
Abstract
Mesenchymal stem cells (MSCs) are characterized by their hematopoiesis-supporting and immunosuppressive capacity, while osteoclasts are main cell components in the endosteal hematopoietic stem cell niche and pivotal players in osteoimmunology. To clarify the association of these 2 kinds of cells, mouse CD11b+ monocytes were cultured onto MSC layers in the presence or absence of macrophage colony-stimulating factor (M-CSF) and receptor activator of NF-κB ligand (RANKL). The results showed that MSCs independently supported osteoclast development and this effect was enhanced by M-CSF and RANKL. Interestingly, tumor necrosis factor-α (TNF-α)–stimulated MSCs turned to inhibit osteoclast formation and protect tusk slices from osteoclastic resorption. Real-time PCR and ELISA assays demonstrated that osteoprotegerin expression at both mRNA and protein levels in TNF-α–stimulated MSCs was up-regulated, at least partially by activating the mitogen-activated protein kinase pathway. Furthermore, TNF-α–stimulated MSCs maintained their immunophenotypic, multipotential differentiation and immunosuppressive characteristics. Moreover, MSCs treated with synovial fluid from rheumatoid arthritis patients modulated osteoclast generation in close relation with the TNF-α levels. This study suggests that MSCs exhibit dual modulatory function on osteoclasts and the result might shed light on understanding the involvement of MSCs in the inflammatory diseases.
Introduction
Mesenchymal stem cells (MSCs) and their progenies are important supporting cells in hematopoietic stem cell (HSC) niche. MSCs are first reported to reside in bone marrow, and they are also found in the circulation and other tissues [1 –5]. Under certain conditions, MSCs can be induced to differentiate into osteoblasts, adipocytes, and chondrocytes, respectively. In addition, it is demonstrated that MSCs support hematopoiesis via providing suitable cytokines and intercellular contact [6 –8]. Moreover, MSCs express negligible levels of both human leukocyte antigen MHC class II and Fas ligand; they do not express co-stimulatory molecules such as B7-1, B7-2, CD40, or CD40L [9]. Recent investigations reveal that MSCs display general immunosuppressive effect on nearly all immune cells including T lymphocytes [10,11], B lymphocytes [12], natural killer cells [13,14], and dendritic cells (DCs) [15] through cytokine secretion and/or direct cell–cell contact. Clinical trials have been developed and performed in the settings of osteogenesis imperfecta [16], diabetes [17], acute graft versus host disease [18,19], experimental autoimmune encephalomyelitis [20], rheumatoid arthritis (RA) [21,22] and so on.
Osteoclasts, the professional bone-resorbing cells, are derived from monocyte/macrophage hematopoietic lineage and most of them reside near the endosteal zone. Mature osteoclasts are polykaryon encoding tartrate-resistant acid phosphatase (TRAP) and secreting acid and lytic enzymes. Therefore, osteoclasts can degrade bone in vivo and produce pits in bone slice or elephant tusk slices in vitro [23]. Osteoclasts can be generated from bone marrow and spleen progenitors in vitro stimulated by macrophage colony-stimulating factor (M-CSF), receptor activator of NF-κB ligand (RANKL) [24,25], and tumor necrosis factor-α (TNF-α) [26,27]. Meanwhile, osteoprotegerin (OPG), a soluble decoy receptor of RANKL, acts as a potent osteoclast forming inhibitor to neutralize RANKL [28]. Therefore, the balance between RANKL and OPG dominates osteoclastogenesis. Pioneer studies focused mainly on osteoclast ability to maintain bone homeostasis. However, increasing data have suggested that function of osteoclasts may be more than bone resorption. The original work done by Kollet and his colleagues [29] demonstrates that osteoclasts mediate stress-induced mobilization of hematopoietic progenitors, linking directly osteoclasts with regulation of hematopoiesis microenvironment [30]. Moreover, co-stimulatory signals activated by multiple immunoreceptors are essential for osteoclast differentiation and the maintenance of bone homeostasis [31]. Thus, osteoclasts become an important target for modulation to prevent hematopoietic- and bone-related immunologic disorders.
Although MSCs exert general immunosuppression on most immune cells, their role in the modulation of osteoclasts needs to be fully defined. Given our previous study that MSCs can inhibit differentiation and function of monocyte-derived DCs [15] and osteoclasts share monocytes as their common precursor cells with DCs [32], we postulated that MSCs might modulate the generation and bone-resorbing capacity of monocyte-derived osteoclasts. To prove this assumption, the effect of MSCs on development of CD11b+ monocytes into osteoclasts was investigated and the phenomenon suggested that MSCs possess dual modulatory function on osteoclasts.
Materials and Methods
Mice
Normal inbred C57BL/6 (H2b) female mice were purchased from the Laboratory Animal Center, Chinese Academy of Military Medical Science (Beijing; http://www.bmi.ac.cn). All 60 mice were sacrificed in accordance with the Chinese Academy of Military Medical Sciences Guide for Laboratory Animals.
MSC preparation and multipotent differentiation assay
Mouse bone-derived MSCs were prepared and their differentiation along osteoblastic and adipoblastic pathways were evaluated according to the protocols in our laboratory [33]. A histochemical kit (Sigma) was used to assess alkaline phosphatase (ALP) activity according to the manufacturer’s protocol, and the mineralization capacity was evaluated by using von Kossa staining. To assess adipogenic capacity, lipid accumulation was identified with in situ Oil Red O staining.
Isolation of CD11b+ monocytes and CD3+ T lymphocytes
Bone marrow mononuclear cells were harvested from mice (5–6 weeks old) femur and tibiae bone marrow suspensions with Ficoll-Paque (1.073 g/mL; Invitrogen, Carlsbad, CA) density gradient centrifugation. Monocytes were further purified from bone marrow mononuclear cells by using the MACS CD11b+ Isolation Kit (Miltenyi Biotec, Bergisch Gladbach, Germany). Mouse spleen CD3+ T lymphocytes were purified with the same protocol as MACS pan T-lymphocytes Isolation Kit (Miltenyi Biotec, Bergisch Gladbach, Germany).
Osteoclast differentiation
CD11b+ monocytes were cultured alone or co-cultured with MSCs in minimum essential medium, alpha modification (α-MEM; Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum (FBS; Hyclone, Logan, UT) at graded cell numbers with or without addition of different doses of recombinant mouse M-CSF, recombinant mouse RANKL (R&D Systems, Minneapolis, MN), and recombinant mouse TNF-α (Peprotech, Rocky Hill, NJ). In some instances, MSCs were pretreated with TNF-α or rheumatoid arthritis synovial fluid (RASF) samples. Half of the culture medium was replaced every 2 days. In transwell assays, MSCs and monocytes were separated by a 0.4-µm pore size membrane (Corning) to avoid cell–cell contact. MSCs were cultured in the lower compartment while monocytes in the upper compartment. The experiments were replicated every 3 days and at least 3 times repeated measures were done.
Tartrate-resistant acid phosphatase (TRAP) assay
At days 3, 6, 9, and 12, the medium was removed and the cell layers were fixed in citrate-buffered 60% acetone solution (pH 5.4) for 30 s, washed twice with distilled water, and air-dried. TRAP-positive (TRAP+) cells were detected using a commercial TRAP staining kit (Sigma-Aldrich, St. Louis, MO). TRAP+ multinucleated cells (3 or more nuclei, TRAP+ MNCs) were counted manually under a light microscope (Nikon TE2000-U).
Bone resorption pit formation assay
Bone-resorbing activity of osteoclasts was assessed using elephant tusk slices prepared from discarded elephant tusks supplied by Dr. Jin (FU DAN University, Shanghai). Tusk slices (diameter 14 mm, thickness 10 µm) were sterilized with 75% ethanol, air-dried under ultraviolet ray, and washed 3 times with culture medium before they were carefully placed at the bottom of the wells of 24-well cell culture plate. MSCs were seeded onto each slice at the density of 1 × 104/cm2 and cultured for 24 h followed by addition of CD11b+ monocytes (5 × 104/cm2). In some experiments, MSCs preincubated with TNF-α (100 ng/mL) for 24 h were used. The osteoclast culture medium (M-CSF 20 ng/mL, RANKL 20 ng/mL, and with or without TNF-α 100 ng/mL) was changed every 2 days and the culture was maintained for 3 weeks. The cells on the tusk slices were first stained with TRAP to detect osteoclasts followed by cell deprivation with ultrasonicated treatment. The cell-free tusk slices were stained with Toluidine blue to detect resorption pits under a light microscope.
Carboxyfluorescein diacetate succinimidyl ester (CFSE) labeling
CD3+ T lymphocytes were labeled with carboxyfluorescein diacetate succinimidyl ester (CFSE) according to a published method [34]. In brief, mouse spleen purified CD3+ T lymphocytes were suspended at a concentration of 107 cells/mL in PBS containing 2% FBS (Hyclone, Logan, UT). After 20 min of incubation in the presence of 10 µM CFSE, the CFSE incorporation was blocked with addition of a large excessive PBS containing 2% FBS. CD3+ T lymphocytes were then washed twice in PBS containing 2% FBS and cultured in α-MEM containing 10% FBS in the presence of concanavilin A (Con A, 2 µg/mL) with or without MSCs. T lymphocytes were harvested at days 2 and 4. The dye dilution was assayed with a FACSCalibur quantification.
FACS analysis
The MACS-sorting efficiency and the cell surface antigen profile of MSCs were analyzed with flow cytometry on a FACS vantage cytofluorometer (Becton Dickinson, San Jose, CA). The following mAbs were used: phycoerythrin-conjugated monoclonal antibodies against mouse CD31, CD44, CD86, stem cell antigen-1 (Sca-1), MHC-la (histocompatibility complex [MHC] class II) and fluoresceinisothiocyanate-conjugated antibodies against mouse CD11c, GR-1, F480, CD3, CD29, CD45, CD105. (All products were purchased from eBio-Science, San Diego.)
Rheumatoid arthritis synovial fluid (RASF)
RASF were obtained from 20 RA patients (2 men and 18 women) who met the 1987 revised criteria of the American College of Rheumatology. And informed consent was obtained from all patients for research purposes. All RASF samples were freshly isolated with centrifugation and filtered to deplete cellular components. Then the cell-free samples were stored at −20°C until use.
Semiquantitative polymerase chain reaction (PCR) and real-time PCR
MSCs were seeded in 6-well plates at a density of 1 × 105/cm2 and allowed for attachment overnight followed by serum deprivation for at least 6 h. TNF-α was added at graded concentration (5, 10, 50, and 100 ng/mL) or for different time period (2, 4, 6, and 8 h). Prior to TNF-α exposure, MSCs were sometimes pretreated for 1 h with chemical inhibitors SB203580HCL, PD98059, and JNK inhibitor II (30 µm/mL each), which are specific inhibitors for p38-MAPK, ERK-MAPK, and JNK-MAPK pathways, respectively. Total RNA was extracted from MSCs with TRIZOL reagent (Invitrogen, Carlsbad, CA) and reverse-transcribed by using mRNA Selective PCR Kit (TaKaRa). RT-PCR was routinely performed. The RANKL, OPG, IL-6, and M-CSF cDNA were also amplified by real-time PCR with the SYBR Green PCR kit (Sigma, St. Louis, MO). Primer sequences used for semiquantitative PCR were as follows: RANKL: forward, 5′-CTCTGTT CCTGTACTTTCGAGC-3′; reverse, 5′-AGTTTTTCGTGC TCCCTCCTTTC-3′. OPG: forward, 5′-TGATGAGTGTGTGTATTGCAGC-3′; reverse, 5′-TCTCTACACTCTCGGCATTC-3′. M-CSF: forward, 5′-CTGCTGCTGGTCTGTCTC-3′; reverse, 5′-GGTGAAGCAGCTGTTCAG-3′. IL-6: forward, 5′-CATCCAGTTGCCTTCTTG-3′; reverse, 5′-TCAAT AGGCAAATTTCCTG-3′. GAPDH: forward, 5′-AAACCCATCACCATCTTCCA-3′; reverse, 5′-GTGGTTCACACCCA TCACAA-3′. Primers used for real-time PCR were as follows: RANKL: forward, 5′-TGGAAGGCTCATGGTTGGAT-3′; reverse, 5′-CATTGATGGTGAGGTGTGCAA-3′. OPG: forward, 5′-AGCTGCTGAAGCTGTGGAA-3′; reverse, 5′-TGTTCGAGTGGCCGAGAT-3′. β-Actin: forward, 5′-GGACCTGACGGACTACCTCATG-3′; reverse, 5′-TCTTTGATGTCACGCACGATTT-3′. Real-time primers of IL-6 and M-CSF are the same as primers used for semiquantitative PCR.
ELISA assay for mouse OPG and human TNF-α
MSCs were seeded in 24-well plates at a density of 8 × 104/cm2 and incubated overnight. Media were removed and replaced with serum-free α-MEM for a minimum of 6 h before stimulation with graded concentration of TNF-α (5, 10, 50, and 100 ng/mL) for 8 h. The culture supernatants were collected with centrifugation and filtered to deplete cellular components. Concentration of mouse OPG in supernatants and human TNF-α in RASF samples were determined according to the reagent protocols of the quantitative determination kit of mouse OPG (R&D Systems, Minneapolis, MN: MOP00) and the human TNF-α immunoassay kit (R&D Systems: DTA00C), respectively. Optical density was read at 450 nm. For each experimental culture well, duplicate ELISA readings were obtained.
Western blot
MSCs were plated in 6-well plates at a density of 1 × 105/cm2 and incubated overnight. The cells were starved in serum-free α-MEM for at least 6 h before stimulated with TNF-α (100 ng/mL) for 0, 5, 10, 30, and 60 min. MSC samples were harvested by cells slicker on ice and lysis buffer (Biorad, Hercules, CA) was added. Thawed lysates were vortexed and centrifuged. Protein concentrations of lysates were determined by using the BCA Protein Assay (Pierce). Proteins were separated on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto nitrocellulose membranes and blocked by incubating the membranes with 5% wt/vol nonfat dry milk. Membranes were incubated with anti-p38 or anti-phosphor-p38, anti-ERK or anti-phosphor-ERK, anti-JNK or anti-phosphor-JNK mAb (Cell Signaling, Beverly, MA), and β-actin (Sigma) in the appropriate dilution overnight at 4°C. After incubation, membranes were rinsed with distilled water and washed 3 times in wash buffer TBST. Secondary antibody conjugated to horseradish peroxidase was added to the membranes in 5% nonfat dry milk in TBST. After 1 h of incubation at room temperature, membranes were washed 3 times with TBST and developed by using an enhanced chemiluminescence kit (Pierce, Rockford, IL).
Statistical analysis
Data were presented as the mean ± standard deviation and analyzed with Student’s t-test. A P value <0.05 was considered statistically significant.
Results
MSCs promoted CD11b+ monocytes to differentiate into osteoclasts
It is well known that osteoclasts are originated from monocyte–macrophage lineage. To observe if MSCs are involved in generation of osteoclasts, CD11b+ cells at purity of >90% (Fig. 1A) were co-cultured with MSCs and the formation of osteoclasts was evaluated with TRAP assay. At day 12, MSC co-culture system yielded ∼2–7 TRAP+-multinucleated cells (TRAP+ MNCs, ≥3 nuclei)/103 CD11b+ monocytes, while no TRAP+ MNCs were found in basal medium (Fig. 1B upper row, C). This stimulatory effect was MSC dose-dependent. Further, MSCs were added to osteoclast culture medium (with M-CSF and RANKL). At day 6, number of TRAP+ MNCs increased 2–3 times even at a relative low cytokines dose (M-CSF 10 ng/mL, RANKL 10 ng/mL) (Fig. 1B down row, D). Moreover, ratio of TRAP+ MNCs/total TRAP+ cells was also increased from 19% to 39% with MSC addition (Fig. 1E). To observe if MSCs could promote the survival of osteoclasts, CD11b+ monocytes cultured for 9 days in the presence of M-CSF and RANKL were co-cultured with MSCs without cytokines addition for another 6 days. The results showed that in the presence of MSCs, the number of survived TRAP+ MNCs was almost 2 times more than that in the absence of MSCs (Fig. 1F). Therefore, the data suggest that MSCs promote osteoclast development, maturation, and its survival.
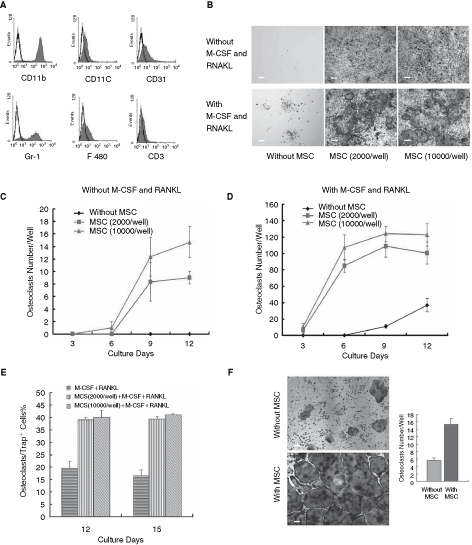
Mesenchymal stem cells (MSCs) exhibited positive modulation effect on the differentiation, maturation, and survival of osteoclasts. CD11b+ monocyte-sorting efficiency was analyzed by flow cytometry (
TNF–α-stimulated MSCs suppressed CD11b+ monocytes to differentiate into osteoclasts
TNF-α is a well-known proinflammatory cytokine and proved to promote osteoclastogenesis [26,27,35 –37], thus it was supposed that TNF-α might strengthen the supportive effect of MSC on osteoclast formation. However, pretreated by TNF-α or cultured in the presence of this cytokine, MSCs were observed to inhibit osteoclasts generated from monocytes (Fig. 2A). First, MSCs (1 × 104/cm2) were preincubated with TNF-α in graded concentration (5, 10, 50, and 100 ng/mL) for 24 h followed by addition of monocytes (1 × 104/cm2) with M-CSF (10 ng/mL) plus RANKL (10 ng/mL) or without the cytokines. Meanwhile, parallel experiments of MSCs and monocytes in the presence of TNF-α and with/without M-CSF and RANKL were also performed to investigate the direct influence of TNF-α on the preosteoclasts effect in presence of MSCs. After cultured for 9 days, cells were stained and TRAP+ MNCs were counted. It was observed that with TNF-α pretreatment or in the presence of TNF-α, MSCs failed to promote osteoclast generation (Fig. 2B). Moreover, they even inhibited osteoclast formation in osteoclast induction medium in a TNF-α dose-dependent manner (Fig. 2C). In addition, the inhibitory effect was more remarkable in co-culture experiment with direct supplementation of TNF-α. As shown in Figure 2C, pretreated MSCs caused a decrease in the number of osteoclasts for >70% when pretreated TNF-α dose reached 50 ng/mL while similar effect was achieved when supplemented TNF-α was 10 ng/mL. Second, it was explored if this effect was associated with TNF-α treatment time period. MSCs (1 × 104/cm2) were preincubated with TNF-α (100 ng/mL) for different time (4, 8, 12, 24, and 48 h) before co-cultured with monocytes (1 × 104/cm2) in the presence of M-CSF (10 ng/mL) plus RANKL (10 ng/mL). After a 9-day culture, we found that prolonged TNF-α treatment on MSCs also strengthened the inhibitory effect on osteoclast generation (Fig. 2D). Third, TNF-α–stimulated MSCs inhibited osteoclast formation at a lower MSCs/monocytes ratio. As shown in Figure 2E, a MSCs/monocytes ratio (1:5) began to prevent monocytes from differentiating to osteoclasts. As the ratio was raised to 10:1, MSCs were able to completely block osteoclast generation in the presence of RANKL and M-CSF. Most importantly, the suppression on osteoclast generation by MSCs was a lasting effect. As viewed in Figure 2F and G, the inhibitory effect maintained at least 12 days no matter in the presence or absence of M-CSF and RANKL.

Tumor necrosis factor (TNF)-α–stimulated mesenchymal stem cells (MSCs) suppressed the differentiation of CD11b+ monocytes into osteoclasts. Upon pretreatment by TNF-α or in the presence of TNF-α, MSCs suppressed CD11b+ monocytes to differentiate into osteoclasts (TRAP+, ≥3 nuclei) alone (
MSCs and TNF-α–stimulated MSCs exhibited opposite roles on resorbing function of osteoclasts
To examine the possible influence of MSCs on osteoclast-resorbing function, pit formation assay was performed. It was found that monocytes cultured with M-CSF and RANKL produced smaller resorption pits on slices of elephant tusk dentine (Fig. 3A and B) compared with monocytes co-cultured with MSCs in the presence of M-CSF and RANKL (Fig. 3A and B). No resorption pits were observed when monocytes were co-cultured with TNF-α–stimulated MSCs or with direct supplementation of TNF-α to co-culture system (Fig. 3A and B). Thus, it is suggested that MSCs exhibit opposite modulatory function not only on differentiation of osteoclasts but also on their bone resorption.
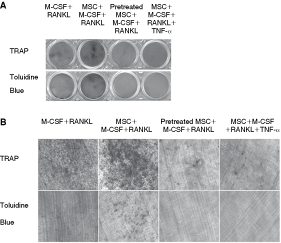
Mesenchymal stem cells (MSCs) and tumor necrosis factor (TNF)-α–stimulated MSCs exhibited opposite role on resorbing function of osteoclasts. In the presence of M-CSF (20 ng/mL) and RANKL (20 ng/mL), monocytes co-cultured with TNF-α–stimulated MSCs or with direct supplementation of TNF-α (100 ng/mL) to co-culture system failed to produce resorption pits on dentine slice (
TNF-α-stimulated MSC suppressed osteoclast generation partially through soluble factors
To search the mechanisms underlying the inhibitory effect of TNF–α-stimulated MSCs, transwell assay was performed. As shown in Figure 4A and B, MSCs strongly suppressed the differentiation of monocytes into osteoclasts in the transwell chamber system in the presence of TNF-α or pretreated by this cytokine. Also, unstimulated MSCs exhibit a remarkable boost on osteoclast formation without cellular contact. Thus, the data indicated that MSCs and TNF-α–stimulated MSCs regulated osteoclast formation mainly through soluble factors.
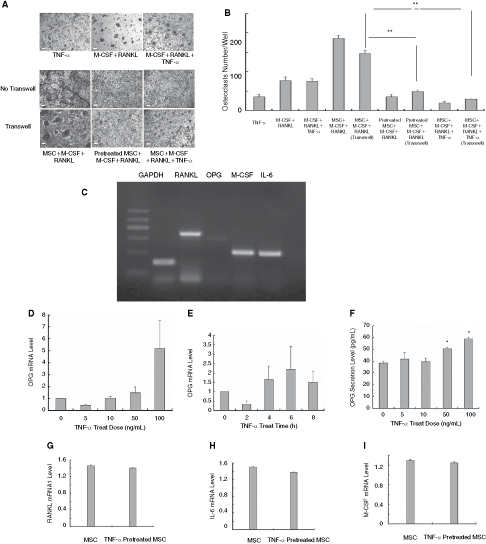
The suppressive modulation on monocyte-derived osteoclasts partially due to the increase of osteoprotegerin (OPG) by mesenchymal stem cells (MSCs). Unstimulated MSCs and tumor necrosis factor (TNF)-α–stimulated MSCs regulated osteoclast generation partially by secreting soluble factors shown by transwell assay (
TNF–α stimulation promoted the expression of OPG mRNA and the secretion of OPG protein by MSCs
To further clarify the complexity earlier, the mRNA levels of several cytokines in MSCs were observed. Semiquantitative PCR results revealed that unstimulated MSCs expressed RANKL, M-CSF, and IL-6 mRNA at higher levels compared with OPG mRNA (Fig. 4C). We then determined whether there was any change of OPG gene expression in MSCs in response to TNF-α. As shown in Figure 4D and E, TNF-α was capable of up-regulating OPG gene expression in MSCs in a time- and dose-dependent manner. Consistently, ELISA results showed an elevation of OPG secretion in a TNF-α dose-dependent manner (Fig. 4F). Meanwhile, real-time PCR results showed that TNF-α–stimulated MSCs down-regulated the expression levels of RANKL, IL-6, and M-CSF. However, no significant differences were observed (Fig. 4G–I).
TNF-α–stimulated MSCs up-regulated OPG and suppressed osteoclast generation via activation of MAPK pathway
In order to analyze the mechanism closely related to TNF-α–induced OPG production in MSCs, we evaluated the activation of MAPK pathway by TNF-α as it represents an inflammatory signaling. As shown in Figure 5A, TNF-α (100 ng/mL) induced the phosphorylation of p38-MAPK, ERK-MAPK, and JNK-MAPK. We further studied the effect of the specific MAPK inhibitors (SB23580HCL, PD98059, and JNK inhibitor II) on OPG mRNA expression. TNF-α–induced OPG mRNA elevation was partially blocked by SB23580HCL (Fig. 5C) and JNK inhibitor II (Fig. 5D) pretreatment. In contrast, pretreatment with PD98059 did not significantly prevent TNF-α–induced OPG mRNA up-regulation (Fig. 5E). Consistent with their roles in OPG mRNA, SB23580HCL (Fig. 5B, low row left; F) and JNK inhibitor II (Fig. 5B, low row middle; G) pretreatment for 1 h before TNF-α–stimulated MSCs rescued the osteoclast formation (P < 0.05 by t-test) while PD98059 (Fig. 5B, low row right; H) exhibited little effect on osteoclastogenesis with statistical significance. Therefore, the results indicated the involvement of p38/MAPK and JNK/MAPK in the inhibitory effect on osteoclasts by MSCs.
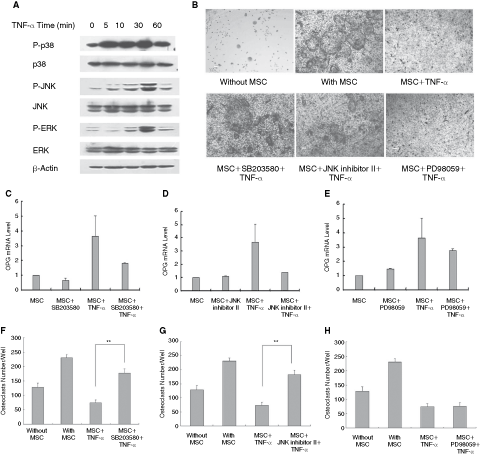
TNF-α–stimulated mesenchymal stem cells (MSCs) increased osteoprotegerin (OPG) expression and suppressed osteoclast generation via activation of MAPK pathway. Tumor necrosis factor-α (TNF-α) was observed to be capable of activating p38, ERK, and JNK/MAPK in MSCs (
TNF-α–activated MSCs retained immunophenotyping characteristics, multipotential differentiation capacities, and immunosuppressive properties
Based on our previous observations, TNF-α–pretreated MSCs were identified as osteoclast inhibitors. However, it remains unknown whether multipotential differentiation capacity and immunomodulatory property of MSCs are changed after TNF-α treatment. It is observed (data not shown) that the cells highly expressed Sca-1 (a murine hematopoietic and mesenchymal stem/progenitor cell marker), CD105 (endoglin), CD29 (β1-integrin), and CD44 (receptor for hyaluronate and osteopontin). Meanwhile, they were homogeneously negative for CD45 (pan-hematopoietic marker), CD31 (endothelial cell marker), MHC-la (MHC-II molecular), and CD86, demonstrating that TNF-α treatment did not change the immunophenotyping characteristics of MSCs.
To confirm the multilineage differentiation capacity of TNF-α–stimulated MSCs, the cells were maintained in osteoinductive and adipoinductive media. Histological staining showed that no difference of differentiation capacity was observed between MSCs and TNF-α–treated MSCs (Fig. 6A).
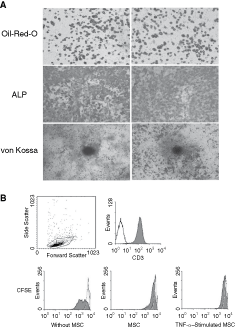
Tumor necrosis factor (TNF)-α–activated mesenchymal stem cells (MSCs) remain certain immunophenotyping characteristics, multipotential differentiation capacities, and immunosuppressive property. TNF-α–stimulated MSCs differentiated to adipocytes, osteoblasts, and osteocytes, assessed by Oil Red O, ALP, and von Kossa staining, respectively (
It is well documented that MSC suppress T-cell activation stimulated by nonspecific mitogens. To observe whether TNF-α–activated MSCs displayed similar features, CD3+ T lymphocytes from the same strain (C57BL/6) were loaded with CFSE and then co-cultured with TNF-α–activated MSCs at a MSCs/T-lymphocytes ratio (1:10) in the presence of Con A (2 µg/mL). T lymphocytes were harvested at days 2 and 4. Consistent with unstimulated MSCs, TNF-α–treated MSCs remarkably inhibited Con A-stimulated T-lymphocytes proliferation (Fig. 6B). Therefore, it is demonstrated that in our culture condition TNF-α did not change the inherent properties of MSCs.
RASF treatment changed the modulatory effect on osteoclasts by MSCs
To observe the potential role of MSCs in osteoclastogenesis in inflammatory conditions, MSCs were treated with RASF, in which the concentrations of TNF-α were detected by ELISA (Fig. 7). It was observed that MSCs promotes osteoclast formation when TNF-α concentration was relatively low in samples, while they inhibited osteoclast generation after treated by samples with high TNF-α concentration. As shown in Figure 7, 11 of total 20 RASF samples (samples 4, 5, 7, 8, 11, 12, 13, 14, 16, 18, and 20) dampened the supportive property of MSCs alone on osteoclasts. However, only 7 of 20 RASF samples (samples 5, 7, 8, 11, 12, 16, and 17)-stimulated MSCs inhibited osteoclast formation in the presence of RANKL and M-CSF (Fig. 7). Thus, the data implied the possible link between osteoclast formation and MSCs in RA joint microenvironment.

Rheumatoid arthritis synovial fluid (RASF) treatment changed mesenchymal stem cell (MSC) modulatory capacity on osteoclasts. The tumor necrosis factor (TNF)-α level in 20 RASF samples was assayed by ELISA. As shown in Figure 7, 11 of 20 total patients’ RASF (at 10% addition) (samples 4, 5, 7, 8, 11, 12, 13, 14, 16, 18, and 20) weakened the independent supportive property of MSCs on osteoclasts. However, only 7 of 20 samples (samples 5, 7, 8, 11, 12, 16, and 17) of stimulated MSCs inhibited osteoclast generation in the presence of RANKL and M-CSF.
Discussion
In this study, we showed that mouse MSCs exhibited dual regulatory effect on differentiation and bone-resorbing function of monocyte-derived osteoclasts. Although both MSCs and osteoclasts reside in bone marrow cavity, they have traditionally been viewed as distinct unrelated cells until Mbalaviele et al. [38] reported human bone marrow MSCs supporting CD34+ hematopoietic stem/progenitor cell differentiate to osteoclasts in vitro. However, osteoclast differentiation is greatly dependent on environmental factors. Herein, we demonstrated that MSCs promoted CD11b+ monocytes differentiating to TRAP+-multinucleated osteoclasts in the absence or presence of RANKL and M-CSF. Strikingly, we observed that upon stimulation with TNF-α, MSCs turned to strongly inhibit the initial differentiation of monocytes into osteoclasts even in the presence of RANKL and M-CSF. This observation indicates that MSCs indeed play a dual role in osteoclast formation instead of a simple positive modulator.
In fact, the promoting effect on osteoclasts is in line with previous findings that MSCs act as hematopoiesis supporter [8]. As one of the cells to create the HSC niche, MSCs supply numerous cytokines and cell surface signals in supporting HSC survival, proliferation, and maintenance. M-CSF is constitutively released by MSCs and induces monocytes to differentiate into macrophages, which proved to be potential osteoclast precursors [39]. Moreover, MSCs secrete high level of the RANKL [24,25] and IL-6 [40], all of which might be attributed to this observation.
In addition, previous reports have shown that osteoclast development requires direct cell–cell interactions of the hematopoietic osteoclast precursors with the neighboring osteoblast/stromal cells [41]. Similarly, in our study, the osteoclasts generated in transwell chamber system slightly decreased indicating that cell surface proteins might affect osteoclast formation by MSCs. Therefore, we suggest that MSCs exhibit positive effect on osteoclastogenesis not only by secreting a cocktail of higher amount pro-osteoclastogenesis cytokines but also by direct cell–cell contact.
TNF-α is one of the most prominent osteoclastogenic cytokines produced in inflammation. It directly induced osteoclasts formation in vitro in the presence of M-CSF [42], stimulates osteoclast differentiation by a mechanism independent of the RANKL–RANK interaction [27], and induces osteoclastogenesis by direct stimulation of macrophages exposed to permissive levels of RANKL [43]. However, our data proved that MSC modulatory effect on osteoclasts changed upon stimulation with TNF-α and this alteration seemed partially dependent upon OPG, a decoy receptor for RANKL. Similarly, previous reports have shown that other cytokines including transforming growth factor-β1, interleukin-4, and interleukin-1β are able to up-regulate OPG expression in marrow stromal cells ST2 [44], osteoblastic cells MC3T3-E1 [44], and endothelial cells [45]. Also, it has been documented that TNF-α can induce OPG expression in osteoblast-like MG-63 cells [46]. Thus, it is conceivable that MSCs, the precursors of stromal cells including endothelial cells and osteoblasts, might possess the same reaction of up-regulating OPG in certain cytokine microenvironment.
To search the mechanism underlying the suppressive effect of TNF-α–treated MSCs, MAPK pathway was chosen to investigate as it has been reported to be involved in OPG regulation in stromal cells [46 –48]. Consistently, we observed that p38-MAPK and JNK-MAPK activation was indispensable for OPG elevation by MSCs.
In addition to the change in expression of OPG, other factors including cell surface proteins might affect osteoclast formation. As viewed in Figure 4 G–I, TNF-α–stimulated MSCs slightly down-regulated the mRNA expression of RANKL, IL-6, and M-CSF. Moreover, the cells contact factor might also attribute to the inhibitory effect as a slight recovery of osteoclasts was observed when MSCs and monocytes were separated in transwell assay. Furthermore, the direct TNF-α supplementation to MSC/monocytes cell culture yield strengthened suppressive effect. The observation suggests that MSC might respond to chronic low-level TNF-α in inflammatory microenvironment. Therefore, it is likely that the effects of MSCs on osteoclasts are complex and will depend on specific conditions. These results suggest that both cytokines and cell surface molecules are involved in the alteration and appeal further detailed investigations to deepen our understanding in the modulatory effects of MSCs.
Currently, the rising interest in MSC-based therapies comes from their plasticity, their immunoregulatory activity, their capacity to engraft and survive long-term in the appropriate target organs, and their contribution to the repair of injured tissues. Therefore, in this investigation the multipotential differentiation capacity and immunomodulatory properties of TNF-α–activated MSCs were highly concerned. After pretreated with TNF-α (100 ng/mL) for 24 h, MSCs were induced to osteoblast and adipocyte to examine if there is any impairment on the multipotential differentiation property. In our culture condition, no difference between unstimulated MSCs and TNF-α–stimulated MSCs was observed. Also, stimulated MSCs were capable of suppressing lymphocyte cell proliferation triggered by Con A in vitro. Further examination of immunophenotype proved that they still occupied a typical MSC antigen profile.
Though MSCs acted as a conditional osteoclastogenesis inhibitor after stimulated by TNF-α, their potential role in complicated inflammatory microenvironment remained to be determined. RA is a chronic inflammatory disease that involves T-lymphocytes and osteoclast activation in pathologic process and results in cartilage and bone destruction at the late phase. TNF-α is a well-known proinflammatory cytokine that plays a pivotal role in RA and antibodies against TNF-α have been successfully used in rodent models with arthritis and in clinical trials [49]. Synovial fluid from RA patients reflected the cytokines microenvironment of the disease and an induced bone resorption enhancement depends on TNF-α and other uncharacterized soluble factors [34]. Therefore, we used RASF to mimic the inflammatory microenvironment and to discriminate the potential roles of MSCs in osteoclastogenesis. Our data showed that MSCs modulated osteoclast generation in close relation with the TNF-α level in RASF. However, the occurrence of suppressive effect was not necessarily correlated with the dose of TNF-α in some patients. The results suggest that OPG secreted by TNF-α–activated MSCs might be counterbalanced by high dose of RANKL in osteoclast induction medium. Moreover, it is possible that other proinflammatory cytokines in RASF may regulate MSCs via unknown mechanism and exhibit different effect on osteoclasts. Also, the controversial results of MSC application in the management of arthritis [21,22] suggest that further studies are required to understand the complicated role of MSCs under proinflammatory microenvironment.
In summary, our study reveals that MSCs possess dual modulatory function in osteoclast differentiation and bone resorption, and MSCs may serve as promising candidates to improve osteoclast-associated disorders.
Footnotes
Acknowledgments
We sincerely thank Dr. Sheng Zhou and Dr. Chen-Jie Fang for critical reviewing of the manuscript. This study was supported by the National Key Basic Research Program of China (2005CB522705), the National Natural Science Foundation (30600309 and 30730043), and High-tech Research and Development Program of China (2007AA021109, 2007AA02Z454).
Author Disclosure Statement
The authors declare no competing financial interests.