
Abstract
Mesenchymal stromal cells (MSCs) and their precursor cells (MPCs) can proliferate and differentiate into multiple mesodermal and some ectodermal and endodermal tissues. Culture-expanded MSCs are currently being evaluated as a possible cell therapy to replace/repair injured or diseased tissues. While a number of mAb reagents with specificity to human MSCs, including STRO-1, STRO-3 (BLK ALP), CD71 (SH2, SH3), CD106 (VCAM-1), CD166, and CD271, have facilitated the isolation of purified populations of human MSCs from primary tissues, few if any mAb reagents have been described that can be used to isolate equivalent cells from other species. This is of particular relevance when assessing the tissue regenerative efficacy of MSCs in large immunocompetent, preclinical animal models of disease. In light of this, we sought to generate novel monoclonal antibodies (mAb) with specific reactivity against a cell surface molecule that is expressed at high levels by MSCs from different species. Using CD106 (VCAM-1)-selected ovine MSCs as an immunogen, mAb-producing hybridomas were selected for their reactivity to both human and ovine MSCs. One such hybridoma, termed STRO-4, produced an IgG mAb that reacted with <5% of human and ovine bone marrow (BM) mononuclear cells. As a single selection reagent, STRO-4 mAb was able to enrich colony-forming fibroblasts (CFU-F) in both human and ovine BM by 16- and 8-folds, respectively. Cells isolated with STRO-4 exhibited reactivity with markers commonly associated with MSCs isolated by plastic adherence including CD29, CD44, and CD166. Moreover, when placed in inductive culture conditions in vitro, STRO-4+ MSCs exhibited multilineage differentiation potential and were capable of forming a mineralized matrix, lipid-filled adipocytes, and chondrocytes capable of forming a glycosaminoglycan-rich matrix. Biochemical analysis revealed that STRO-4 identified the beta isoform of heat shock protein-90 (Hsp90β). In addition to identifying an antibody reagent that identifies a highly conserved epitope expressed by MSCs from different species, our study also points to a potential role for Hsp90β in MSC biology.
Introduction
Ovine models of human orthopedic and cardiac disease/trauma have been widely used, as sheep share similarities with human anatomy, physiology, immunology, and embryonic development [8 –17]. While several human MSC-specific markers such as STRO-1, CD106, CD146 have been described in the literature [2,3], significant progress in examining the therapeutic potential of MSCs in ovine models of human disease has been limited due to lack of specific reagents that enable the isolation and characterization of an equivalent cell population in ovine tissues.
Therefore, the development of biological reagents, such as monoclonal antibodies, reactive with both ovine and human MSCs would greatly facilitate the capacity to monitor and equate the functional properties of both ovine and human MSC populations in preclinical and clinical trials, respectively. In the present communication, we describe a monoclonal antibody reagent, STRO-4, which identifies clonogenic MPCs (CFU-F: colony-forming unit-fibroblastic) in unfractionated ovine and human BM and is highly expressed by their MSC progeny following ex vivo expansion. Using biochemical and mass spectrometry methods, we found that STRO-4 identifies a cell surface form of the ubiquitously expressed chaperone protein, Hsp90β. Importantly, STRO-4 was found to selectively isolate ovine and human MPCs that exhibit high proliferative potential and multilineage differentiation capacity, suggesting that antibodies to Hsp90β may be used as an effective single marker that identifies a highly conserved epitope expressed by MSCs from both ovine and human tissues. Finally, while outside the scope of the present study, our findings highlight a potential role for Hsp90β in MSC biology.
Materials and Methods
Tissue samples
Normal human adult BM aspirates were obtained under the approved guidelines of the Human Ethics Committee of the Royal Adelaide Hospital. Ovine BM aspirates were obtained under the approved guidelines of the Animal ethic committee of the Institute of Medical and Veterinary Science. Ovine and human bone marrow mononuclear cells (BM MNCs) were prepared by density gradient separation essentially as described previously [2]. Human and ovine abdominal wall-derived adipose tissue; dental pulp tissue, and periodontal tissue were isolated and processed as previously described [18
–21] under the approved guidelines of the Animal Ethic Committee of the Institute of Medical and Veterinary Science. Cultures of human and ovine BM MSCs, adipose stromal/stem cells (ASCs), dental pulp stem cells (DPSCs), and periodontal ligament stem cells (PDLSCs) were established and maintained in α-MEM supplemented with 20% fetal calf serum (FCS), 2 mM
Immunization of mice and production of antibody-secreting hybridoma cell lines
Using the QE4G9 mAb (a kind gift from Ravi Krishnan, The Queen Elisabeth Hospital, Woodville, Australia), CD106 (VCAM-1)-immunoselected clonogenic ovine MPCs were established as primary cultures as previously described [22]. The progeny of ex vivo expanded MPCs, MSCs were harvested and resuspended in 300 µL PBS supplemented with 20 µg muramyl dipeptide (Sigma Chemical Company, St. Louis, MO) as adjuvant. BALB/c mice were immunized intraperitoneally with 5 × 106 ovine MSCs, and subsequently boosted a further three times at 3-week intervals to ensure adequate affinity maturation of the immune response. Three days prior to fusion, 5 × 106 cells resuspended in 100 µL PBS were administered via the tail vein. Immediately prior to fusion, mice were sacrificed and their spleens aseptically removed. Hybridomas secreting antibodies reactive with cultured MSCs were made by fusing the NS1-Ag4-1 murine myeloma cell line and spleen cells derived from BALB/c mice immunized with trypsinized cultured ovine MSCs. Fusion of splenocytes and myeloma cells was performed essentially as previously described [23,24]. Hybridomas were screened for their low reactivity with BM MNCs and reactivity with the primary immunogen (cultured ovine MSCs) and human-cultured MSCs. The STRO-4 hybridoma was also selected for its reactivity and enrichment of ovine and human colony-forming MPCs (CFU-F).
Mesenchymal stromal cell culture
Colony-forming efficiency assays were performed at plating densities ranging from 0.1 to 1 × 104 unfractionated or immunoselected BM MNCs per square centimeter in triplicate six-well plates over a 12-day period. The cells were grown in α-MEM supplemented with 20% FCS, 2 mM
Differentiation assays
To induce osteogenesis, ex vivo expanded MPCs were cultured in α-MEM supplemented with 10% FCS, 100 µM
Magnetic-activated cell sorting (MACS)
This was performed as previously described [2]. In brief, ∼1–3 × 108 normal ovine or human BM MNCs were incubated with anti-VCAM-1/CD106 or STRO-4 neat supernatant, for 1 hour on ice. The cell preparations were then incubated with goat anti-mouse IgG streptavidin microbeads and finally streptavidin FITC (1/50; Caltag Laboratories, Burlingame, CA) for 30 min on ice before being separated on a Mini MACS magnetic column (Miltenyi Biotec Inc., Auburn, CA) according to the manufacturer’s instructions.
Indirect immunofluorescence and flow cytometric analysis
Prior to immunolabeling, cultured cells (ovine and human MPCs, DPSCs, PDLSCs, ASCs, MG63, and SAOS) were incubated in blocking buffer (HBSS + 20 mM Hepes, 1% normal human AB serum, 1% bovine serum albumin (BSA: Cohn fraction V; Sigma Aldrich Pty Ltd, NSW, Australia), and 5% FCS for 20 min on ice. Aliquots of 1 × 105 cells were resuspended in 100 µL of supernatant of either STRO-4 or a nonbinding isotype-matched IgG1 antibody (1B5 kindly provided by Professor L.K. Ashman, University of Newcastle, Australia) for 45 min on ice. The cells were then washed in HBSS with 5% FCS and incubated with a goat anti-mouse IgG (γ-chain specific) phycoerythrin (PE) (1/50; Southern Biotechnology Associates, Birmingham, AL) for 45 min on ice. Prior to analysis, cells were washed twice in HBSS with 5% FCS and resuspended in PBS/1% paraformaldehyde. Flow cytometric analysis was performed using a Coulter Excel flow cytometer (Coulter Corp., Hialeah, FL). Positivity for each antibody was defined as the level of fluorescence greater than 99% of what was observed when isotype-matched, nonbinding control antibodies were used. Twenty thousand events were collected per sample as list mode data and analyzed using Coulter ELITE software.
Immunoprecipitation, protein identification, and western blot analysis
Cell lysates were prepared as previously described [2]. Goat anti-mouse Ig-coupled Dynabeads (Dynal, Oslo, Sweden) were washed twice in 1% (v/v) NP40–50 mM Tris-HCl, 150 mM NaCl, 1 mM EDTA [TSE] prior to the addition of neat STRO-4 supernatant and isotype-matched nonbinding control (1B5). This mixture was then incubated at 4°C for a minimum of 6 h, with rotation. The resulting prearmed Dynabeads were washed twice in 1% NP40-TSE, and the beads collected using a magnetic particle collector (MPC-1, Dynal). To these, 1.0 mL aliquots of the appropriate NP40 cell lysate were added. The samples were incubated for 2 h at 4°C with rotation. The immunoprecipitates were then washed twice in 1% (v/v) NP40-TSE, once in 0.1% (v/v) NP40-TSE, and once in TSE, pH 8.0. The supernatant was then removed and samples stored at –20°C or used immediately for electrophoresis. Each immunoprecipitate represented the material from 5 × 106 cell equivalents. Samples were boiled for 3 min in 25 µL reducing sample buffer (62.5 mM Tris, 3% [w/v] SDS, 10% [v/v] glycerol, and 5% [v/v] 2-mercaptoethanol) and analyzed by 10% (w/v) SDS-polyacrylamide gel electrophoresis using Coomassie blue as previously described [24].
For mass spectrometry, excised bands were subjected to 16 h of tryptic digest at 37°C. The samples were then desalted and concentrated using a Millipore C18 ZipTip and a 1 µL aliquot was spotted onto a sample plate with 1 µL of matrix (α-cyano-4-hydroxycinnamic acid, 8 mg/mL in 70% v/v AcN, 1% v/v TFA) and allowed to air-dry. MALDI mass spectrometry was performed with an Applied Biosystems 4700 Proteomics Analyzer with TOF/TOF optics in MS mode. A Nd:YAG laser (355 nm) was used to irradiate the sample. The spectra were acquired in reflectron mode in the mass range 750–3,500 Th. The data was exported in a format suitable for submission to the database search program Mascot (Matrix Science Ltd, London, UK).
For immunoblots, SDS-PAGE resolved proteins were transferred to polyvinyl difluoroacetate (PVDF; MSI Membranes, Geneworks, Adelaide, Australia) at 30 mA overnight in a wet blotting apparatus (Hoefer Scientific Instruments, San Francisco, CA). After blocking for 2 h with 5% skim milk powder–0.05% Tween-20 in PBS, the filters were incubated with a polyclonal antibody to human HSP90β (Santa Cruz Biotechnologies Inc., Santa Cruz, CA) for 1 h at room temperature. The primary antibody was subsequently detected with goat anti-mouse–alkaline phosphatase conjugate (Amersham Biosciences, Poole, UK) and immunoreactive proteins were visualized on a FluorImager (Molecular Dynamics, Sunnyvale, CA) using ImageQuant software (Molecular Dynamics).
Results
Generation of the monoclonal antibody STRO-4 that identifies ovine and human clonogenic MPCs
A panel of mouse mAb reactive with ovine- and human-cultured MSCs were generated following the fusion of splenocytes derived from mice immunized with CD106+ (VCAM-1) selected ovine BM MSCs. Preliminary screens were designed to identify mAbs that exhibited limited reactivity with human and ovine peripheral blood (PB) MNCs, but reacted with cultured ovine and human MSC-like populations from a variety of tissues, including periodontal ligament (PDLSCs), dental pulp (DPSCs), and adipose (ASCs). One such mAb, STRO-4 was found to react at uniformly high levels with both human and ovine stem cell populations from these various tissues (Fig. 1).

STRO-4 binds to a cell surface antigen expressed by human and ovine mesenchymal stromal cell (MSC)-like populations derived from different tissues. Single cell suspensions of passage 2 human (
While exhibiting limited reactivity with BM MNCs from human and sheep (Fig. 2A and 2B), enrichment of STRO-4+ BM cells using MACS yielded populations of human and ovine cells with enhanced colony-forming unit-fibroblast (CFU-F) activity. Specifically, STRO-4+ selection resulted in a 16- and 8-fold enrichment of CFU-F relative to their incidence in unseparated BM MNC samples prior to MACS separation (Fig. 2C and 2D). Immunostaining of primary cultures of human and ovine STRO-4-selected cells with antibodies to hematopoietic and MSC-related markers revealed that while lacking measurable expression of leukocyte (CD14, CD45) and endothelial (CD31) markers, STRO-4+ MSCs expressed appreciable levels of CD29, CD44, and CD166 (Fig. 3).
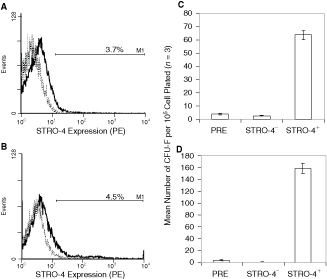
STRO-4 is expressed by a minor population of ovine and human bone marrow (BM) mononuclear cells (MNCs) that includes essentially all clonogenic mesenchymal precursor cells (MPCs). Single-color immunofluorescence and flow cytometry were performed on specimens of Ficoll-separated ovine and human BM MNCs to examine the expression of STRO-4+. The mean incidence of STRO-4+ cells in the ovine (
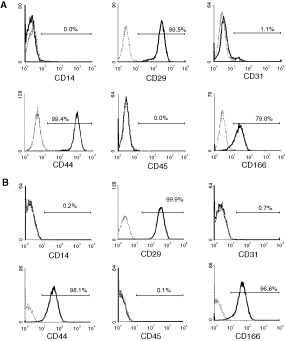
Ex vivo expanded STRO-4− selected mesenchymal stromal cells (MSCs) lack expression of hematopoietic and endothelial markers and express common MSC markers. Single-color immunofluorescence and flow cytometry was performed on single cell suspensions of Passage 2 STRO-4+ ovine and human bone marrow (BM) MSCs with antibodies to available hematopoietic and mesenchymal stromal-related markers that reacted with both ovine (
Developmental potential of human and ovine STRO-4+ BM MSCs in vitro
Comparison of the in vitro developmental potential of human (Fig. 4A) and ovine (Fig. 4B) STRO-4+ cells (n = 3 BM samples) revealed that these cells were capable of readily forming clusters of Oil Red O-positive lipid-laden adipocytes when cultured in adipogenic inductive media. Similarly, human and ovine STRO-4+-derived populations formed Alizarin Red-positive mineralized deposits after several weeks culture in osteogenic induction conditions. The STRO-4 selected BM MSC also displayed chondrogenic morphology when cultured in an aggregate culture system in the presence of TGFβ1. Furthermore, human (Fig. 4C) and ovine (Fig. 4D) STRO-4+ MSCs when plated in the presence of TGFβ1 were capable of secreting elevated levels of glycosaminoglycan, a major component of the cartilage-like matrix [2].
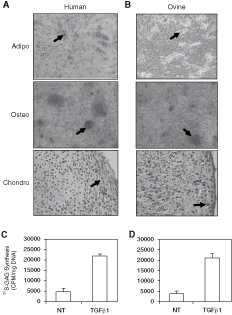
Developmental potential of STRO-4+ selected ovine- and human-cultured mesenchymal precursor cell (MPC) secondary cultures of mesenchymal stromal cells (MSCs) derived from STRO-4+ selected ovine and human MPCs were induced under either adipocytic, osteogenic, or chondrocytic conditions. The presence of clusters of lipid-containing adipocytes were detected by Oil Red O staining (arrow) within 2 weeks of adipogenic induction in the human (
Characterization of the antigen recognized by STRO-4
Immunoprecipitation was used to purify the STRO-4 antigen from the human osteosarcoma cell line, MG63, which expresses high cell surface levels of the antigen (data not shown). Proteins precipitated with STRO-4 and 1B5 were resolved by SDS-PAGE and visualized using Coomassie blue staining. A single protein with an apparent molecular mass of 90 kDa was purified to near homogeneity using STRO-4 (Fig. 5A). To obtain a protein identification, MALDI-TOF mass spectrometry and peptide mass fingerprinting was performed on tryptic peptides generated from the digestion of SDS-PAGE purified protein. The peptide masses were used to search SWISS-PROT and TREMBL databases using Mascot (Matrix Sciences Ltd, London, UK). Of the 91 mass values searched, 17 (25%) matched peptide masses generated for human heat shock protein-90 beta isoform (HSP90β) (Fig. 5B). To confirm that the STRO-4-reactive 90 kDa peptide was Hsp90β, whole cell lysates and STRO-4 immunoprecipitated extracts were resolved by SDS-PAGE and western blot analysis was performed using a commercially available rabbit polyclonal antibody reactive with the C-terminus of the human Hsp90β peptide. The STRO-4-immunoprecipitate exhibited specific reactivity with the Hsp90β-specific antibody at the appropriate molecular weight of 90 kDa (Fig. 5C).
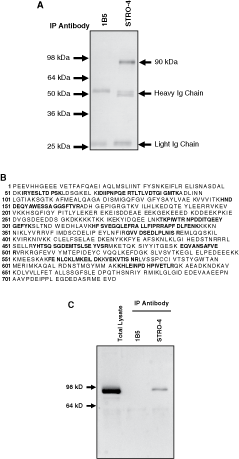
The STRO-4 antibody identifies heat shock protein-90β (HSP90β). Plasma membrane lysates were prepared from MG63 cells as described in the methods. The cell lysates were incubated with either STRO-4 supernatant or the isotype-matched control antibody, 1B5, and immunoprecipitated by magnetic bead separation. Immunoprecipitated proteins were resolved on a 10% SDS-PAGE gel and visualized using Coomassie blue staining (
Discussion
Large animals are frequently used as models for preclinical research, due to their similarities with humans, with regard to organ size and structure [25 –27]. Increasingly, different culture-expanded MSC preparations are looked upon as potential cell-based therapies to augment tissue repair following trauma or disease processes. MSCs possess numerous attributes including their plasticity, ease of ex vivo expansion, and apparent lack of immunogenicity making them suited for a range of applications [28]. However, the majority of studies have relied on the use of plastic adherence as the main mode of isolation for MSCs, a technique that results in a heterogeneous population, comprised of immature and mature stromal cells, endothelial cells, and macrophage. The development of monoclonal antibodies that preferentially identify MSCs from hematopoietic cells has allowed the isolation and expansion of partially purified MSC cultures without the presence of mature stromal elements or other accessory cell populations. Despite the growing numbers of reagents that can be used to facilitate the isolation of human MSC precursor populations [29], few if any reagents have been described to isolate an equivalent cell population in other species.
The present communication describes the generation and characterization of a novel monoclonal antibody, STRO-4, that identifies a minor subset of human (∼2.5%–4.5%) and ovine (∼1.5%–3.7%) BM cells and was not expressed by PB MNCs. The antigen was raised against CD106+ (VCAM-1)-selected ovine MPCs that have previously been shown to be effective in regenerating bone tissue in a preclinical ovine model of nonunion bone fracture [22]. Secondary screens also showed that the STRO-4 antigen was highly expressed on the cell surface of different culture-expanded MSC-like populations derived from a variety of human and ovine tissues. Importantly, the STRO-4 antibody exhibited the capacity to isolate essentially all of the clonogenic MPCs from BM aspirates in both species. Furthermore, the progeny of cultured human and ovine MPCs derived from the STRO-4+ fraction of BM, demonstrated the capacity for multilineage differentiation in vitro, and exhibited an immunophenotype associated with cultured MSCs (CD29+/CD44+/CD166+/CD14−/CD45−/CD31−). Collectively the data presented here show that in adult human and ovine BM, the antigen identified by STRO-4 is highly specific for stromal elements and their precursors. Therefore, the STRO-4 mab appears well suited as a common reagent for the isolation and characterization of different human and ovine MSC-like populations destined for different preclinical and clinical studies, respectively.
Using a biochemical approach, STRO-4 was found to identify a unique epitope expressed on heat shock protein-90 beta (Hsp90β) [30 –32]. Hsp90β belongs to a family of proteins that are induced by cellular stresses such as temperature change [33]. This highly expressed family of proteins account for ∼1%–2% of total protein in unstressed cells and up to 4%–6% of cellular proteins in stressed cells [34]. Although sharing 85% sequence identity with its environmental stress-inducible homolog, Hsp90α, the predominantly cytosolic protein Hsp90β is constitutively expressed. Hsp90 proteins are multifunctional, with roles in “client” protein folding, intracellular transport, maintenance, and degradation of proteins, as well as facilitating cell signaling [35] and differentiation [36]. Furthermore, Hsp90b has been shown to suppress the aggregation of a wide range of “client” proteins, thereby acting as a general protective chaperone [37 –39].
While considered to be predominantly an intracellular protein, recent studies suggest that both Hsp90α and Hsp90β can exist at the cell surface [40,41]. Expression of Hsp90β at the cell surface has been described in a number of cell types including oligodendrocyte precursor cells (OPC) [42] and neuronal cells, where it plays a role in cell migration [43]. Moreover, cell surface Hsp90β has been described in a range of cancer cell types including fibrosarcoma and melanoma cells [44], where it has been linked to heightened metastatic potential of melanoma cells [45]. In addition to their role in cell migration, HSP90 proteins also play a survival role in cancer cells by stabilizing a number of proteins involved in the oncogenic process, including v-Src, the fusion oncogene Bcr/Abl, p53, PI3K, and AKT [46].
The role of Hsp90 in MSC proliferation and differentiation is yet to be determined and awaits further study. However, it is tempting to speculate, that like cancer cells, MSCs may exploit surface-expressed Hsp90 to facilitate cell motility, allowing MSCs to be recruited to sites of tissue damage. Moreover, MSCs like cancer cells possess extensive proliferative (self-renewal) capacity and are dependent upon many of the signaling pathways exploited by cancer cells to proliferate and/or differentiate, including the glucocorticoid receptor (GR) pathway [47]. Given their critical role in maintaining the health of cells by ensuring the appropriate folding of critical proteins and the degradation of unwanted and/or harmful proteins, Hsp90 expression by MSCs may well be critical to their survival and well-being.
In summary, in the present study, the ovine and human MPC populations selected on the basis of STRO-4 cell surface expression demonstrated extensive proliferation in vitro, while retaining their capacity for differentiation into bone, cartilage, and adipose tissues. Moreover, to the best of our knowledge, this represents the first report demonstrating that the Hsp90β protein is expressed on the cell surface of immature MPC in vivo. Moreover, these studies suggest that antibodies to this molecule may be used as an effective means of isolating and enriching MPCs from various ovine and human tissues.
Footnotes
Acknowledgment
This work was supported by grants from the Australian National Health & Medical Research Committee (A.C.W.Z., S.G., and P.M.B.).