
Abstract
In a continuous effort to improve the generation of therapeutic grade human embryonic stem cell (hESC) lines, we focused on preserving developmental capacity of the embryos, minimizing the exposure to xenomaterials, increasing derivation efficacy, and reducing the complexity of the derivation procedure. In this study, we describe an improved method for efficient derivation of hESC lines from blastomeres of biopsied embryos. Our protocol substituted feeder cells of mouse origin with human foreskin fibroblasts (HFFs), limited serum exposure of cells to formation of the initial outgrowth, and increased derivation efficacy from 12.5% (one hESC line out of 13 biopsies) to 50% (3 out of 6 biopsies) by using early population doubling (PD) HFFs. In addition, it eliminated a need for embryo-blastomere coculture, thus reducing the complexity of the culture and enabling continued development of the biopsied embryo under optimal conditions. All derived lines maintained normal karyotype and expressed totipotent phenotype including the ability to differentiate into trophectoderm and all three germ layers.
Introduction
S
In this study, we report a substantial advancement in the previously reported method for generating hESC from biopsied blastomeres that uses human feeder cells instead of MEF and limits exposure to FCS to the formation of initial outgrowth from biopsied blastomeres. Moreover, we demonstrate that the coculture with parental embryo is not essential for the derivation of hESC line from biopsied blastomeres, allowing culture of the biopsied embryo under optimal conditions. In this way, we minimized the exposure of derived hESC to both xenoantigens and xenopathogens, while preserving the viability of the embryos.
Materials and Methods
Embryos
Embryos used in these experiments were frozen surplus embryos donated by consenting patients. The protocol has been approved by IRB and SCRO oversight committees.
Alkaline phosphatase assay
The assay was performed according to the manufacturer’s instructions (ATCC).
Immunostaining
Undifferentiated or in vitro differentiated hESC cultures were fixed in 90% acetone or 3.8% paraformaldehyde and then permeabilized in 90% acetone and incubated with the primary antibodies against SSEA-4, TRA 1-60, TRA 1-81 (Millipore); nanog (R&D); Oct 3/4 (Santa Cruz Biotechnology Inc.); and SMA, βIII tubulin, or αFP (Sigma). All secondary antibodies were purchased from Jackson ImmunoResearch, whereas Hoechst was obtained from Invitrogen.
RT-PCR
RNA was isolated using RNAeasy Plus Minikit (Qiagen) and converted to cDNA using SuperScript III CellsDirect cDNA Synthesis System (Invitrogen). The PCR was performed using Platinum PCR SuperMix (Invitrogen). The PCR products were analyzed by gel electrophoresis using ethidium bromide (1 µg/mL) stained 2% agarose gels and photographed under ultraviolet illumination. The primer sequence, annealing temperature, number of PCR cycles, and fragment size are provided in Supplementary Table 1; Supplementary materials are available online at http://www.liebertpub.com).
Karyotyping
Karyotyping has been performed by certified Cytogenetics Laboratory (Children’s Hospital and Research Center, Oakland, CA) after 1 and 6 months of continuous culture of each hESC line.
DNA profiling
DNA profile [6] for each hESC line was determined by DNA testing laboratory Identigene (Houston, TX).
Results
Generation of hESC lines with minimal exposure to xenomaterials
In the first set of experiments, our aim was to minimize the exposure to xenopathogens during derivation of hESC lines from biopsied blastomeres. Hence, we attempted to establish hESC line from biopsied blastomeres on feeders of human origin and to minimize the period in which derivation medium contains FCS. From various human origin cells that support derivation and growth of hESC, we choose human foreskin fibroblasts (HFFs), the most popular MEF alternative.
Thirteen grade 1 and 2 surplus frozen human cleavage stage embryos donated by three consenting couples (W5, W6, and W8) were thawed and incubated in Quinn’s cleavage medium for at least 3 h under standard culturing conditions (Fig. 1A). Blastomeres were removed from each embryo using established biopsy procedure [5]. Biopsied blastomeres and parental embryos were cultured in the same drop of Quinn’s cleavage medium for 24 h. Parental embryos were then transferred to Quinn’s blastocyst medium for additional 48–72 h and at blastocyst stage processed for cryopreservation. About two-thirds of biopsied blastomeres (69.2%) divided during the coculture period. All blastomeres were transferred on to irradiated HFF PD14 in drops of Quinn’s cleavage medium supplemented with 10 µg/mL laminin (day 0). Three days later, cultures were assessed for blastomere attachment to feeders. Eight out of nine (88.9%) divided blastomeres attached and five out of eight (62.5%) gave an initial cell outgrowth by day 5. Although two biopsied blastomeres that did not divide during coculture attached, neither of them gave an outgrowth. Starting on day 3, medium in drops containing attached blastomeres has been refreshed every day by replacing one-third of the volume with Quinn’s blastocyst medium supplemented with laminin, 10 ng/mL leukemia inhibiting factor (LIF), and 25 ng/mL basic fibroblast growth factor (bFGF), all of human origin. From day 5, Quinn’s blastocyst medium was replaced with standard hESC medium (80% KnockOut-DMEM, 20% KSR, 25 ng/mL bFGF) enriched with 10% FCS, and LIF, and replaced in drops on a daily basis. Most of trophectoderm-like outgrowth died on days 6 and 7. On day 9, we detected an initial hESC colony in one out of five cultures (20%). Two days later, a colony was dissected within the same drop. The procedure was repeated on day 14. After the second dissection, small hESC clumps were transferred into 4-well dish with freshly seeded HFF PD14 feeders. The next day, FCS and LIF were withdrawn from the attached hESC clumps. Culturing was continued in the standard serum-free hESC medium. The newly derived cell line had a typical morphology of hESC. We used enzymatic assays (alkaline phosphatase/AP activity), immunostaining (Oct 3/4, nanog, SSEA-4, TRA 1-60, and TRA 1-80), and RT-PCR (nanog, Oct 3/4, TDGF1, DNMT3B, GABRB3, GDF3, and TERT) to confirm the expression of pluripotency markers (Fig. 1B and 1C). The ability to differentiate into derivatives of all three embryonic germ layers has been confirmed by immunostaining and RT-PCR (Fig. 1D and 1E). Brachyury, WT1, and SMA were chosen as markers of mesoderm; GATA4, SOX17, and αFP as markers of endoderm; and βIII tubulin, nestin, and PAX6 as markers of ectoderm derivatives. Moreover, we also detected the expression of two trophoblast markers CDX2 and hCG-β, confirming totipotency of newly derived line W8-8A. The W8-8A hESC line has shown and retained stable normal female karyotype (46, XX) during 6 months of continuous culture(Fig. 1F). Analysis of 13 short-tandem repeat (STR) loci provided unique DNA profile for W8-8A hESC line that did not match any other hESC line present in the laboratory eliminating any chance of cross-contamination with other hESC lines (Fig. 1G).
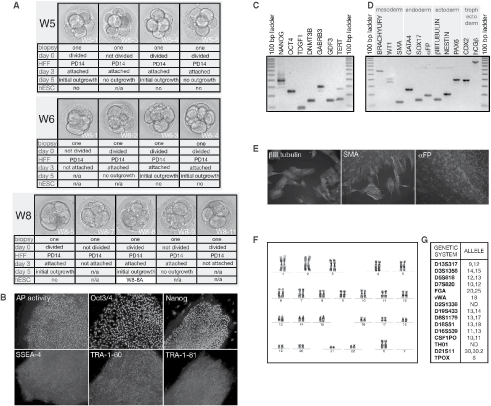
Generation of human embryonic stem cells (hESCs) from biopsied blastomeres on feeders of human origin. (
In this set of experiments, we demonstrated the feasibility of the hESC derivation from biopsied blastomeres on human feeders. We also found that 10-day exposure to FCS is sufficient to obtain a successful hESC line derivation.
Improvement of hESC derivation efficacy on HFFs
With at least 40 population doublings (PDs) before exhibiting the first signs of cellular senescence and anticipated mitotic life span of 60–90 PDs, HFFs appear to be much more “robust” cells than MEF, the most widely used feeder cells for derivation and maintenance of hESC. Indeed, MEFs are used for this purpose only at very low passages (PD2-4), whereas HFFs are known to support hESC derivation from blastocysts at PD9-25 [7,8], and also from a single viable blastomere within an arrested four-cell stage embryo [9]. Although there are no comparative studies showing whether PD of the HFFs affects derivation efficacy, we hypothesized that, similar to MEF, HFFs with lower PD may be more efficient in supporting hESC derivation. To determine whether this was the case, we conducted a second set of experiments in which we compared hESC derivation efficacy from genetically identical blastomeres seeded on PD14 and PD6 HFFs.
Six grade 1 and 2 surplus frozen human cleavage stage embryos donated by two consenting couples (W10 and W13) were thawed and incubated in Quinn’s cleavage medium for at least 3 h under standard culturing conditions (Fig. 2A and 2H). To obtain genetically identical samples for the comparison, two blastomeres were biopsied from each embryo. Similar to the previous experiment, about two-thirds of biopsied blastomeres divided (66.7%) during the initial 24 h coculture with parental embryos. A half of the biopsied material from individual embryo was then transferred on to HFF PD14 and another half on PD6. Culture conditions were as described in the first set of experiments. By day 3, all blastomeres that divided during coculture with a parental embryo, attached to feeder layers, regardless of HFF PD. On day 5, an initial outgrowth was detected in all cultures on PD6 (100%), and in three out of four on PD14 (75%). Growing hESC colonies were observed in two out of four cultures (50%) on PD6 (W10-1A and W13-1C) several days later. However, we did not observe any hESC colony growth on PD14. Both newly derived lines, W10-1A and W13-1C, had typical hESC morphology, and in a series of assays demonstrated their pluripotency (Fig. 2B–E, I–L). The W10-1A hESC line has shown and retained stable normal female karyotype (46, XX), whereas the W13-1C hESC line exhibited stable normal male karyotype (46, XY) during 6 months of continuous culture (Fig. 2F and 2M). For both hESC lines, analyses of 15 STR loci provided unique DNA profiles that did not match any other hESC line present in the laboratory, eliminating any chance of cross-contamination with other hESC lines (Fig. 2G and 2N).
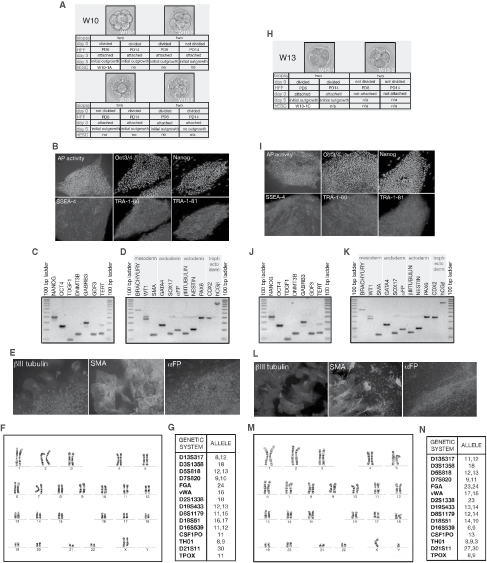
Improvement of human embryonic stem cells (hESCs) derivation efficacy on human foreskin fibroblasts (HFFs). (
In this set of experiments, we demonstrated that the derivation of hESC lines from biopsied blastomeres on HFFs could be improved by the use of feeder cells/fibroblasts with lower PD. Combined results from both sets of experiments suggested that even though HFF PD does not have a dramatic effect on blastomere attachment (100% for PD6 vs. 92.3% for PD14), fibroblasts with lower PD are likely to be better in supporting the formation of an initial outgrowth (100% for PD6 vs. 66.7% for PD14). Superiority of lower PD HFFs in providing an appropriate microenvironment for hESC growth was even more pronounced when we examined their effect on derivation efficiency: it was fourfold higher on PD6 (50%) than on PD14 (12.5%).
Elimination of the embryo coculture
Method developed by Klimanskaya et al. [4] suggested that the coculture of biopsied blastomeres with parental embryo for the initial 24 h could have a beneficial effect for embryonic cells taken out from their microenvironment. However, established assisted reproductive technology protocols recommend different types of culture media for the development of the fertilized egg to cleavage stage embryo (cleavage medium) and from the cleavage stage to blastocyst formation (blastocyst medium). Keeping cleavage stage embryo for the extended period of time (24 h) in cleavage medium, a medium required to initiate the division of biopsied blastomeres, may retard embryo progress to blastocyst stage. In the final set of experiments, our aim was to determine whether coculture with parental embryo is essential for the successful derivation of hESC lines from biopsied blastomeres.
Two grade 2 surplus frozen human cleavage stage embryos donated by one consenting couple (W14) were thawed and incubated in Quinn’s cleavage medium for at least 3 h under standard culturing conditions (Fig. 3A). To obtain genetically identical samples for comparison, we followed the same protocol as in the second set of experiments, and performed two blastomere biopsies on each embryo. A half of biopsied blastomeres from each embryo were then transferred into another drop of Quinn’s cleavage medium, whereas another half remained for 24 h in the same drop with parental embryo. From one of the embryos (W14-1) both individually cultured and the cocultured blastomere divided within the first 24 h, whereas from the second embryo (W14-2) neither one did. Two blastomere-derived cell aggregates from embryo W14-1, as well as undivided blastomeres from the embryo W14-2, were transferred into individual drops with PD6 HFFs. Culturing conditions were the same as described in the first two sets of experiments. Whereas neither blastomere from the embryo W14-2 progressed further, both blastomere-derived aggregates from embryo W14-1 attached and formed an initial outgrowth by day 5. A week later, only the outgrowth from individually cultured biopsied blastomere gave rise to hESC line (W14-1A). Newly derived line W14-1A had typical hESC morphology, and in a series of assays demonstrated pluripotency (Fig. 3B–E). The W14-1A hESC line has shown normal female karyotype (46, XX) (Fig. 3F). Analyses of 15 STR loci provided unique DNA profile for W14-1A that did not match any other hESC line present in the laboratory eliminating any chance of cross-contamination with other hESC lines (Fig. 3G).

Generation of human embryonic stem cells (hESCs) from biopsied blastomeres on human foreskin fibroblasts (HFFs) without coculture with parental embryo. (
Although the number of samples in the last set of experiments was too small to determine whether the presence of parental embryo affects hESC derivation efficacy, it unequivocally demonstrated that the coculture of biopsied blastomeres with parental embryo is not necessary to generate hESC line. Consequently, parental embryo can be cultured under the optimal conditions for its development into blastocyst.
Discussion
In this study, we report an improved method of the hESC derivation from biopsied blastomeres that represent a major step toward the generation of xeno-free therapeutic grade hESC under conditions that enable preserving of the embryo. Given the complex nature of the ethical issues related to research using human embryos, this recently developed approach based on preimplantation genetic diagnostics technique alleviated some of the ethical concerns related to embryo demise. In addition, it presented a novel strategy for obtaining totipotent ESC population from earlier stage of development than blastocyst, before the initial lineage specification that leads to blastocyst formation. However, the requirement for MEF feeder layer in this approach awoke concerns of extensive exposure to xenopathogens and xenoantigens. These concerns were partially eliminated in standard protocols for hESC derivation from blastocyst stage embryos by the introduction of feeders of human origin. Combining the advantages from both approaches, we are the first to demonstrate that hESC lines could be derived from biopsied blastomeres on feeders of human origin, in this case HFFs, while preserving the developing embryo. Furthermore, our protocol limited the use of FCS to establish the initial cell outgrowth and eliminated a need for blastomere coculture with the biopsied embryo.
Our data have important implications regarding the use of human fibroblast feeders in hESC derivation. They clearly demonstrate that the fibroblasts that spent shorter period of time in culture and/or have undergone lower number of PDs better support derivation of hESC from biopsied blastomeres. This suggests that the secretory phenotype of the irradiated fibroblasts is affected by the extent of time spent in culture and/or the number of PDs they have undergone before the irradiation-induced arrest. This was true even though we used fibroblasts within the first quarter of their life span (PD6 vs. PD14). It remains to be seen whether these differences in secretory phenotype affect the derivation efficiency from the blastocyst stage embryos and/or the long-term maintenance and expansion of hESC in culture. In addition, our results open a possibility that by comparing the secretory profile of irradiated HFFs with lower and higher PD, one can identify factors that may be critical for the derivation process and potentially, and may also improve hESC maintenance.
Owing to the fact that we could use blastomeres with identical genetic make-up, we were in a position to eliminate effects due to genetic heterogeneity between embryos and could minimize the number of embryos used for these studies. In general, this approach can be utilized to efficiently study epigenetic changes induced by derivation under different culturing conditions/microenvironments. In addition, described method can be applied for generating hESC lines from cryodamaged embryos with a few viable blastomeres that, if left untouched, would not develop into blastocyst.
Footnotes
Acknowledgments
Experimental work was performed in StemLifeLine facilities and supported by StemLifeLine, Inc. D.I., G.G., E.C., and A.K. were supported by StemLifeLine, Inc. T.Z., O.G., and S.J.F. were supported by CIRM grants RL-00648-1 and RC1-00113-1.