
Abstract
Marker and functional heterogeneity has been described for embryonic stem cells (ESCs). This property has been correlated with the presence of ESC subpopulations resembling pluripotent cell lineages of the embryo. The ability to efflux Hoechst (Ho) displayed by side population (SP) cells has proven valuable as a marker to identify multipotent stem cells from a variety of tissues. Here we report that cultures from different ESC lines consistently show an SP population that displays antigens of undifferentiated ESCs, distinct drug efflux properties, and an expression pattern of ABC transporters, inner cell mass (ICM), and epiblast genes, which distinguish it from the non-SP ESC fraction. This SP population contains pluripotent cells that differentiate into ectoderm, mesoderm, and endoderm in embryoid body and teratoma assays. Further, purified SP cells efficiently integrate into developing morulae and contribute to ICM. Under standard ESC culture conditions, SP and non-SP populations display ability to convert into each other; however, an equilibrium establishes between these fractions. Using protocols customized for SP ESCs, we report that cells with similar efflux properties can be identified in the ICM of peri-implanted blastocysts. Our results indicate that ESCs display heterogeneity for the SP marker, and the SP population of these cultures contains cells that phenotypically and functionally resemble efflux-active ICM cells of the peri-implanted embryo. Our observations suggest an involvement of the SP phenotype in ESC maintenance and early embryo development, and support the idea that ESCs are composed of distinct phenotypic and functional pluripotent subpopulations in dynamic equilibrium.
Introduction
E
The ability to actively efflux the fluorescent dye Hoechst 33342 (Ho) displayed by side population (SP) cells [17] has been exploited as a marker to identify and purify stem cells from a variety of tissues [18 –21]. SP cells can be identified by FACS as an unstained (Holow) cell population that displays sensitivity to the ABC transporter inhibitor Verapamil (VP) [17,18]. Tissue-derived SP fractions are enriched in primitive cells that differentiate into cell types characteristic of the tissue from which they originated [17 –20,22,23], indicating that the SP marker co-segregates with multipotent stem cells. Results from ABC KO mouse models suggest that the SP phenotype is controlled by multiple genes [24,25], and reflects an ability to translocate biomolecules, including cell metabolites and xenobiotics [26]. However, the precise function of the SP phenotype in stem cells remains to be elucidated.
Although significant attention has been devoted to the SP cells of adult tissues, little is known about the SP cells throughout embryo development. In the post-implanted mouse embryo, multipotent SP cells can be detected as early as day 8 post-coitum [23 –25]. Recently, cells with VP-sensitive ability to efflux Ho have been described for the ICM of the blastocyst [27], suggesting that SP cells emerge earlier in development and the SP phenotype may not be exclusive to multipotent stem cells.
Together with recent reports on marker and functional heterogeneity in ESCs, these observations led us to investigate whether ESCs contained SP cells, and if so, whether these SP cells displayed pluripotency and resembled cell types of the peri-implanted embryo. We found that cultures from multiple ESC lines consistently exhibited an ESC sub-population of Ho-effluxing cells that was almost completely blockable by VP, demonstrating that it represented SP cells. This SP population displayed antigens of undifferentiated ESCs, distinctive drug efflux properties, and characteristic expression pattern of ABC transporters, ICM, and epiblast genes, which distinguished it from the non-SP ESC fraction. In vitro and in vivo differentiation studies showed that this population contained cells that displayed pluripotency, and increased ability to both contribute to developing morulae and integrate into the ICM of blastocysts, consistent with the properties of ICM-like cells. Purified SP cells reconstituted ESC cultures, and an equilibrium established between the SP and non-SP fractions under ESC conditions, suggesting that an inherent program controlled this property. Last, using staining conditions customized for the SP cells of ESCs, we identified cells with similar drug efflux properties in the ICM of peri-implanted embryos. Together, our observations indicate that ESC display heterogeneity for the SP marker, and the SP population of these cultures constitutes a phenotypically and functionally distinct subpopulation that contributes to ESC maintenance and contains cells that resemble efflux-active ICM cells of peri-implanted blastocysts. Our observations demonstrate that the SP marker is not exclusive to multipotent stem cells but is also present in embryo-derived pluripotent cell types, and suggest an involvement of the SP phenotype in ESC maintenance and early embryo development. Last, our results support the idea that ESCs contain distinct subpopulations resembling pluripotent cell lineages of the embryo in a dynamic equilibrium.
Materials and Methods
ESC lines and culture maintenance
The mouse ESC lines AB.2.2 (gift from M. Matzuk and S. Songyang, Baylor College of Medicine), D3 (ATTC CRL-11632), R1/E (ATCC SCRC-1036), and 7AC5/EYFP (ATCC SRC 1033) were grown on gamma-irradiated mouse embryo fibroblasts (MEFs) and maintained at 37°C in 5% CO2. ESC medium consisted of 85% Dulbecco's modified essential medium (DMEM)-high glucose (GIBCO), 15% ES-screened FBS (Hyclone), 0.1 mM nonessential amino acids (GIBCO), 2 mM glutamine (GIBCO), 1 mM sodium pyruvate (GIBCO), 0.1 mM beta-mercaptoethanol (Sigma), and 1,000 U/mL leukemia inhibitory factor (LIF) (Chemicon). Cultures were fed daily and passaged with 0.05% trypsin–0.53 mM EDTA (Invitrogen) each 2–3 days, when they reached ∼60% of confluence.
Hoechst staining
SP staining experiments were performed as in [17] with minor modifications. Briefly, single-cell solutions produced by trypsinization of ESC cultures were centrifuged 8 min at 100g and rinsed once with DMEM containing 10% ES-screened FBS to remove traces of trypsin. Cell pellets were then resuspended at 106 cells per milliliter in prewarmed ESC medium containing Hoechst 33342 (Ho) (Sigma) at the indicated concentrations and incubated at 37°C for 90 min. SP controls were run in parallel under identical conditions but included both Hoechst and Verapamil (Sigma) at the indicated concentrations (Fig. 1). After the 37°C incubation step, samples were cooled on ice for 5 min and centrifuged at 100g for 8 min at 4°C. Hoechst-stained cells were resuspended in ice-cold ESC medium containing 2 μg/mL of Propidium Iodide (PI) (Sigma) and maintained on ice in the dark until FACS sorting, usually for no more than 30 min.

Verapamil (VP)-sensitive side population (SP) in embryonic stem cell (ESC) lines. Representative Hoechst 33342 FACS profiles for the mouse ESC lines R1/E (
Immunophenotyping of SP cells
For antibody staining, Hoechst-stained cells were resuspended in ice-cold DMEM medium containing 2% fetal bovine serum (FBS) at 107 cells per milliliter. Samples were kept on ice during the whole staining procedure. Primary antibodies were added at 1/50–1/100 dilutions for 30 min. Secondary antibodies were added at 1/500 dilution for 15 min. Samples were rinsed twice with ice-cold DMEM containing 2% FBS and centrifuged at 100g for 8 min at 4°C. Rinsed cells were then resuspended in ice-cold ESC medium containing 2 μg/mL of PI and analyzed by FACS. We used the following antibodies: mouse IgM anti-SSEA1 (Millipore), Alexa 488 anti-c-KIT (eBioscience), PE anti-c-KIT (eBioscience), Alexa 647 anti-CD9 (eBioscience), PE-anti-CD31 (eBioscience), FITC-anti-SCA1 (eBioscience), FITC anti-CD34 (eBioscience), FITC anti-CD150 (eBioscience), and FITC anti-CD45 (eBioscience). Secondary antibodies were: FITC goat anti-mouse IgM (Abcam), PE goat anti-mouse IgM (Abcam). Isotype controls included: FITC-IgM Isotype (BD Pharmingen), PE-IgM Isotype (Cederlane), PE-IgG Isotype (Cederlane), and FITC-IgG Isotype (Cederlane).
FACS analysis and sorting
Analysis and sorting of ESCs were performed on a triple-laser flow cytometer (MoFlow, Cytomation) as in [17,18]. Briefly, Hoechst-stained cells were excited at 350 nm and their emission analyzed at 405/30 nm (Hoechst blue) and 670/40 nm (Hoechst red). SP cells were identified as a Hoechst low (Ho low) cell fraction that displayed sensitivity to VP. In each experiment, the SP gate was set on the basis of the 50 μM VP control sample and included at least 95% of VP-sensitive Ho low cells. Dead cells were excluded from the analysis. For RT-PCR studies, cells were directly sorted into centrifuge tubes containing RNA lysis buffer. For cell growth and differentiation studies, cells were sorted into 1.5 mL plastic centrifuge tubes containing ESC medium containing 20% ES-FBS and kept on ice. An aliquot of the collected sample was removed at the end of the sort and reanalyzed to establish purity. Only SP samples displaying at least 95% of purity were included in our studies. Analysis of collected FACS data was performed using FlowJo software (Tree Star Inc., Or, USA,
Ho and Rho co-efflux studies in ESCs
Simultaneous efflux of Ho and Rhodamine was performed as in [24] and [22] with minor modifications. Briefly, single ESCs were resuspended in warm ESC medium at a concentration of 106 cells/mL and incubated with 0.1 μg/mL for Rhodamine 123 (Sigma) without Hoechst for 30 min at 37°C (accumulation phase). Samples were then cooled on ice for 5 min, centrifuged at 100g for 8 min at 4°C, and rinsed with cold ESC medium. These pellets were resuspended in warm ESC medium at a concentration of 106 cells/mL, incubated with Hoechst for 90 min (efflux phase), and processed immediately for FACS analysis, as indicated earlier. Excitation and emission of Rhodamine were carried out at 488 and 530 nm, respectively.
Embryoid body and teratoma formation assays
For differentiation assays, sorted Hoechst-stained ESCs were spun down at 100g for 8 min and resuspended in differentiation medium containing 90% DMEM-high glucose (GIBCO), 10% FBS (Hyclone), 0.1 mM nonessential amino acids (GIBCO), 2 mM glutamine (GIBCO), and 50 μg/mL penicillin and streptomycin (GIBCO). For EB studies, cells were transferred into low attachment plates (Costar 2.2) and cultured in suspension for the indicated time as in [28]. For teratoma assays, single-cell suspensions (1.5 million cells) were directly injected in the hind limb of NOD/SCID mice. Animals were checked for the presence of palpable tumors in their legs once per week. Mice bearing tumors were anesthetized and sacrificed, and their tumors removed. Tumor samples were rinsed in PBS+/+ and fixed in 4% formalin, after which they were embedded in paraffin. Five micrometer serial sections were then processed for hematoxilin–eosin staining.
Indirect immunofluorescence microscopy
Processing of ESC cultures for immunofluorescence studies was performed as in [29] with minor modifications. Briefly, 2-day-old ESC cultures were rinsed three times with PBS−/− (GIBCO) and fixed in cold 2% paraformaldehyde (Sigma) PBS−/− (GIBCO) for 20 min. Fixed samples were then permeabilized with 0.2% Triton X-100 (Sigma) PBS+/+ (GIBCO) and rinsed three times with 0.1% Tween 20 (Sigma) PBS+/+. The antibody blocking solution consisted of 5% FBS (Hyclone) in 0.1% Tween 20 (Sigma) PBS+/+. Primary and secondary antibodies were used at 1/100 and 1/500 dilutions in blocking solution, respectively. Antibody-stained samples were incubated for 5 min with a nuclei-staining solution containing 50 μg/mL DAPI in PBS+/+, after which they were rinsed twice with PBS+/+. Samples were kept for 4 h in the dark at 4°C before immunofluorescence microcopy analysis.
PCR assays
RNA isolation was performed by using the RNAqueous kit (Ambion). We used Superscript II (Invitrogen) and recombinant Taq (Invitrogen) for RT and PCR, respectively, as recommended by the manufacturer's instructions. Realtime QPCR was performed on the ABI Prism 7500 Sequence Detection System (ABI, Foster City, CA). Amplification was performed using the Sybr Green Master Mix (ABI). The “ΔΔCt method” was used to quantify gene expression levels in the SP and non-SP populations. Gapdh was used as an internal control to normalize relative amounts of transcripts. Relative gene expression values were plotted as the average ± standard error of at least three independent experiments. We used the following sets of primers, some of which have been previously characterized in ESCs [6,9,30,31]: Oct4: 5′tctggagaccatgtttctgaagtg3′, 5′gaaccatactcgaaccacatcctt3′; Nanog: 5′agggtctgctactgagatgctctg3′, 5′caaccactggtttttctgccaccg3′; Sox2: 5′aagaaaggagagaagtttggagcc3′, 5′gagatctggcggagaatagttgg3′; Rex1: 5′cctgacggatacctagagtgcatc3′, 5′tgctccacttcctccaagctt3′; Pecam1: 5′tggtcatcgccaccttaatag3′, 5′cagaaatcttctcgctgttgg3′; Tbx3: 5′atctgccagtgcactttgttaga3′, 5′tgttcttcagccccgacttccatac3′; Gdf3: 5′cagccttatcaacggcttctg3′; 5′aggttggaacctgtgaggtga3′; Fgf4: 5′gcccaacaactacaacgccta3′; 5′ccttcatggtaggcgacactc3′; Blimp1: 5′agcatgacctgacattgacacc3′; 5′ctcaacactctcatgtaagaggc3′; Stella: 5′taaccctaaaccccggtgtca3′, 5′tccctccggattttttcactct3′; Fgf5:5′gctgtgtctcaggggattgt3′, 5′cactctcggcctgtcttttc3′; Nodal: 5′tttgagccaagaagaggatctggt3′, 5′ggtggaaaatgtcaatggtgagtg3′; Eomes: 5′cctggtggtgttttgttgtg3′, 5′tttaatagcaccgggcactc3′; Uvo: 5′catcagtgtgctcacctctgg3′, 5′ttgaagacaggagcgttgtca3′; Bry: 5′atgatcaccagccactgcttt3′, 5′ttggcatcaaggaaggcttta3′; Gata4: 5′ccccacaaggctatgcatct3′, 5′tccgagcaggaatttgaagag3′; Gapdh: 5′atgacatcaagaaggtggtg3′, 5′cataccaggaaatgagcttg3′; Abcc1: 5′ctactccatggctgtgtccat3′, 5′atcatggagtccactgtgtcc3′; Abcc2: 5′ctggtctcaaccaccctgtaa3′, 5′gagctatctccccagctcagt3′; Abcc3: 5′gaagttcaaggactcccgcat3′, 5′cactctgcctgatccctttca3′; Abcc4: 5′caccatgacacttcccctaga3′; 5′actagctggggagagcttttg3′; Abcb1b: 5′tatgccagagcttgcaggta3′, 5′gaaccggaaacaagcagca3′; Abcg2: 5′acctggccttaatgctattctgg3′; 5′gcaggttgaggtgctccatttat3′; Beta Tubulin 3: 5′tcactgtgcctgaacttacc3′, 5′ggaacatagccgtaaactgc3′; Msx3: 5′gagtctctgctggaggctga3′, 5′gtattgcttctggtgaaacttgc3′; Flk1: 5′ccccatttcaatggagaaaca3′, 5′gcatgtcaatgtctgcatggt3′; Col IV: 5′gaagtcatggctttcctgtgg3′, 5′acgaaaaccaagtgcgcttta3′; Afp: 5′gttttctgagggatgaaacctatg3′, 5′gaagctcttgtttcatggtctgta3′; Actin: 5′gaatcctgtggcatccat3′, 5′gacagcactgtgttggca3′.
Alkaline phosphatase assay
For analysis of alkaline phosphates activity, we used the quantitative alkaline phosphatase ES characterization kit (Millipore/Chemicon). SP and non-SP fractions were purified and counted by FACS. Identical numbers of sorted cells were collected in ES medium. Samples were then immediately processed for alkaline enzymatic activity as recommended by the manufacturer instructions. This enzymatic activity was measured on a spectrophotometer by reading amount of product produced after catalytic colorimetric reaction.
Efflux studies in blastocysts
Briefly, 3.5 dpc embryos were incubated in M2 medium containing 0.1 μg/mL of Rho without Ho for 30 min at 37°C (accumulation phase), like in ESC studies. They were then rinsed thrice in drops of ice-cold M2 medium, and immediately transferred to M2 medium containing 4 μg/mL of Ho, like in ESCs, for 30 min at 37°C (efflux phase). After this incubation, the embryos were rinsed thrice in ice-cold M2, and immediately analyzed under UV fluorescence microscope. Efflux of Rho and Ho was visualized using FITC and DAPI filters, respectively.
Diploid aggregation studies
Diploid aggregation studies were performed by using the “darning needle technique” [32] with minor modifications. Morula/eight-cell stage embryos were flushed from oviducts of C57Bl/6J;129S6/SvEv hybrid females, placed in microwells containing M16 medium (Sigma), antibiotics, and NEAA, and kept at 37°C in the incubator until sorting of the ESCs was completed. FACS-purified ESCs were collected in ESC medium containing antibiotics. Sorted ESC samples were then pelleted down and resuspended in fresh ESC medium containing antibiotics. ESCs were allowed to form clumps for 2–3 h at 37°C before aggregation. Selected ESC clumps were then allowed to aggregate with the morulae overnight. The next day, aggregates were analyzed for the presence of YFP+ cells under a fluorescence microscope, and further processed for confocal microscopy studies.
Confocal microscopy studies of chimeric embryos
Embryos containing YFP+ cells were fixed in 4% PFA overnight and permeabilized with 0.2% Triton X-100 (Sigma) PBS+/+ (GIBCO) before staining. The antibody blocking solution consisted of 5% FBS (Hyclone) in 0.1% Tween 20 (Sigma) PBS+/+. Primary and secondary antibodies were used at 1/100 and 1/500 dilutions in blocking solution, respectively. We used the following primary antibodies: rabbit anti-YFP (Clontech) and rat anti-Oct4/3 (Millipore/Chemicon). Secondary antibodies included goat anti-rabbit IgG-FITC (Cederlane) and donkey anti-rat IgG-Cy5 (Cederlane). Samples were analyzed by confocal laser microscopy in a Zeiss Axiovert 100M microscope. Dual-fluorochrome images were collected by using the LSM 510 acquisition software (Zeiss,
Results
A VP-sensitive SP population in multiple ESC lines
SP cells actively efflux Hoechst 33342 (Ho) and can be identified as a Holow cell population that displays sensitivity to the ABC transporter inhibitor VP [17,18]. ESC-derived cells exhibiting weak Ho staining have been previously described [33]. However, the identity of these cells and their sensitivity to VP were not characterized. Here we asked whether ESCs contained SP cells, and if so, we aimed at characterizing their phenotype and biological properties. Based on the original method for detecting SP cells [17], we set up VP and Ho dose–response experiments for commonly used ESC lines, including R1, D3, and Ab2.2. We found that under standard culture conditions, all the analyzed ESC lines consistently exhibited a small subpopulation of cells that effluxed Ho in a dose- and VP-dependent manner (Fig. 1). Concentrations of Ho between 5 and 4 μg/mL produced a Holow cell population that almost completely receded in the presence of 50 μM VP (Fig. 1), indicating that this property involved an active process that was blockable by VP. Such a population was absent in feeder cultures (Supplementary Fig. 1; Supplementary materials are available online at
The SP population displays antigenic markers of undifferentiated ESCs and a distinctive pattern of early development genes
We then aimed at characterizing the antigenic phenotype of these SP cells by first asking whether they expressed markers of undifferentiated ESCs. Thus, we investigated the expression of SSEA1, Pecam1, c-KIT, and CD9, as undifferentiated ESCs robustly express these markers [6,7,34 –36]. FACS analysis of Ho-stained ESCs revealed that almost the entire SP fraction expressed these antigens (Fig. 2A–2D), suggesting that these SP cells represented undifferentiated ESCs. Because hematopoietic and other tissue-derived stem cells have been shown to display SP phenotype [17,18], we also investigated the expression of hematopoietic and adult stem cell antigens. However, neither the SP nor the non-SP populations raised antigenic signal for the pan-specific stem cell markers, CD34 and Sca1, or the hematopoietic markers, CD150 and CD45 (Fig. 2E–2H, and Supplementary Fig. 2), indicating that both fractions contained a great proportion of undifferentiated ESCs.
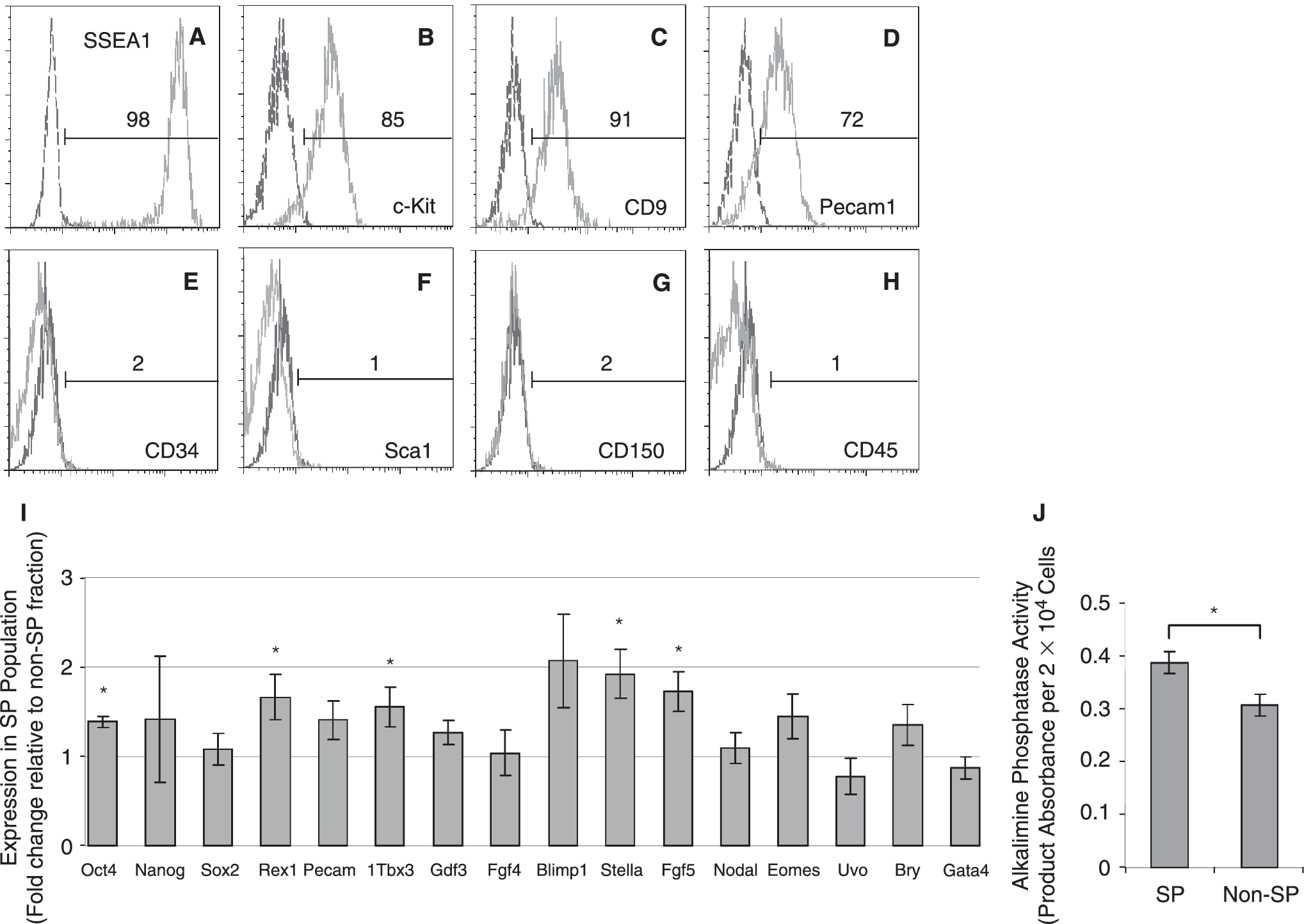
The side population (SP) displays characteristic markers of undifferentiated embryonic stem cells (ESCs) and distinct expression pattern of early development genes. (
Nonhomogeneous expression of early development genes has been described for ESCs grown under standard conditions, and this property has been correlated with the presence of ESC populations resembling cell lineages of the embryo [6 –13]. Thus, we analyzed the expression of representative gene markers of early development in the SP and non-SP populations of ESCs. As shown in Fig. 2I, QRT-PCR studies indicated that the SP population expressed well-known ICM genes at levels comparable to or higher than those of the non-SP fraction. These included Oct4, Nanog, Rex1, and Sox2, which have been described as key regulators of pluripotency networks, as well as Pecam1, Tbx3, Gdf3, and Fgf4 (Fig. 2). Further, the SP population consistently displayed increased levels of Oct4, Rex1, Tbx3, and Stella, consistent with the great percentage of SSEA1+ and Pecam1+ cells in this fraction, which have been shown to resemble ICM-like cells [6,9,13]. However, the SP population also expressed moderate levels of Fgf5, a typical epiblast marker, which has been described up-regulated in pluripotent epiblastic stem cell (EpiSc) lines established from the post-implanted embryo [37,38], as well as in the so-called early primitive ectoderm-like (EPL) cells, which have been derived from ESCs under controlled differentiation conditions [39]. Together with the presence of a small fraction of SSEA1+ Pecam1- cells in the SP population (Fig. 2), this observation suggested that the SP population also contained epiblast-like cell types [6,9,13]. Analysis of markers typically up-regulated in EpiSc, such as Nodal, Eomes, and Bry [37,38], and in EPLs such as Uvo [39], indicated no significant differences between the SP and non-SP populations, suggesting that early epiblast-like cell types rather accounted for the expression of Fgf5 in the SP fraction. Biochemical analysis of protein extracts derived from FACS-purified SP and non-SP populations revealed that the SP fraction exhibited increased alkaline phosphatase activity (Fig. 2J), which is absent in EpiSc [37,38]. Together these results indicated that the SP population displayed a distinct pattern of pluripotency and early development markers that distinguished it from the non-SP fraction, and resembled ICM and early epiblast lineages of the peri-implanted embryo.
To experimentally address this hypothesis, we then performed in vitro and in vivo functional assays.
Reversibility of SP and non-SP ESC populations
Recent studies [9,13] suggest that under standard ESC culture conditions, ESCs contain pluripotent subpopulations in dynamic equilibrium, each with a different combination of developmental regulated genes spanning from ICM to epiblast-like cell phenotypes. However, while ICM and early epiblast-like cell populations display ability to convert into each other, late epiblast cell-like subpopulations such as EpiSc cannot do so under these conditions [9,13]. Thus, we examined the in vitro cell growth properties of the SP and non-SP populations under ESC conditions by culturing them separately. We found that sorted SP cells gave rise to cultures of tightly compact colonies that could be expanded by single-cell dissociation methods and retained robust expression of SSEA1 (Fig. 3A–3E). Despite the high purity of the starting population, SP-derived cultures contained both SP and non-SP cells (Fig. 3F). Interestingly, the frequency of SP cells in the SP-derived cultures always dropped and reached a steady level resembling that of unsorted cultures after 5 days (Fig. 3F), indicating that the SP cells readily converted into non-SP cells. Antigenic analysis of the SP and non-SP fractions of these cultures revealed that they both expressed SSEA1, c-KIT, CD9, and Pecam1 (Fig. 3G–3J), indicating that the SP cells reconstituted undifferentiated ESCs. A similar phenomenon was observed for the non-SP-derived cultures, which also contained both SP and non-SP populations; however, we noticed that the appearance of SP cells was slower, as we were able to detect these cells after the third passage (Fig. 4A–4E). To rule out the possibility of cross-contamination between SP and non-SP fractions during cell sorting, we also analyzed the presence of SP and non-SP populations in single-cell-derived cultures. Once more, cultures derived from either single SP or non-SP cells consistently showed SP and non-SP populations (Fig. 4G and 4H), and after three passages, their proportions reached steady levels comparable to the ones of unsorted cultures. Together, these data indicated that the SP and non-SP cells of ESC cultures displayed ability to convert into each other, and an equilibrium consistently established between these two fractions. These results closely resembled the properties of recently described ICM- and early epiblast-like ESC subpopulations [9,13], and together with our marker data, it suggested that the SP fraction was composed of similar cell types, and an intrinsic program [13] controlled the equilibrium between SP and non-SP populations.
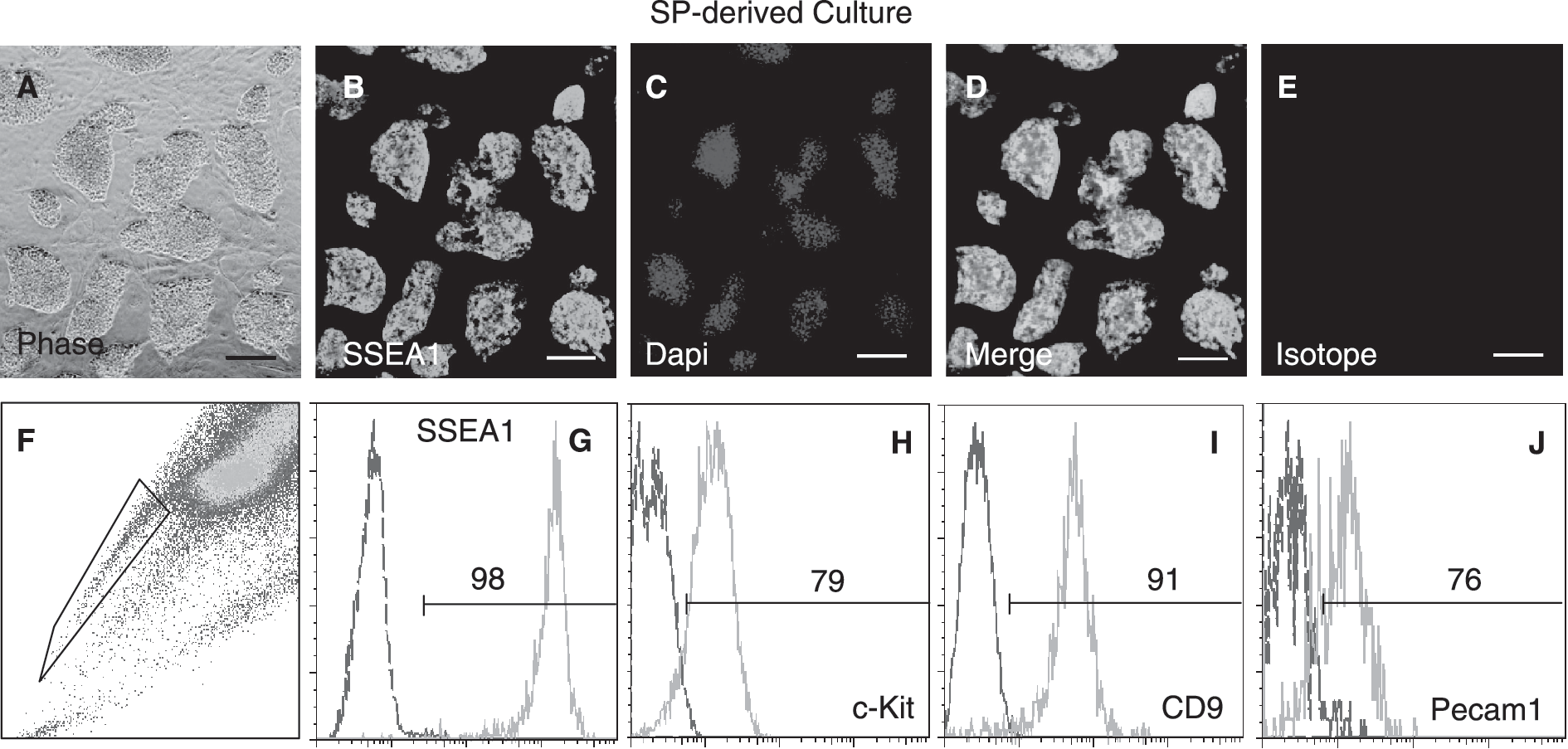
Purified side population (SP) cells can be expanded in culture under standard embryonic stem cell (ESC) conditions producing SP and non-SP cells that express markers of undifferentiated ESCs. (
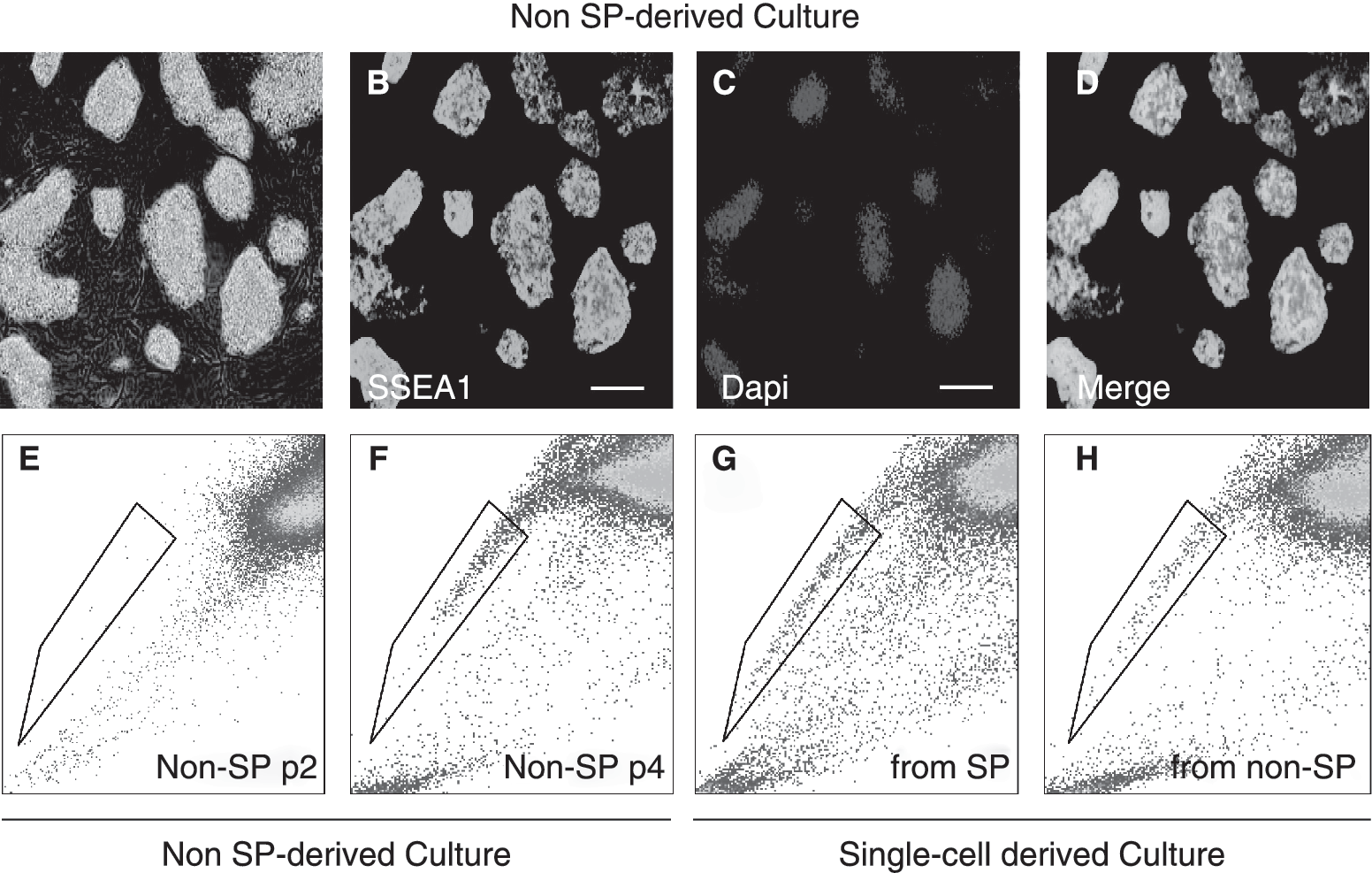
Reversibility between side population (SP) and non-SP embryonic stem cell (ESC) populations. (
The SP population contains pluripotent cells that contribute to ICM
The pluripotency markers and in vitro cell growth properties shown by the SP population suggested that it could contain pluripotent cells. Thus, we investigated the differentiation potential of the SP and non-SP populations by in vitro and in vivo approaches. FACS-purified SP and non-SP cells grown in suspension in the absence of LIF-generated robust embryoid bodies (EBs) (Fig. 5A) that expressed characteristic markers of ectoderm, mesoderm, and endoderm (Fig. 5B and Supplementary Fig. 3). These included: beta tubulin-3, Msx3, and GFAP (for ectoderm); Bry, and Flk1 (for mesoderm); and ColIV, AFP, and Gata4 (for endoderm). Although both SP and non-SP fractions displayed ability to differentiate into the same derivatives, SP-derived EBs expressed increased levels of transcripts for most of the analyzed markers (Fig. 5B), indicating robust in vitro differentiation potential for the SP population.
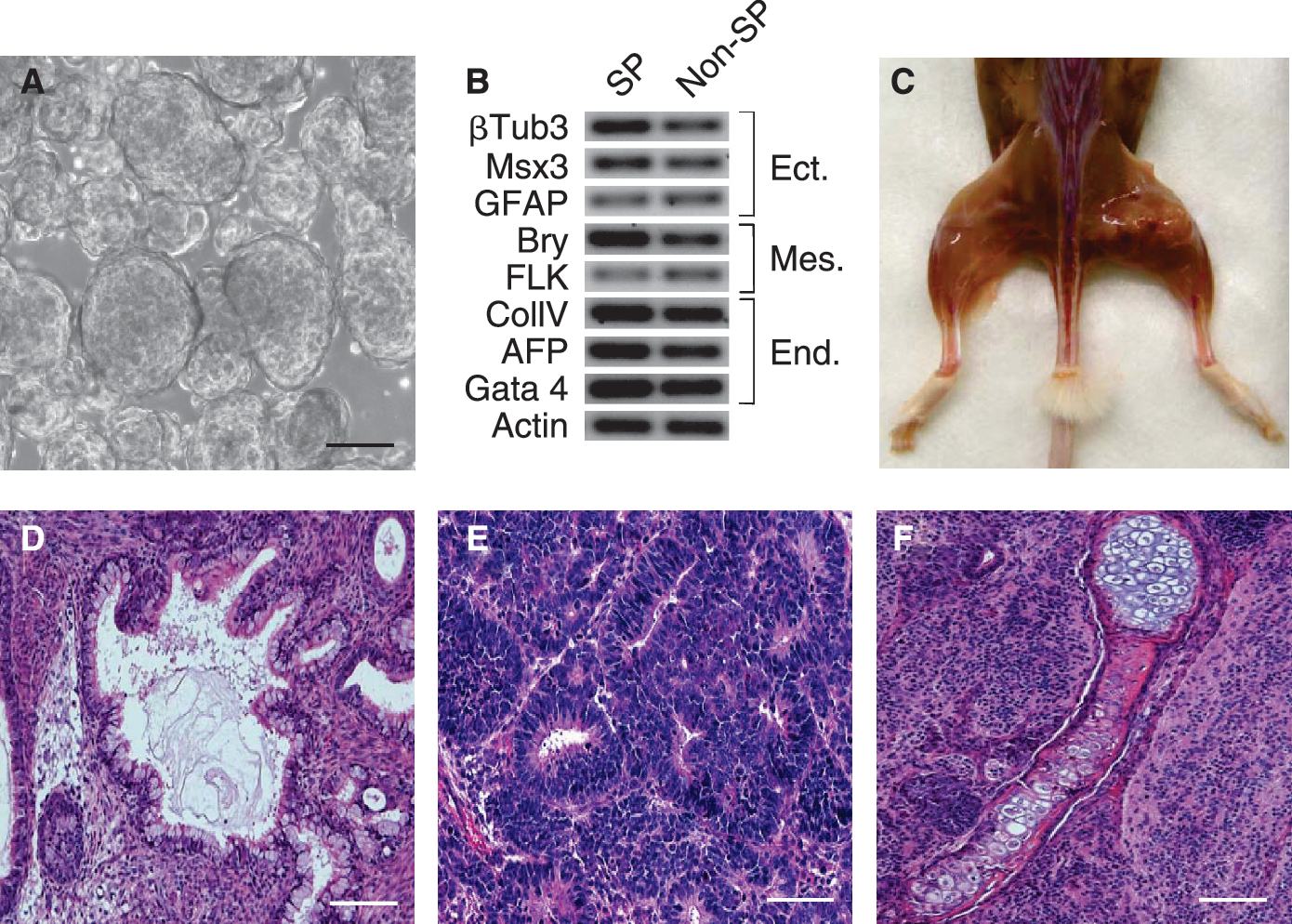
Purified side population (SP) cells display in vitro and in vivo pluripotency. (
The differentiation properties of the SP and non-SP fractions were also investigated in teratoma assays. Results from these studies showed that immunodeficient mice transplanted with either SP or non-SP cells exhibited encapsulated tumors at the sites of injection (Fig. 5C). The histology of these tumors confirmed that they represented teratomas, as they were composed of multiple semi-organized tissues of ectodermic, endodermic, and mesodermic origin, which included glandular, neural, cartilage and bone (Fig. 5D–5F and Supplementary Fig. 3). These results demonstrated that similar to the non-SP fraction, the SP population contained pluripotent cells with ability to differentiate into tissues of the three embryo germ layers. However, SP-derived teratomas developed faster, indicating increased teratoma formation potential for the SP population.
The molecular profile, in vitro cell growth properties, and differentiation potential of the SP population indicated that it contained pluripotent cell types reminiscent of the peri-implanted embryo, as they resembled ICM and early epiblast cells [6,9,13]. To address this hypothesis and further assess the extent of pluripotency of the SP and non-SP populations in vivo, we analyzed their ability to contribute to chimera formation in ES-morula aggregation assays, as pluripotent cell types of the post-implanted embryo, such as EpiSc, do not contribute to developing morula but remain either outside the blastocyst or within the blastocoele [37,38]. In order to track the contribution of the SP and non-SP cells into the aggregates, we used the ESC line 7AC5, which constitutively expresses YFP, and like other ESC lines displayed a characteristic SP population (Supplementary Fig. 3). Fluorescence microscopy analysis of the aggregates indicated that purified SP and non-SP cells incorporated into developing morulae (Fig. 6A–6C). However, SP cells did it with increased ability as 51% (n = 41) of the SP aggregates incorporated YFP+ cells (vs. 12% for non-SP cells (n = 43) and 34% for unstained ESCs (n = 35)). Further, confocal microscopy analysis of these aggregates at the blastocyst stage revealed that the SP cells contributed to the ICM, as determined by colocalization of YFP and Oct4 proteins (Fig. 6D–6I). Comparative studies showed that the SP cells contributed more efficiently to ICM than the non-SP cells, as SP-derived aggregates produced a greater proportion of blastocysts with YFP+ ICM cells than the non-SP ones (Supplementary Fig. 5). These results indicated that the SP population was enriched in cells that functionally resembled ICM-like cells [6,9], as different from pluripotent cells of the post-implanted embryo, they were compatible with the ICM of the blastocyst, and thus able to contribute to ICM. Together with the marker profile and in vitro cell growth properties displayed by the SP population, these observations indicated phenotypic and functional differences between the SP and non-SP ESC fractions.

Purified side population (SP) cells incorporate into developing morulae and contribute to the inner cell mass (ICM) of blastocysts. Aggregation of YFP+ SP cells with mouse embryos at the 8 cell/morula stage. (
SP ESCs and ICM cells share similar drug efflux properties
Recently, Ho- and Rho-effluxing cells have been described for the ICM of blastocysts, and expression of Mdr1/Abcb1b and Bcrp1/Abcg2 has been reported for these cells [27,40]. This led us to investigate whether the SP and non-SP fractions of ESCs exhibited differential ability to efflux these molecules and differentially expressed characteristic ABC transporters. As shown in Fig. 7A, analysis of ESC samples co-stained with Ho and Rho revealed that the SP population efficiently effluxed Rho, as the SP fraction displayed a fluorescence peak of lower intensity than the one produced by the non-SP population. QRT-PCR analysis of the SP and non-SP populations also revealed differential expression of ABC transporters, as the SP population expressed higher levels of Mrp3/Abcc3, Mdr1/Abcb1b, and Bcrp1/Abcg2 (Fig. 7B). Interestingly, Abcc3 has not been associated with SP phenotypes in mouse. It is thus possible that the combined activities of Abcc3, Abcb1b, and Abcg2 may be specific for the SP phenotype of mESCs. These results indicated that a distinct expression pattern of ABC transporters and an ability to efflux molecules also distinguished the SP and non-SP ESC fractions. More importantly, our observations suggested that this ability to co-efflux Ho and Rho would also be present in ICM cells.

Side population (SP) embryonic stem cells (ESCs) and inner cell mass (ICM) cells share similar drug efflux properties. (
To address this hypothesis, we also investigated whether the staining conditions we used for the SP ESCs would allow us to identify cells with similar Ho and Rho efflux properties in the ICM of peri-implanted blastocysts. Although efflux of these dyes has been described for ICM cells [27,40], whether these cells display ability to co-efflux these molecules, like the SP ESCs, is unknown. As shown in Fig. 7C–7J, alive blastocysts simultaneously stained with Ho and Rho displayed heterogeneous and overlapping patterns of Ho and Rho efflux-active cells that localized to the ICM. These results demonstrated that the SP population of ESC cultures also shared Ho and Rho efflux properties with ICM cells, further suggesting that the SP population contained cells that resembled ICM cells of the peri-implanted blastocyst.
Discussion
Recently, marker and functional heterogeneity has been described for ESCs, and this property has been correlated with the presence of ESC subpopulations resembling pluripotent cell lineages of the embryo. Here we investigated whether ESCs displayed SP phenotype, a marker which has been traditionally associated with adult stem cells. We found that under standard ESC culture conditions, different ESC lines consistently showed a distinct pluripotent SP population that displayed antigens of undifferentiated ESCs, and molecular, and functional properties, which distinguished it from the non-SP ESC fraction, suggesting it contained ICM- and early epiblast-like cells. Using staining conditions for SP ESCs, we found that cells with similar drug efflux properties could be identified in the ICM of peri-implanted embryos. Our results indicate that ESCs display heterogeneity for the SP marker, and the SP population of these cultures contains pluripotent cells that phenotypically and functionally resemble efflux-active cells of the ICM of the peri-implanted blastocyst. Our observations suggest an involvement of the SP phenotype in ESC maintenance and early embryo development.
Our results demonstrate that the SP marker is not exclusive to multipotent stem cells but is also present in cell types that resemble pluripotent lineages of the peri-implanted embryo. For instance, we found that the SP population contained a great proportion of Pecam1+ SSEA1+ cells that have been shown to resemble ICM-like cells in ESC cultures [6,9,13]. Consistent with this observation, we found that the SP fraction expressed several well-known ICM markers including Oct4, Nanog, Rex1, Sox2, Pecam1, Gdf3, Tbx3, Fgf4, and Stella. However, the SP population also contained a small percentage of Pecam1- SSEA1+ cells, which have been shown to represent epiblast-like cells [6,9,13], and may explain the expression of the epiblast marker Fgf5 in the SP fraction. Our molecular and functional data suggest that these cells would correspond to early epiblast-like populations, as we were unable to find significant differences in the expression levels of other markers described up-regulated in EpiSc, such as Nodal, Eomes, and Bry [37,38], and EPLs, such as Uvo [39]; also SP ESCs displayed reversibility and ability to contribute to developing morula, which is consistent with the properties of ICM and early epiblast cells [6,9,13]. Correlated with these results, the relative levels of expression of Fgf5 between SP and non-SP populations also seemed lower than the ones reported between ESCs and EpiSc [13,38]. The SP population also displayed elevated levels of Rex1, Tbx3, and Stella, which have been reported down-regulated in both EpiSc and EPLs [37 –39], as well as alkaline phosphatase activity, which is absent in EpiSc [13,37,38]. Together with the results of the functional assays, the antigenic and molecular profiles suggest that the SP population of ESC is heterogeneous and contains cells that resemble pluripotent cell types of the ICM and early epiblast of the peri-implanted embryo.
In agreement with our marker data, we found that SP ESCs displayed in vitro and in vivo pluripotentiality, as they differentiated into derivatives of mesoderm, ectoderm, and endoderm in embryoid body and teratoma assays. In addition, purified SP ESCs exhibited increased ability to integrate into developing morulae, which is consistent with the properties of ICM and early epiblast cells rather than EpiSc [37,38]. However, our current study cannot rule out the presence of a small fraction of EpiSc-like cells within the SP population, as these cells are pluripotent and express SSEA1. Similarly, we cannot exclude the possibility that the SP fraction may contain primordial germ cell (PGC)-like cells, as we found increased expression of the PGC markers Stella and Blimp in it, although expression of the latter did not seem to show statistically significant difference. However, ICM cells have been shown to express Stella [41,42], and Stella+ ESCs have been reported to functionally resemble ICM cells but not PGCs [13], which is in agreement with our results. Functional analysis of different antigenic fractions of the SP population will help address this aspect. Nevertheless, our antigenic and molecular analyses, EB, teratoma, and ES-morula aggregation assays clearly demonstrate that the SP population contains pluripotent cells that phenotypically and functionally resemble pluripotent ICM-like cells.
Supporting these results, we found that purified SP cells reconstituted ESC cultures that contained both SP and non-SP populations. Similarly, non-SP cells were able to produce cultures containing both populations. This reversibility is consistent with the properties of other ICM- and early epiblast-like ESC subpopulations [9,13]. Also, we found that an equilibrium consistently established between SP and non-SP fractions under standard ESC conditions, which suggests that an intrinsic program [13] controls the conversion of SP and non-SP phenotypes. Because the conversion of SP into non-SP ESCs seems to occur faster than the reciprocal, progression from the SP to non-SP phenotype would be the default program under ESC culture conditions. This might explain why while most of the ICM cells of the embryo display SP phenotype, only a small population of ESCs exhibits this property. This also suggests that ICM and epiblast-like SP cells may be in dynamic equilibrium [13], which would explain the heterogeneity of these markers in the SP fraction. Because multiple signals maintain pluripotency in ESCs [3,5], it is possible that the proportion of ICM-like and epiblast-like SP phenotypes within the ESCs changes depending on culture conditions that favor one state or the other [9,13].
Last, using staining conditions for the SP ESCs, we found cells with similar efflux properties in the ICM of peri-implanted blastocysts. These cells displayed heterogeneous and overlapping patterns of Ho and Rho efflux, resembling the ones we found for ESC. Our observations appear to be in agreement with recent reports describing heterogeneity for other markers in the ICM [43,44]. Our results suggest that the ICM-like cells of the SP fraction of ESCs would correspond to the efflux-active ICM cells of the peri-implanted embryo, and expression of Abcg2, Abcb1b, and Abcc3 may contribute to these properties. Supporting this idea, expression of Abcg2 and Abcb1b has been described for early embryos and associated with Ho and Rho efflux [40]. In addition, ESTs for Abcc3 has been described for the morulae stage, when SP-like cells start to emerge (data not shown). Together, this supports the idea that the SP phenotype is developmentally regulated [23 –25], and controlled by different transporters in different stem cells along development [24,25]. In ESCs, this phenotype would reflect an ability to translocate molecules that is inherent to efflux-active cells of the ICM and early epiblast progenitors contained in it [43,44].
In summary, our results indicate that ESC contain an SP population that constitutes a phenotypically and functionally distinct fraction within ESCs, as it displays markers and biological properties that distinguish it from the non-SP ESC fraction. This SP population reconstitutes ESC cultures and contains cells that resemble efflux-active ICM cells of peri-implanted blastocysts. Together, our observations indicate that the SP marker is not exclusive to multipotent stem cells, but it is also present in embryo-derived luripotent cell types, and suggest an involvement of the SP phenotype in ESC maintenance and early embryo development. Further studies on the SP ESCs will help unveil the precise role and molecular mechanisms controlling the SP phenotype in stem cells.
Footnotes
Acknowledgments
We thank all the members of the Goodell's lab for insightful discussions; Claudia Andreu-Vieyra from the Matzuk's lab for critiques and help in blastocyst studies; and Isabel Lorenzo from the BCM Transgenic Mouse and ES Targeting Core for technical advice. This work was supported in part by Canadian Institutes of Health Research and National Institutes of Health (NIH) Postdoctoral Fellowships to D.S.V, and NIH grant RO1-DK058192 to M.A.G.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.