
Abstract
Human umbilical cord blood mononuclear cells (HUCB) have been shown to have a therapeutic role in different models of central nervous system (CNS) damage, including stroke. We evaluated the possible therapeutic potential of HUCB in P7 rats submitted to the Rice-Vannucci model of neonatal hypoxic–ischemic (HI) brain damage. Our results demonstrated that intraperitoneal transplantation of HUCB, 3 h after the HI insult, resulted in better performance in two developmental sensorimotor reflexes, in the first week after the injury. We also showed a neuroprotective effect in the striatum, and a decrease in the number of activated microglial cells in the cerebral cortex of treated animals. We suggest that HUCB transplantation might rescue striatal neurons from cell death after a neonatal HI injury resulting in better functional recovery.
Introduction
N
Human umbilical cord blood (HUCB) is a source of hematopoietic stem/progenitor cells, endothelial progenitors, and mesenchymal stem/progenitors. Since 1989, when the first successful umbilical cord transplantation was reported in a child with Fanconi anemia, thousands of transplants have been carried out, as a therapy for certain malignant and nonmalignant hematological disorders [3]. Moreover, in the last few years, preclinical studies have shown that HUCB injected systemically in the acute phase of animal models of stroke have a therapeutic effect. These cells can reduce the area of brain infarction [4] and the inflammation [5], and increase the regenerative capacity of the brain [6], improving behavioral recovery [7].
However, the mechanisms involved in neuronal cell death after ischemic lesions differ in the newborn and the adult brain, with age-dependent differences in mitochondrial responses to injury [8], and increase activation of caspase-3 in the immature than in the juvenile or the adult brain. In the former, there is also an increased secondary loss of tissue caused by maturational and trophic disturbances related to the primary loss of tissues that are important for the early postnatal development of the brain [9].
The potential of HUCB to reduce the neurologic deficits associated with neonatal hypoxia–ischemia was shown in rats in a study that assessed the functional benefits of these cells, using a walking-pattern analysis test. The HUCB cells, when injected intraperitoneally 24 h after the insult, migrated to areas of brain damage in large numbers, and alleviated the spastic paresis of the animals [10]. However, the mechanisms involved in this improvement were not investigated.
In our study, we injected HUCB into newborn rats after the induction of HI brain damage in order to evaluate the possible therapeutic role of these cells and the mechanisms involved in this effect. Our results demonstrated that transplantation of HUCB cells resulted in a better performance in two of three developmental sensorimotor reflexes after the HI insult. We also showed that in the treated group, there was a reduction in cell death in the striatum and a decrease in the number of activated microglial cells in the cerebral cortex. We suggest that HUCB cells have a neuroprotective role in the HI model, and as a consequence, the treated animals have better functional recovery than the untreated group.
Materials and Methods
HUCB sources and preparation
Human umbilical cord blood was collected from full-term newborns in bags containing citrate phosphate dextrose as an anticoagulant (JP Indústria Farmacêutica, Brazil). All the mothers who donated cord blood gave their informed consent. After centrifugation at 1,200g for 30 min at 4°C, the mononuclear cells were isolated by a Ficoll density gradient, washed with 0.1 mol/L PBS, resuspended in bovine fetal serum with 10% DMSO (Merck, Darmstadt, Germany), and frozen in liquid nitrogen until the injection.
Neonatal hypoxic–ischemic brain damage
Protocols were approved by the animal care committee of the Federal University of Rio de Janeiro. At postnatal day 7 (P7), male Lister-Hooded rat pups were submitted to HI following the Rice-Vannucci model of neonatal HI brain damage [2]. In this model, P7 pups had the right common carotid artery ligated using 5–0 silk suture under sevoflurane anesthesia (Sevorane, Abbott, São Paulo, Brazil). After a 2-h recovery period, pups were placed in a hypoxia chamber with a flow of O2 (8%) and N2 (92%) for 90 min. The hypoxia chamber was kept in a temperature-controlled water bath, to maintain the temperature inside the chamber at 37°C. Sham controls underwent anesthesia and incision only. The pups were returned to their dams, and the litter size was adjusted to 6–8 pups per dam. The number of animals used for each experiment is shown in Table 1.
T
aCresil violet staining and immunohistochemistry (anti-human nuclei) were done in tissue sections from these animals. We have not used sham-operated animals for the histological analysis, since the contralateral hemisphere was used as a control.
Abbreviation: HUCB, human umbilical cord blood mononuclear cells.
Transplantation
Cryopreserved HUCB were rapidly thawed at 37°C and resuspended in DMEM/F12 (Invitrogen, Carlsbad, CA, USA). Nucleated HUCB were counted with a cytometer to ensure adequate cell numbers for transplantation. Viability was determined by the Trypan blue dye exclusion method. The final dilution was 2 × 106 cells in 200 µL DMEM for intraperitoneal injection, 3 h after the hypoxia, in the treated group (HI + HUCB). The untreated group (HI + vehicle) received an intraperitoneal injection of 200 µL DMEM. Thus, we had three experimental groups: sham, HI + vehicle, and HI + HUCB. In order to label the cells, HUCB were incubated with CellTrace Far Red DDAO-SE (Invitrogen) for 30 min, at 37°C, before they were injected into three animals of the HI+HUCB group.
Neurological outcome
Three developmental sensorimotor reflexes were evaluated, and reflex performance was compared among the groups. The reflexes were evaluated at the indicated times:
a. Cliff aversion reflex: Animals were placed with their forepaws overhanging the edge of a board. The time required (in seconds) to turn away from the edge (90°) was recorded at 2, 4, and 7 days after the injury.
b. Negative geotaxis reflex: Pups were placed head downward on an inclined board (35°) 30 cm long. The hind limbs of the pups were placed in the middle of the board. The time (in seconds) required to rotate their bodies head-up (180°) was recorded at 2, 4, and 7 days after the injury.
c. Gait: Animals were placed in the center of a white paper circle 13 cm in diameter. The time (in seconds) that they took to place both forelimbs outside the circle was recorded. Since none of the animals moved from the center of the circle at P9, we evaluated this test at 4, 7, and 10 days after the injury.
Pups were given 5 min of rest between each reflex assessment. If the animal was unable to perform the reflex within 60 s, the maximum time was assigned.
Histological and immunohistochemical analysis
Animals were allowed to survive for 2 or 7 days after HI, and they were then reanesthetized with sevoflurane. Animals were fixed by transcardiac perfusion with a solution of 4% paraformaldehyde in 0.1 M phosphate-buffered saline, and the brains were removed. After cryoprotection, 14-µm thick coronal sections were cut, with the use of a cryostat.
After blocking with 5% normal goat serum in phosphate-buffered saline for 30 min, sections were incubated with primary antibody at 4°C overnight, followed by the appropriate secondary antibody for 2 h at room temperature.
Primary antibodies (dilution and supplier in parentheses) were directed against cleaved caspase-3 (1:100; ABCAM, Cambridge, MA, USA), CD68 (ED1; 1:100; Serotec, Dusseldorf, Germany), glial fibrillary acidic protein (GFAP; polyclonal: 1:500; Dako, Carpinteria, CA, USA), and human nuclei (1:100; Chemicon International, Temecula, CA, USA).
Secondary antibodies were Alexa Fluor 488-conjugated goat anti-rabbit (1:200; Invitrogen), Cy3-conjugated goat anti-rat IgG (1:1,000; Jackson, West Grove, PA, USA), or Cy3-conjugated goat anti-rabbit (1:800; Jackson). Sections were mounted using Vectashield (Vector, Burlingame, CA, USA).
For assessment of tissue damage, three sections, starting from the level of the lateral ventricle, 154 µm from each other, were stained with Fluoro-Jade C or immunostained for ED1. Fluoro-Jade C stain mainly identifies degenerating neurons and it was done as previously described by Leonardo et al. [11]. The number of stained cells in one field/section was counted, under 20× magnification, in the striatum and in the parietal cortex, for Fluoro-Jade C (2 days after the injury); and in the striatum, cerebral cortex, dorsolateral subventricular zone, and external capsule for ED1 (7 days after the injury). Three other sections were immunostained for activated caspase-3 (2 days after the injury). The number of cells expressing activated caspase-3 was counted in one field of the parietal cortex, under 40× magnification, in a 7.68-µm thick confocal reconstruction. In the striatum, the cells were counted under 20× magnification, in a 9.25 µm confocal reconstruction.
Fluorescence was documented using confocal imaging microscopy (Zeiss LSM 510 META; Carl Zeiss, Gottingen, Germany) or Apotome microscopy (Carl Zeiss).
Statistical analysis
Statistical comparisons among groups were determined using the Mann–Whitney test for the histological analysis. For the functional outcome analysis, statistical comparisons were made using one-way ANOVA with Bonferroni’s posttest. P < 0.05 was considered statistically significant. All analyses were performed using GraphPad Prism version 4.02 for Windows (GraphPad Software, San Diego, CA, USA).
Results
Rice-Vannucci model of neonatal HI resulted in extensive brain damage
Combining permanent occlusion of the right common carotid artery with 90 min of hypoxia (92% N2 + 8% O2) resulted in moderate to severe brain damage in P7 male pups. We chose to work only with males because recent studies have indicated differences between the sexes in susceptibility to HI [12]. Only 2 days after the HI insult, we observed, in cresyl violet-stained sections, areas of brain damage in the hemisphere ipsilateral to the ischemia, including the cerebral cortex and the striatum (Fig. 1A). Seven days after the insult, the damage evolved to an atrophy of these areas. At this time there was an ex vacuum ventriculomegaly in the ipsilateral hemisphere, and porencephalic cysts were formed (Fig. 1B). Immunostaining for GFAP revealed that these cysts were surrounded by reactive astrocytes (Fig. 1C).
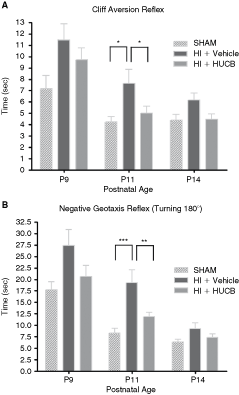
Human umbilical cord blood mononuclear cells (HUCB) transplantation resulted in a better functional outcome after neonatal hypoxic–ischemic (HI) brain injury. To evaluate the neurofunctional outcome, we tested the cliff aversion and negative geotaxis reflexes at 2, 4, and 7 days after HI insult. At all ages examined, the HI + vehicle group (n = 25) showed a retarded performance in cliff aversion (
HUCB transplantation resulted in a better functional outcome after neonatal HI brain injury
Negative geotaxis reflex. In this reflex, we recorded the time that the animals took to turn 180°, rotating their bodies head-up. At P9, P11, and P14, HI + vehicle animals needed more time to perform this reflex, when compared to sham-operated animals (Fig. 2B). At P11 this difference was significant (P < 0.001). HI + HUCB animals tended to perform this reflex faster than HI-vehicle animals at P9, P11, and P14 (Fig. 2B). At P11 this difference was significant (P < 0.01).
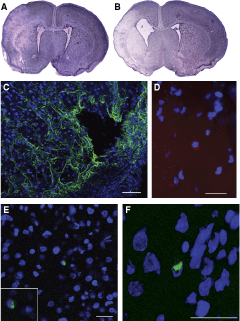
HI brain damage and engraftment of HUCB. (
Cliff aversion reflex. The HI + vehicle group required the longest time to perform this test when compared with the other two groups, at all ages examined (Fig. 2A). At P11, this difference was significant (P < 0.05). HI-HUCB animals showed a delay in performing this reflex at P9. However, at P11, HI-HUCB animals performed similarly to the sham animals, with a significant difference from the HI-vehicle group (Fig. 2A; P < 0.05).
Gait. The time taken to move off a circle of 13 cm in diameter did not differ among the groups: all the animals spent the same time to move off the circle, at all ages examined (data not shown).
These results indicate that HI brain injury retards the development of the cliff aversion and negative geotaxis reflexes, and that HUCB transplantation can prevent this impairment. The gait reflex is not altered after a moderate to severe HI insult.
HUCB transplantation had a neuroprotective effect in the striatum
In vitro studies have demonstrated that HUCB cells secrete many neurotrophic factors, some of them involved in neuroprotection [13]. In order to assess whether HUBC transplantation was able to exert a neuroprotective effect in vivo, we quantified the number of Fluoro-Jade C-positive cells (a marker for degenerating neurons) in the cerebral cortex and in the striatum 2 days after the HI insult when massive primary neuronal death occurred. In the HI + vehicle group, we found a large number of dying neurons in both areas, only in the hemisphere ipsilateral to the ischemia (Fig. 3A), with a staining pattern similar to what was previously shown by Leonardo et al. [11]. There was a significant decrease in the number of dying neurons in the ipsilateral striatum of the HI + HUCB group (Fig. 3B,E; P < 0.05), but no significant changes were found in the cortex of this group (data not shown). These observations indicate that HUCB are able to promote neuroprotection in a group of striatal neurons.
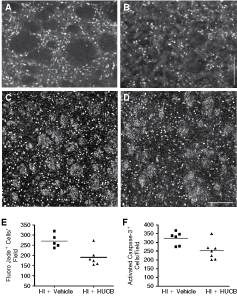
HUCB transplantation reduced cell death in the striatum. (
HUCB transplantation reduced activated caspase-3-mediated cell death in the striatum
Following neonatal HI in the rat, the predominant form of cell death is a continuum phenotype, with activation of apoptotic cascades, including caspase-3 activation, and mitochondrial structural and functional failure [14]. We counted the number of cells expressing activated caspase-3 in the striatum and in the cerebral cortex 2 days after the HI insult. In the ipsilateral hemisphere, we found numerous cells expressing activated caspase-3 in both areas, although these cells were rare in the contralateral hemisphere. Comparing HI + vehicle (Fig. 3C) and HI + HUCB animals (Fig. 3D), we found fewer cells expressing activated caspase-3 in the striatum of the HI+HUCB animals (Fig. 3F; P < 0.05). We also observed many axons expressing activated caspase-3 in the striatal white matter in both groups (Fig. 3C,D). In the cerebral cortex, we found no differences between the groups (data not shown). These observations indicate that HUCB treatment can protect a group of striatal cells from activated caspase-3-dependent cell death.
HUCB transplantation reduced microglial activation in the cerebral cortex
Although microglial cells have recently been shown to promote repair after a brain injury, an uncontrolled and extended inflammatory reaction is detrimental to the central nervous system (CNS) [15]. Because HUCB were shown to secrete some anti-inflammatory cytokines in vitro [16], we wanted to know if HUCB cell transplantation would have an effect on the activation of microglial cells in the subacute response of HI injury, when the persistent activation of these cells may be responsible for a secondary neuronal damage.
We quantified the number of cells expressing the CD68 (ED1) antigen, expressed by activated microglia and macrophages, in different brain areas 7 days after the insult. HI + vehicle animals had an increased number of ED1+ cells in the cerebral cortex ipsilateral to the ischemia (Fig. 4C). These cells tended to accumulate in areas of astrogliosis, as shown by GFAP (an astrocytic marker) expression (Fig. 4B). In these animals, we were also able to see the formation of porencephalic cysts in the cerebral cortex, which were surrounded by astrocytes and also by activated microglia, forming a wall around the cyst (Fig. 4E). The number of ED1+ cells in the ipsilateral cerebral cortex was significantly reduced in the HI + HUCB animals (Fig. 4D) when compared to the HI + vehicle group (Fig. 4C) (Fig. F; P < 0.001). In the contralateral hemisphere, we failed to find these cells in the cerebral cortex of both groups (Fig. 4A).
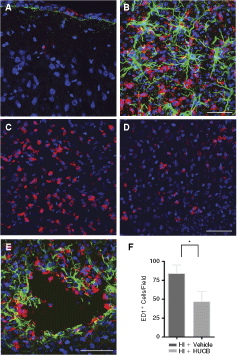
HUCB transplantation reduced microglial activation in the cortex. Coronal sections of neonatal rat brain tissue 7 days after the hypoxic–ischemic (HI) insult. We identified a large number of activated microglia (expressing the antigen ED1+, in red) and reactive astrocytes (expressing GFAP, in green) in the cortex ipsilateral to the ischemia (
We also quantified the number of ED1+ cells in the subventricular zone, striatum and in the external capsule, areas where neural stem/progenitor, neuronal, and oligodendroglial cell death, respectively, occur after an HI insult [17,18]. In these areas, we found an increase in the number of ED1+ cells in the ipsilateral hemisphere, compared to the contralateral hemisphere, in both groups. However, in these regions, there was no significant difference in the number of ED1+ cells between the treated and untreated groups (data not shown).
These results indicate that HUCB cell transplantation may have an anti-inflammatory effect due to the reduction in the number of activated microglial cells in the cerebral cortex ipsilateral to the ischemia, in the subacute phase after the HI brain damage.
Few HUCB migrated to areas of brain injury
In some animals, we labeled the cells with CellTrace in order to follow the cells after transplantation. Two days after the injection, we were able to see a few cells in the cerebral cortex (Fig. 1D) and in the striatum (data not shown) ipsilateral to the ischemia, but not in the contralateral hemisphere. Similar results were found when we used an anti-human nuclei antibody to identify the injected cells (Fig. 1E, F).
Discussion
Human umbilical cord blood is a source of hematopoietic, mesenchymal [19,20], and endothelial [21] stem cells and/or progenitors available for transplantation in the first hours after birth, allowing a possible rapid intervention, as soon as the diagnosis of a HI encephalopathy is made. Presently, in addition to its use for the treatment of hematological diseases, some preclinical studies have suggested a possible therapeutic effect of these cells in different models of CNS damage, including stroke, in adult animals [22]. In our study we investigated the possible therapeutic role of these cells in a model of neonatal HI brain injury.
In the newborn, there is a massive loss of neurons [23], as well as a huge inflammatory response [24] in the first 24 h after the HI insult, and for this reason we systemically injected the cells 3 h after the HI injury, in order to evaluate the possible beneficial effect of a rapid intervention. We have not injected the cells immediately after the injury because we think that cord blood cells, as a therapy, probably would be injected in the children a few hours after the HI insult, given the time needed to make the diagnosis and obtain the mononuclear cell fraction.
The maturation of neurological reflexes reflects the postnatal development of the CNS, and can be influenced by different factors. After a neonatal HI brain damage, some of these reflexes are affected in the first weeks after the insult [25], with a correlation between the degree of cerebral atrophy and the time taken to perform the reflexes [26]. In our study, the neurofunctional outcome was measured by three neonatal reflexes, from 2 to 10 days after the injury. Four days after the injury, cliff aversion and negative geotaxis reflexes were retarded in HI animals injected with the vehicle, although the gait reflex was not altered. At this time point, HI animals injected with HUCB performed better in these two reflexes similar to sham-operated animals. Cliff aversion and negative geotaxis were previously shown to closely correlate with long-term functional deficits, measured by the water-maze test [27]. Thus, our results indicate that HUCB transplantation is associated with a better functional outcome after an HI injury.
In order to analyze the mechanisms involved in this functional effect, we evaluated the possible neuroprotective role of HUCB transplantation, quantifying the number of neurons undergoing cell death in the cerebral cortex and striatum using Fluoro-Jade C as a marker. In the group that received therapy, we showed a reduction in the number of dying neurons in the striatum, 2 days after the HI insult. Fluoro-Jade C selectively labels neurons [28], and in order to investigate cell death from neurons and glial cells we used immunostaining for activated caspase-3 to identify cells undergoing death via this pathway. With this technique we demonstrated a decrease in cell death, in the treated group, in the striatum. However, with both techniques we were unable to find differences in the number of cells dying in the cortex. Together, these results suggest that only a subset of striatal neurons, probably those less susceptible to HI injury, could be rescued by the HUCB transplantation. Or, it could also suggest that HUCB-mediated mechanisms involved in neuroprotection after a neonatal HI injury are more efficient in the striatum. It was previously shown that GABAA and GABAB coactivation has neuroprotective effects against in vitro ischemia [29]. HUCB transplantation could, for example, promote neuroprotection by the release of a factor involved in the modulation of this activity in striatal neurons. In addition, we suggest that cell therapy results in a decrease in the number of striatal cells in which the caspase-3 death pathway was activated after HI. Because the basal ganglia show increased susceptibility to HI injury in the full-term infant [30], HUCB transplantation may protect these neurons from caspase-3-dependent cell death.
Microglia are the brain-resident immune cells. When activated by endogenous signals, such as neuronal death, or by pathogen components, these cells play an important role in the removal of toxic substances and debris creating an environment that allows regeneration. However, the brain has a poor ability to tolerate this response, since it involves the secretion of neurotoxic factors such as nitric oxide and reactive free radicals [15].
We showed that HUCB transplantation reduced the number of activated microglia and macrophages in the cerebral cortex ipsilateral to the ischemia 7 days after the injury. This anti-inflammatory effect may prevent the side effects associated with prolonged microglial activation and may be partially responsible for the better functional outcome that we observed after administration of HUCB. The failure to observe the same effect in other brain regions in which the number of activated microglia also increases after HI injury (external capsule, striatum, and SVZ) may be related to regional heterogeneity in the signals that control microglia activation. Since there are many “on” and “off” signals modulating the activity of these cells [31], our hypothesis is that the “off” signals directly or indirectly released after the transplantation were sufficient for microglial inactivation in the cerebral cortex but not in other areas. Alternatively, different regions might have different time periods for activation, and we investigated only the period of 7 days after the treatment.
The effects that we observed after the transplantation were not correlated with a significant engraftment of the injected cells in the brain. In this respect, a recent study has demonstrated that intraperitoneal injection of HUCB 24 h after HI injury results in a significant improvement in spastic paresis, and in these animals a large engraftment of the cells in the brain was also demonstrated with no differentiation into neurons or astrocytes [10]. In our study, we injected the cells 3 h after HI, and only a small number of the injected cells were found in the brain regions after 2 days. Our hypothesis is that at 3 h after the injury, when we injected the cells, the expression of chemokines involved in the migration of HUCB to the brain is still increasing [32 –34]. Our results are in accordance with those of Schwarting et al. [35], who showed an anti-inflammatory effect in the ischemic hemisphere of adult mice injected with bone marrow hematopoietic stem/progenitor cells before the cells were detected in the brain. They correlated this result with an early immunomodulatory effect in the spleen. HUCB are also able to modulate the stroke-induced inflammatory response, reducing leukocyte infiltration in the brain [5] and increasing the production of interleukin-10 (IL-10), an anti-inflammatory cytokine in the spleen [36].
Grafting of the transplanted cells at the lesion site might not be necessary for the functional beneficial effect, because it has been shown that HUCB induce neuroprotection in an animal model of stroke, without entering the CNS [37]. It is possible that circulating HUCB secrete several cytokines and growth factors when circulating in vessels adjacent to regions of neurodegeneration [38]. Supporting this hypothesis are several lines of evidence showing that HUCB express high levels of mRNA of many neurotrophic factors, including glial-derived neurotrophic factor (GDNF), brain-derived neurotrophic factor (BDNF), nerve growth factor (NGF), neurotrophin-3 (NT-3), and neurotrophin-4/5 (NT-4/5). BDNF and NT-4/5 were also found in culture supernatants of HUCB [13], as well as vascular endothelial growth factor (VEGF) [16], recently shown to be neuroprotective [39], and IL-10, an anti-inflammatory cytokine. Since BDNF is neuroprotective against neonatal HI brain injury in vivo, blocking caspase-3 activation [40], the secretion of this factor could be responsible for part of the effects we have observed. Moreover, BDNF, NT-3, and NGF are also considered “off” signals for microglia activation, restricting their function in the inflamed brain [31], and may account for our results.
Several questions should be addressed before the results from the preclinical studies are transferred from bench to clinic. For example, it is important to determine the best time window, as well as the ideal number of cells for therapy. It would also be interesting to test whether two or more injections at different times could improve the therapeutic effect, and, in addition, whether a synergic effect with other therapies could be obtained. It was shown, for example, that blood brain barrier permeabilization with mannitol enhances the therapeutic efficacy of systemically administered HUCB in a model of neonatal HI (41). Moreover, in recent years, one of the most successful neuroprotective approaches tested in both preclinical and clinical trials of HI encephalopathy is hypothermia [42], raising the question of whether it would be possible to save more neurons by combining hypothermia with HUCB cell therapy in the first hours after birth.
Footnotes
Acknowledgments
This work was supported by CNPq, CAPES, and FAPERJ. The authors wish to thank Dr. Lenir Orlandi and Prof. Dr. Carlos Alexandre Netto for their excellent intellectual support and Mr. Felipe Marins for technical assistance.
Part of this work was presented at the 38th Annual Meeting of the Society for Neuroscience (Pimentel-Coelho PM, ES Magalhaes, LM Lopes, LC deAzevedo, MF Santiago and R Mendez-Otero. (2008). Human umbilical cord blood cell transplantation in a model of neonatal hypoxic–ischemic encephalopathy. 38th Annual Meeting of the Society for Neuroscience, Washington, DC).
Author Disclosure Statement
No competing financial interests exist.