung and breast cancer patients’ BM-CFU-F cultured with standard medium (20% of FBS) are thought to produce stromal cell progeny that can only undergo a limited number of cell divisions, cannot undergo terminal maturation, and remain in an inadequate maturation state. In these studies we tested whether the patients’ population of MSCs, CFU-F, can respond to mitogenic activity in CM from healthy BM. The candidate growth factors included PDGF, EGF, bFGF, TGF-β1, and Dkk-1 [28,29,35,37–45]. Optimal concentrations of IL-1, PDGF, bFGF, EGF, TGF-β1, and Dkk-1 in CM from healthy CFU-F-derived stromal cell colonies were capable of sustaining the proliferation of CFU-F from healthy bone marrow. This last observation is in agreement with the results of other authors, who have demonstrated an optimal concentration of IL-1, PDGF, bFGF, EGF, TGF-β1, Dkk-1, and IL-3 in CM from healthy BM cultures and from nonadherent hematopoietic MNC cultures, which were capable of sustaining proliferation of fibroblastic progenitors and differentiation to fibroblast by themselves or in the presence of <20% FBS [35,37,46–48]. In contrast, CFU-F from BM of LCP and BCP did not appear to respond to these healthy microenvironmental regulators.
As we described before PDGF, EGF, bFGF, and TGF-β1 production by BM-mesenchymal stromal cells is important as well as the expression of their receptors for the regulation of fibroblast colonies formation, confluence capacity of stromal cells, and plasticity of MSCs [45,49]. In preceding works we have found a reduction in the percentage of mesenchymal stromal cells expressing receptors type I, II, III of TGF-β, type I of bFGF and EGF, as well as a decreasing in their expression per stromal cell in the BM primary cultures from untreated advanced BCP and LCP compared to HV [50]. These deficient expressions of TGF-β, EGF, and bFGF receptors could be taken as other responsible factor for the deficient cloning efficiency of patients’ MSCs and the lack of answer to CM from healthy stroma.
In summary, the results presented in this article clearly show morphological and functional alterations in MSCs, CFU-F, from untreated advanced cancer patients. The possibility to correlate these alterations with future clinical evolution of these advanced LCP and BCP in the near future is of critical importance.
Footnotes
Acknowledgments
This work was supported by grants from the Agencia Nacional de Promoción Científica, Tecnológica y de Innovación Productiva, Argentina (PICT98, #05-03541), Fundación Roemmers (Subsidio 2005-2006), and CONICET (PIP2005). All the experiments comply with the current laws of Argentina and have ethics approval.