
Abstract
Cell transplantation-based therapy could be an effective way for the treatment of many diseases. Among numerous somatic stem cells isolated and purified, skin-derived precursors (SKPs) are abundant autologous cells, providing a large reservoir of cells for therapeutic transplantation. Previous studies showed that SKPs could be differentiated into neural and mesodermal progeny in vitro. In the present study, we attempted to differentiate SKPs to muscle progenitors in vitro. After treatment with a combination of growth factors, SKPs were differentiated into cells expressing markers of muscle progenitors, including Pax7. Furthermore, some of these cells expressed desmin, TnT, Mstn, and Myog, suggesting their differentiation into the muscular lineage. After single point injection, the differentiation of SKPs from green fluorescent protein positive donors into muscle precursors was also demonstrated in vivo. Additionally, donor SKPs migrated to the niche of muscle progenitors, participated in the regeneration of recipient muscles, and expressed markers of muscle progenitors, including Pax7, M-cadherin, and MyoD. After recovery of donor cells from recipient muscles at 3 weeks postinjection, some of the injected SKPs were converted to myogenic precursors, based on the expression of specific markers (Pax7 and MyoD). Some of these donor cells also expressed muscle makers (desmin, TnT, and Myog). At 20 weeks postinjection, the injected SKPs were localized to recipient muscles without decreases in their population size. In summary, these findings indicated that SKPs could develop into muscle progenitors and differentiated muscle cells in vitro and in vivo, thus providing valuable autologous cells for the treatment of muscle diseases.
Introduction
S
Transplantation of muscle stem cells has been regarded as an efficient method for the treatment of muscular diseases in mouse models [11,13]. However, their extremely low numbers and difficulties for enrichment limit their utility for cell therapy. Besides myogenic cells derived from muscles, several alternative sources for muscle repair have been found including mesangioblasts [14], bone marrow-derived myogenic progenitors [15,16], adult muscle stem cells [17], and ES-derived muscle precursors [18,19]. However, these alternative sources from somatic stem cells, similar to bone marrow-derived myogenic progenitors, are rare and exhibit a limited ability for expansion in vitro. Although ES-derived muscle cells have been shown to have therapeutic potential in the treatment of muscular dystrophy [18,19], its utility is still restricted. Previous studies suggested that it was difficult to differentiate ES cells into muscle precursors in vitro, because of inefficient paraxial mesoderm formation during embryoid body (EB) differentiation [18,19].
Skin-derived precursors (SKPs), derived from skin [20,21], can provide abundant autologous cells for cell therapy. These cells are easily acquired and expanded in vitro. Previous studies suggested that SKPs could be differentiated into many cell types, including neural cells, muscle cells, adipocytes [20,21], and insulin-producing cells [22] in vitro, suggesting their great potential for therapeutic application. Here, we differentiated SKPs to muscle precursors in vitro through a rapid protocol and demonstrated that SKPs could be engrafted to the niche of muscle progenitors through single point injection. Moreover, the injected donor cells could be retained in the recipient muscle up to 20 weeks postinjection without decreases in their population size. Through immunostaining in tissue sections and recovery of injected cells from recipient muscles, we demonstrated that these cells were converted to muscle precursors and expressed many myogenic markers. As abundant SKPs are easily acquired autologous progenitors, the transplantation of SKPs is a promising therapy for degenerative muscle diseases.
Materials and Methods
Mice
Neonatal and 4-week-old male Balb/C nude mice were purchased from Institute of Genetics and Developmental Biology, Chinese Academy of Sciences. Neonatal GFP transgenic mice were obtained from our laboratory. The animals were all maintained in accordance with institutional rules following the Guidelines for the Care and Use of Animals in Research. Mice were allowed free access to water and foods and raised in a constant photoperiod (14L: 10D).
Cell culture
Skin-derived precursors (SKPs) were isolated using a previously described method [20,21], with modifications. In brief, skin from abdomen and back was dissected free of other tissues, cut into 1–2 mm3 pieces, washed three times in phosphate-buffered saline (PBS), and then digested with 0.125% trypsin for 45 min at 37°C. Tissue pieces were then washed twice with PBS. Skin pieces were manually dissociated by pipetting into a 1-mL pipette, a process that was repeated until the tissue could not be broken down further. The cell suspension was then poured through a 40-µm cell strainer. Dissociated cells were centrifuged at 1,000 rpm for 8 min and were washed twice with medium (DMEM-F12, 1:1). Then cells were resuspended in 10 mL medium containing B-27 (Gibco-BRL, Carlsbad, CA), 20 ng/mL EGF, and 40 ng/mL bFGF (both from Sigma). Cells were cultured in 60-mm2 tissue culture flasks (BD Falcon, Mississauga, Ontario, Canada) in a 37°C, 5% CO2 tissue culture incubator. To passage floating clusters of cells, medium containing spheres was centrifuged and the pellet was manually dissociated with a 1-mL pipette. The cells were re-seeded in fresh medium as described earlier. Cells were passaged every 6–8 days.
For myospheres preparation [23], hind-limb muscles from neonatal mice were isolated, minced by scissors, and digested with 0.125% trypsin for 1 h at 37°C. A single cell suspension was obtained after filtering the samples through a 40-µm cell strainer. Isolated cells were pre-plated overnight on culture dishes in DMEM supplemented with 20% fetal bovine serum (FBS) 2 mM glutamine, 1% penicillin–streptomycin. Myospheres were formed after 3–4 days.
The murine ES cell lines R1 was cultured in DMEM with 20% FBS and 1,000 U/mL leukemia inhibitory factor (LIF, BD). To induce embryonic body (EB) formation [22,23], ES cells were dissociated with trypsin and suspended in Petri dishes (Alpha Medical Instrument Corp) with 10% FBS/DMEM without LIF. For 3–5 days, EBs were collected for further analysis.
Neuronal and glial differentiation of sphere colonies in vitro
For differentiation [20,21], spherical colonies were trypsinized and dissociated into single cells, then cultured in DMEM supplemented with 2% FBS (Gibco) on poly-
In vitro induction of mouse SKPs to muscle progenitors
SKPs were seeded into 35-mm dishes at a density of 10,000 per mL in growth medium supplemented with 10% FBS (Invitrogen, Carlsbad, CA). Beginning 24 h after seeding, cultures were treated for 24 h with 10 µM 5-azacytidine (Sigma). Cultures were then washed twice with PBS, and the culture medium was changed to differentiation medium containing 2% FBS, 50 µM hydrocortisone (Sigma), and 4 ng/mL IGF-1 (Peprotech, Rocky Hill, NJ), 4 ng/mL bFGF (Sigma). The medium was changed twice a week for 12 days until the experiment was terminated. To perform immunostaining for Pax7 (R&D), desmin (Abcom), TnT (Abcom), cells were rinsed twice with PBS, fixed with 4% paraformaldehyde for 15 min, permeated using 0.2% Triton X-100 for 10 min, blocked with 5% BSA for 1 h, and then incubated overnight with primary antibody at 4°C overnight. The following secondary antibodies were used according to the manufacturer’s recommendations. For some samples, pre-immune IgG of rabbit, goat, or mouse were used as negative controls.
In vivo induction of mouse SKPs to muscle progenitors
Four-week-old male Balb/C nude mice were used for the in vivo model of muscle regeneration. Fifty microliters of 10 µM CTX (cardiotoxin; Sigma) was injected into the TA muscle. One day later, 5 × 106 mSKPs from GFP transgenic mice suspended in 50 µL PBS were administered (single point injection) into the same TA muscle (tibialis anterior). The other TA muscle served as internal control receiving PBS. Three weeks or 20 weeks later, TA muscles were removed, embedded in ornithine carbamoyl transferase, frozen in liquid-nitrogen-cooled isopentane, and cut on a cryostat into 7 µm serial sections. Sections were fixed in 4% paraformaldehyde and stained with antibody of laminin (Abcom), Nuclei were stained with 0.01 mg/mL of Hoechst 33342 (Sigma) for 10 min. Samples were viewed under a laser-scanning confocal microscope (Leica).
For the recovery of GFP+ cells, hind-limb muscles from recipient mice transplanted with GFP+ SKPs were removed, minced by scissors and digested with collagenase type A (Sigma) for 1 h at 37°C. A single cell suspension was obtained after filtering the samples through 40 µm cell strainer. Adherent cells were kept in culture for immuofluorescence. After culturing in differentiation medium for 1 day, adherent cells were fixed in 4% paraformaldehyde and stained with antibodies for Pax7 (R&D), desmin (Abcam), MyoD (Abcam), myogenin (Abcam), or TnT (Abcam).
Real-time PCR
Total RNAs were isolated from cells using Trizol reagent (Invitogen) according to the manufacturer’s instructions. A total of 2 µg of RNA was reverse-transcribed by using avian myeloblastosis virus reverse transcriptase (Promega) and 2 µg of random hexamer primers (Takara). All real-time PCRs were conducted using ABI-Prism 7300 (Applied Biosystems, Foster City, CA), by Shanghai GenePharma Co., Ltd. The sequences of primers used are listed in Table 1. Results were normalized to β-actin levels using the comparative Ct (2−⊗⊗Ct) method. The results are expressed as fold induction over controls.
P
Immunofluorescence staining
Sections were fixed in 4% paraformaldehyde solution before staining for immunfluorescence. After washing three times in PBS, the samples were permeabilized with 0.2% Triton X-100 (Sigma) for 20 min. At room temperature, sections were rinsed several times in PBS and then incubated in 5% BSA for 1 h at room temperature. After the BSA solution was aspirated, the samples were incubated with appropriate primary antibodies of GFP (Santa Cruz, CA), Pax7 (R&D), MyoD (Abcom), M-cadherin (Abcom) at a concentration 1 µg/mL at 4°C overnight. After rinsing in PBS three times, samples were incubated with FITC-labeled, Cy5-labeled, or TRITC-labeled secondary antibodies, respectively (Zhongshan Biotechnology Beijing, People’s Republic of China), at 37°C for 1 h. After rinsing in PBS, nuclei were stained with 0.01 mg/mL of Hoechst 33342 (Sigma) for 10 min. Samples were viewed under a laser-scanning confocal microscope (Leica). For some samples, pre-immune IgG of rabbit, goat, or mouse were used as negative controls.
Statistics
To estimate GFP+ satellite cells and GFP+ muscle fibers, six sections, each 10 µm thick and 200–300 µm apart, were prepared for each muscle specimen and the number of GFP+ cells and GFP+ muscle fibers in each section was counted. We also selected five random views in each section and counted the total number of satellite cells and myofibers for each section. The ratio between GFP+ satellite cells and the estimated total number of satellite cells was determined. The ratio between GFP+ muscle fibers and the estimated total number of muscle fibers was determined. One-way ANOVA was used for statistical evaluation of the data. Values were presented as mean ± SEM. P value <0.05 was considered statistically significant.
Results
Differentiation potentials of SKPs and ES cells into muscle progenitors
SKPs and ES cells were cultured in growth medium according to the protocol described in Materials and Methods. After growth factors withdrawal, SKPs were differentiated in 2% FBS [20,21]. ES cells were cultured in DMEM with 20% FBS and 1,000 U/mL leukemia inhibitory factor [24,25]. After differentiation for 3 to 5 days, SKPs-derived cells and EBs were collected for RNA isolation and real-time RT-PCR analysis. We examined the expression of pluripotent markers, Oct4 and Sox2, at different days after differentiation. The expression of Oct4 mRNA was rapidly decreased during the differentiation of SKPs and ES cells (Supplementary Fig. 1). (Supplementary materials are available online at http://www.liebertpub.com/) In contrast to EBs, expression of Sox2 was increased during differentiation of SKPs, consistent with the preference for SKPs to develop into neural lineage [20,21]. We also analyzed the expression of Hoxb1 [24] and Tbx6 [26,27] (paraxial mesoderm markers). Both of these genes were up-regulated during differentiation of SKPs and ES cells, suggesting their potential to differentiate into muscle precursors. However, the expression of Pax7 and Myf5 (muscle progenitor markers) was higher during the differentiation of SKPs as compared with that of EB cells (Supplementary Fig. 1), indicating a stronger potential for SKPs to differentiate toward muscle progenitors.
Comparison of morphology and gene expression patterns between myospheres and SKPs
Since SKPs could spontaneously differentiate into muscle cells in vitro [20 –22], we examined the similarities between SKPs and muscle progenitors in their morphology and gene expression patterns. Muscle myospheres were isolated according to the protocol described before [23]. After culturing for 1 week, muscle progenitors and SKPs formed colonies with a similar morphology (Fig. 1). We further analyzed the expression of marker genes, including Oct4, Stat3, nestin, and fibronectin in these spheres. As shown in Figure 1, these genes were highly expressed in both spheres. After growth factor withdrawal, these spheres started to differentiate in vitro. Subsequently, βIII-tubulin-, MAP2−, and NFM-expressing neural cells appeared, indicating the neural differentiation potential of myospheres and SKPs (Supplementary Fig. 2). These findings are consistent with previous reports demonstrating the competence of SKPs and muscle precursors to differentiate toward neural cells [20 –22,28].

Comparison of the colonies morphology and gene expression patterns between skin-derived precursor cells (SKPs) and myospheres. (
In vitro differentiation of SKPs to muscle progenitors
SKPs were differentiated in the medium containing 2% FBS, 50 µM hydrocortisone IGF-1, FGF, after treatment with 5-azacytidine (Fig. 2A). After 12 days of culture, SKPs-derived skeletal myocytes adopted a bipolar morphology as compared with the morphology of SKPs (Fig. 2a1, 2a2). Moreover, some SKPs-derived myotubes could be spontaneous twitch (Supplementary Video 1). At 12 days after differentiation, Pax7+ muscle progenitors, desmin+, and TnT+ muscle cells were detected (Fig. 2B). In contrast, these proteins were not detected in undifferentiated SKPs (Supplementary Fig. 3). We further analyzed the expression of Pax3, desmin, Mstn, and Myog mRNAs using real-time RT-PCR following the induction of differentiation. As shown in Figure 2C, the expression levels of these genes were elevated in a time-dependent manner as compared to non-differentiated SKPs. At 12 days after differentiation, their expression levels were comparable to those found in myospheres, indicating the conversion of SKPs to muscle progenitors and muscle cells. We also determined the percentage of cells showing positive staining for myogenic markers. As shown in Figure 2D, 41% ± 11% of cells were Pax7+, 59% ± 20% of cells were desmin+, and 26% ± 4% of them were TnT+.
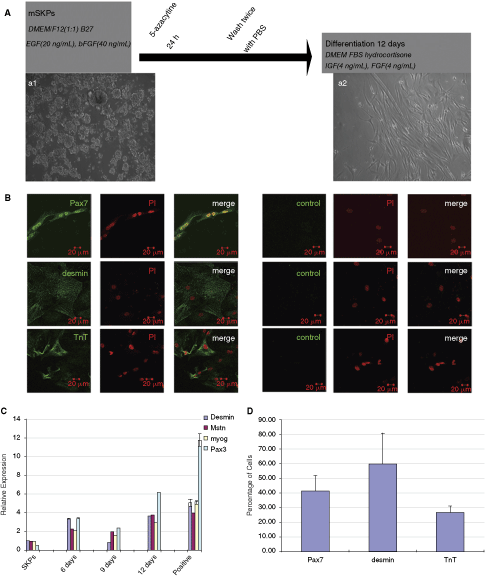
In vitro differentiation of skin-derived precursor cells (SKPs) into muscle progenitors. (
The donor SKPs were localized to the niche of muscle progenitors after single point injection into recipient mice
To investigate the differentiation potential of SKPs to muscle progenitors in vivo, we transplanted GFP+ SKPs into recipient mice, following single point injections to TA muscles as described in Materials and Methods. To localize injected cells, we isolated SKPs from GFP transgenic mice, and found the GFP proteins expressed in their nuclei (Supplementary Fig. 4A). Three weeks postinjection, the recipient mice were analyzed for the existence of GFP+ cells in their muscles. Previous studies suggested that adult muscles contained two major types of progenitors that were involved in myogenesis [2,29 –31]. Satellite cells are localized in the basement membrane juxtaposed to muscle fibers and interstitial cells are localized in the interstitial spaces of skeletal muscle. In response to injury or following disruption of the basal lamina, satellite cells can fuse to form multinucleated myotubes. We analyzed the existence of GFP+ cells in these two regions of recipient muscle. We found that donor GFP+ cells were enriched in both regions at 3 weeks postinjection (Fig. 3A–3D, Supplementary Figs. 4B and 5A). The number of SKPs that localized to the niche of satellite cells per muscle was 2.67% ± 1.53%. Moreover, GFP+ cells could form multinucleated myotubes (Fig. 3G). The number of fibers derived from SKPs was 4.33% ± 1.53%, which is lower than those derived from myospheres (14.33% ± 2.52%). These results suggested that the injected cells could migrate to the niches of muscle progenitors in vivo.
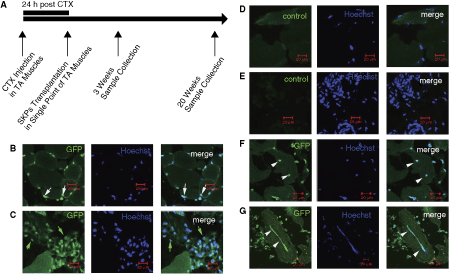
Engraftment of green fluorescent protein positive (GFP+) donor cells to the niche of myogenic progenitors in recipient muscles. (
In vivo differentiation of SKPs to muscle progenitors following single point injections into muscles of recipient mice
To further investigate the differentiation of SKPs in recipient mice, we focused on GFP+ cells localized to the niche of satellite cells. Immunostaining analysis indicated that injected GFP+ cells expressed multiple markers of satellite cells, including Pax7, M-cadherin, and MyoD. However, only a proportion of these donor GFP+ cells were localized in the niche of satellite cells expressing these markers (Fig. 4A, Supplementary Fig. 5B) with Pax7, M-cadherin, and MyoD expressed in 42% ± 5%, 39% ± 12%, and 57% ± 16% of GFP+ donor cells (Fig. 4B).
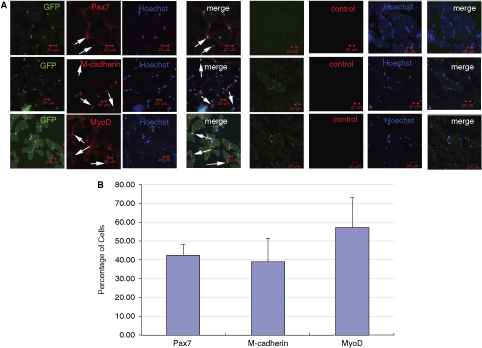
In vivo differentiation of skin-derived precursor cells (SKPs) to muscle progenitor through single point injection of SKPs to recipient mice. (
Three weeks postinjection, we recovered GFP+ donor cells from recipient muscles and analyzed the expression of muscle progenitor markers (Pax7 and MyoD) and muscle marker genes (desmin, Myog, and TnT) in the donor cells based on immunostaining. We found that all of these muscle progenitor markers and muscle genes were expressed in recovered GFP+ donor cells (Fig. 5). Because Pax7, desmin, and TnT were not expressed in SKPs (Supplementary Fig. 3), these results suggested that SKPs were converted to muscle progenitors and muscle cells in the recipient myogenic niche.
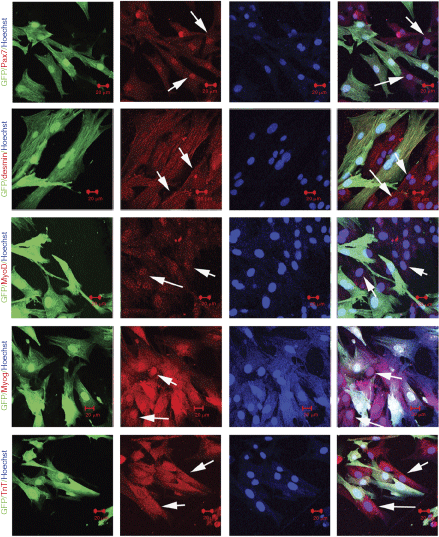
Expression of muscle progenitor makers and mature muscle markers in green fluorescent protein positive (GFP+) donor cells recovered from recipient mice at 3 weeks postinjection. Fluorescence of GFP and staining of muscle progenitor markers Pax7 and MyoD, and mature muscle genes (desmin, Myog, and TnT) in recovered donor cells at 3 weeks postinjection. GFP− cells from host mice also expressed these myogenic markers (arrows). Scale bars = 20 µm.
SKP-derived donor cells participate in the regeneration of recipient muscles and their long-time retention in recipient muscles at 20 weeks postinjection
We investigated whether the injected donor cells could participate in the regeneration of recipient muscle. Previous studies indicated that activated satellite cells could fuse to form a new fiber, and the nuclei derived from satellite cells were centrally located [31]. We found that there were GFP+ donor cells in the middle of the recipient muscle fibers, suggesting the involvement of donor cells in the regeneration of recipient muscle (Fig. 3B). More importantly, we found that there were still GFP+ donor cells present in recipient muscles at 20 weeks after injection (Fig. 6A) and most of them were localized in the niche of satellite cells (Fig. 6A). We further calculated the percentage of GFP+ donor cells in the niche of satellite cells (Fig. 6B). Compared with 3 weeks postinjection, the percentage of GFP+ donor cells at 20 weeks after injection did not show a decrease, suggesting the stable retention of these injected cells in recipient muscle.
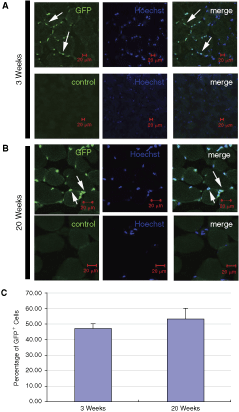
Prolonged and stable retentions of green fluorescent protein positive (GFP+) cells in the satellite cell niche of muscles in recipient mice. (
Discussion
Although ES cells are widely accepted as pluripotent cells with the potential to differentiate into different lineages after transplantation, the differentiation of ES cells toward muscle progenitors is still difficult [18,19,32]. It is possible that ES cell differentiation does not recapitulate the earlier differentiation of embryos and lacks the production of somites, where the signals for the derivation of muscle progenitors are originated [33]. In our study, we compared the spontaneous differentiation of ES and SKPs into muscle progenitors in vitro. Our results suggested that genes expressed in muscle progenitors, such as Pax7 and Myf5, were highly expressed during SKPs differentiation (Supplementary Fig. 1). These data suggested that SKPs were capable of differentiating into muscle progenitors. Moreover, we observed a relative high efficiency of SKPs differentiation into muscle progenitors and muscle cells in vitro and in vivo (Figs. 2 and 4). An explanation for the differentiation preference of SKPs to muscle progenitors was that the developmental origin of SKPs and muscle progenitors are similar. As SKPs are derived from dermal papilla [34 –37], whereas dermis originates from dermomyotome, and muscles originate from the border of dermomyotome [33].
The use of ES-derived cells for the treatment of degenerative muscle diseases is hampered by the complicated methods needed for the isolation and enrichment of EB-derived cells, the low efficiency of ES cell differentiation into muscle progenitors, and the formation of teratomas after transplantation [19]. SKPs could be freshly isolated and could provide autologous cells for transplantation [20,21]. Here we successfully differentiated SKPs into muscle progenitors through a simple protocol with a relatively high efficiency and these SKP-derived muscle progenitors were stably retained in recipient muscles without tumor formation at 20 weeks post-transplantation. These data indicated that SKPs-derived muscle cells could be a promising source for cell transplantation-based therapy.
After transplantation of GFP+ SKPs to recipient mice, the GFP+ cells could migrate to the niche of muscle progenitors, as earlier as 3 weeks postinjection (Fig. 3B and 3C). After recovery of GFP+ cells following dissociation of the recipient muscles, most of these GFP+ cells expressed muscle progenitor markers including Pax7 and MyoD. Some of these recollected cells also expressed mature muscle markers including desmin, Myog, and TnT. These findings demonstrated that SKPs could differentiate into muscle cells in vivo.
We did not detect specific localization of GFP+ cells in other tissues of recipient mice at 3 weeks postinjection (data not shown), suggesting that SKPs could specifically migrate to the niche of muscle progenitors, after damage was induced by CTX in the recipient muscle. In previous studies, hematopoietic stem cells were found to migrate to their niche in bone marrow, after transplantation into irradiated recipient mice [38 –40]. In this process, the homing efficiency was regarded as one major criterion to influence transplantation efficiency and therapy. Here, we found a very quick and efficient localization of donor GFP+ SKPs into the niche of muscle progenitors of recipient mice (Fig. 3). It is possible that damage induced by CTX in recipient muscle led to the release of chemokines that attracted the donor SKPs, due to their expression of unique plasma membrane proteins [41,42]. Because we did not observe any prominent localization of GFP+ cells in muscles of recipient mice without CTX-induced damaged (data not shown), the migration of SKPs to the damaged muscle is likely to be an active process.
In summary, we induced the differentiation of skin-derived progenitors from GFP+ transgenic and detected the long-term retention of injected GFP+ cells in the niche of muscle progenitors of recipient mice up to 20 weeks postinjection (Fig. 6). And the population size of these donor cells did not decrease with time. This finding indicated the efficient conversion of SKPs to muscle progenitor cells in vivo. Our studies suggested that SKPs could be to a promising source for cell transplantation-based treatment for muscle diseases.
Footnotes
Acknowledgments
We thank Professor Aaron Hsueh (Stanford University) for reading the manuscript. This work was supported by the National Basic Research Program of China (2007CB947401, 2006CB944006) and Chinese National Manned Space Program (Project 921-2).