
Abstract
The canals of Hering or biliary ductules have been described to connect the bile canaliculi with the interlobular bile ducts, and thus forming the distal part of the biliary tree. Studies in the last two decades suggested that the cells constructing these ductules could behave as hepatic progenitor cells. The canals of Hering are confined to the periportal space in the rat, while they have been reported to spread beyond the limiting plate in human liver. The distribution of the distal biliary ductules in normal human hepatic tissue has been investigated in our recent experiments. We could demonstrate the presence of interlobular connective tissue septa in a rudimentary form in healthy livers. The canals of Hering run in these septa in line with the terminal branches of the portal vein and hepatic arteries. This arrangement develops in the postnatal period but regresses after early childhood. The canals of Hering can be identified by the unique epithelial membrane antigen (EMA)−/CD56+/CD133+ immunophenotype. The canals of Hering leave the periportal space and spread into the liver parenchyma along rudimentary interlobular septa outlining the hepatic lobules. Our observations refine the original architectural description of the intraparenchymal portion of the canals of Hering in the human liver. The distinct immunophenotype supports their unique biological function.
Introduction
T
Most data refer to the canals of Hering as the site of the hepatic progenitor cell compartment [1,2,10]. This structure was described by Hering as “hepatic capillaries” [11], which maintained the link between bile ducts and the hepatocyte canalicular system. Later, it had been proposed to be the niche for hepatic progenitor cells [12,13]. The niche is a special microenvironment, which has a major impact on the maintenance and activation of the stem/progenitor cell compartment [14]. Therefore, the exact identification and characterization of this structure is necessary to understand its behavior under normal and pathological conditions.
The canals of Hering are usually shown as short, straight ductules at the border of the periportal connective tissue and liver parenchyma, but probably this conformation is oversimplified. We have recently characterized the hepatic progenitor cell niche in rat liver by laser scanning confocal microscopy [15]. Long, branching ductules have been observed, strictly inside the periportal connective tissue. They had contact with the bile canalicular system at the limiting plate. Their unique CK19+/CK7− immunophenotype has made their identification within the biliary tree easier. However, no CK7− biliary structures could be observed in human liver specimens. Furthermore, the canals of Hering have been described to spread into the hepatic lobule in the normal human liver [16]. These observations suggest that the organization of the canals of Hering in human liver is different from the traditional simple view as well as from the architecture we saw in rat liver.
In our present study, we set out to collect normal human liver tissue from individuals of various ages and analyzed the architecture and immunophenotype of the biliary ductules by confocal microscopy.
Materials and Methods
Normal human liver specimens were collected from cadavers of spontaneous premature birth neonates without developmental abnormalities and individuals who died suddenly in accidents without morphological signs and anamnestic data of any liver disease. (Age and gender of the patients: 3 males, 23rd week of pregnancy; 4 females, 23rd week of pregnancy; 1 male, 39th week of pregnancy; 2 males, 3 years; 1 female, 3 years; 1 female, 13 years; 1 male, 20 years; 1 male, 26 years old.) All autopsies were performed within 24 h following death. The liver samples were thoroughly examined on formalin-fixed, paraffin-embedded liver sections with H&E, diastase PAS, Prussian blue, orcein and Masson’s trichrome stainings; no fibrosis, ductular reaction, or other pathological alterations were observed. Snap frozen liver samples were stored at −80°C.
Frozen sections (10–20 µm) were fixed in methanol and were incubated at room temperature (1 h) with a mixture of primary antibodies (Supplementary Table 1;Supplementary materials are available online at http://www.liebertpub.com/) followed by the appropriate fluorescent secondary antibodies (JacksonImmunoresearch, West Grove, PA). All samples were analyzed by confocal laser scanning microscopy using Bio-Rad MRC-1024 system (Bio-Rad, Richmond CA).
The procedure has been approved by the ethical committee of Semmelweis University.
Quantitative analysis of immunohistochemical staining
The livers of two 3-year-old children and the 20-, 26-year-old adults were used for quantitation. Consecutive frozen sections were co-stained for CK7/EMA, CK7/CD56, and CK7/CD133. The number of CK7-stained structures was determined. The double-stained structures were counted and the results were given as percent of the CK7+ structures.
Results
Distribution of hepatic ductules in the liver parenchyma
When sections from the livers of 3-year-old healthy children were stained for panCK, CK7, and laminin, hepatic ductules surrounded by basement membrane could be observed in the parenchyma far from the portal spaces (Fig. 1A and 1B). Low power examination revealed that these ductules were not randomly arranged. They outlined dimly polygonal structures with terminal veins in the centers and portal triads at the corners, that is, the classical hepatic lobules. When micrographs of 40 serial sections stained for cytokeratin-7 were digitally aligned and merged (Fig. 1C), this kind of perilobular arrangement of the ductules was even more obvious. Two other characteristics could be observed on this composite image: (i) no CK7+ biliary ductules were present inside the hepatic lobules; (ii) the ductules at the interlobular border spread until the half of the porto-portal distances, which resulted in watershed-like empty gaps in the middle of these stretches.
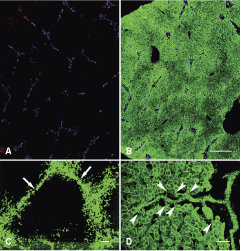
Confocal images of normal human liver from a 3-year-old child. (
High power examination of individual biliary ductules showed that these narrow tubules did really extend beyond the limiting plate. The ductules spread in virtual “empty” spaces among hepatocytes on cytokeratin antibody stained sections (Fig. 1D), where only the epithelial elements of the hepatic tissue were decorated. When the ductules terminated on hepatocytes, they were surrounded by typical U-shaped basement membrane (Fig. 2B inset), and the hepatocytes did not participate in the composition of the ductules beyond these connections. The “empty” space around the ductules was filled by collagenous matrix (Fig. 2A) and contained CD31+ small blood vessels (Fig. 2B), some of which were also labeled by the arterial marker NG-2 [17] (Fig. 2C). Taken together, confocal analysis of normal human liver revealed the deposition of small amounts of extracellular matrix between hepatic lobules, with expanding biliary ductules and blood vessels. This kind of arrangement of the blood vessels was described earlier as the vascular septum [18,19]. The small amount of matrix could not be visualized on traditional histological sections by special stainings on any of our liver samples.
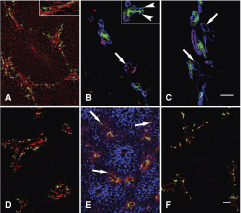
Confocal images of normal human livers from a (
Alterations of the hepatic ductules with age
The maturation of the biliary system continues in postnatal life in humans [20,21], and major changes are also observed in rats [15,22]. Therefore, we decided to examine the distribution of the canals of Hering in healthy livers of individuals of various ages.
The portal areas in the liver of immature neonates born on the 23rd week of pregnancy were “closed.” There were numerous bile ducts in the periportal connective tissue, especially at the periphery, as the remnants of the ductal plate. However, no signs of vascular septa were seen; no matrix deposition, blood vessels, or biliary ductules could be observed outside the limiting plate in any of the examined specimens (Fig. 2D).
Early signs of vascular septum formation could already be recognized in a liver sample derived from a fetus of the 39th week of pregnancy. It was mostly outlined by Thy-1-positive myofibroblasts, but a few CK7+ ductules were also present in these septum fundaments outside the portal fields (Fig. 2E).
The vascular septa and all of its components were mostly developed in the livers of young children of the age of 3 years (Fig. 1A and 1B). The only available liver specimen from a 13-year-old girl contained relatively regressed vascular septa, and although all the elements described earlier were present in the livers of young adults (20 and 26 years), they were more scarce (Fig. 2F).
Immunophenotypic characterization of bile ductules
There are several proposed markers for hepatic progenitor cells in human liver (1,2), but most of them did not distinguish hepatic ductules of the vascular septa from larger interlobular bile ducts in our hands. Some of the markers (AFP, chromogranin, synaptophysin, DMBT, DLK, CEA, CK20, CK14) did not label any biliary structures, while others (EpCAM, E-cadherin, CK7, CK19) stained the complete biliary tree (data not shown). Only three markers reacted differentially with bile ducts and ductules. Epithelial membrane antigen (EMA) resulted in a very sharp characteristic linear apical staining in the interlobular bile ducts. Conversely, it was absent in the ductules even on cross sections (Fig. 3A and 3B). The staining pattern of CD133 (Fig. 3C) and CD56 (Fig. 3D, 3E, and 3F) was opposite. In specimens up to the age of 3 years, there was a consistent apical CD133 staining in all segments of the biliary tree. However, in samples from older individuals the staining was strictly confined to the small ductules of the vascular septa. The distribution of CD56 in all specimens was similar to this latter case: it labeled exclusively the small ductules.
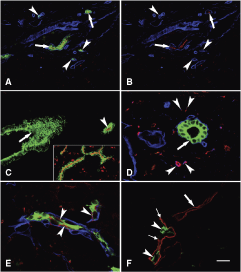
Immunophenotypical characterization of the hepatic ductules in (
Quantitative evaluation of the immunohistochemical reactions (Table 1) showed that only a small portion of the biliary structures were stained for EMA, and this staining was restricted to the periportal area. No such preferential distribution was noticed with the two other markers. Almost all ducts/ductules were decorated by CD133 in the livers of the children, while in adulthood the ratio of CD56+ and CD133+ ductules were similar. The CK7 antibody reacted sometimes with very small bile ductules occasionally appearing as single cells especially along the vascular septa. Since the CD56 and CD133 reaction was not as strong and diffuse as the CK7, the number of the CD56 and CD133 ductules was probably underestimated.
T
Abbreviation: EMA, epithelial membrane antigen.
( ) Total number of counted bile ducts.
Discussion
We have analyzed the architecture of biliary ductules in normal human livers and observed them to circumscribe the classical hepatic lobules by participating in the formation of the so-called vascular septa (Fig. 4). This arrangement develops in postnatal life and can be most obviously seen in early childhood. The hepatic ductules are characterized by a unique EMA−/CD56+/CD133+ immunophenotype.
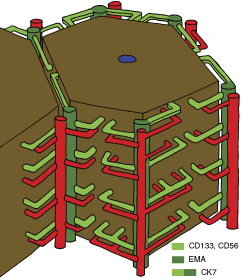
Schematic representation of hepatic lobule (brown), with bile ducts/ductules (green) and accompanying arterioles (red). Note that these structures extend only halfway into the porto-portal distances; however, they cover the whole “lateral” surface of the lobule. For simplicity, the portal vein branches are not shown.
There are several candidates for the liver stem/progenitor cell niche. Kuwahara et al. [10] proposed four structures to harbor such cells: the canals of Hering, intralobular bile ducts, periductal “null” mononuclear cells, and peribiliary hepatocytes. After all, stemness has been proposed to be not an entity but function [23] and—depending on the situation—different cell populations can behave as hepatic progenitor cells. Insofar, most evidence shows that the canals of Hering have the highest potential to behave as hepatic progenitor cells [1,2,6,13]. Therefore, the accurate architecture of these structures is a key issue to understand their behavior under normal and pathological conditions.
The canals of Hering were originally described [11] as short straight ducts at the limiting plate, which connect the bile canaliculi to the interlobular bile ducts. However, Theise et al. [16] demonstrated the extension of hepatic ductules through the limiting plate into the hepatic lobule. Our results confirm the presence of these structures deep in the hepatic parenchyma; moreover, a clear orientation of the ductules could be observed on our confocal images. The ductules spread into the parenchyma along the porto-portal axis. The hexagonal structures outlined by hepatic ductules correspond to the classical hepatic lobules. High power examination of individual ductules revealed their close correlation with bile canaliculi enabling their drainage.
In addition to former observations [19,24] that venular branches are present in vascular septa, we noticed NG-2-stained arterioles running in line with the ductules. There have been speculations about bile ductule escorting hepatic arterioles [18,19,25,26,27] but no convincing evidence has been published so far. Gouw et al. [19] and van der Heuvel et al. [28] emphasize the importance of the microvascular compartment for the efficient regeneration of ductules. Since—contrary to several other species—no arterio-portal anastomoses exist in the human liver, the presence of arterial blood would be advantageous for the regenerative function of the ductules. The proximity of the blood vessels to the bile ductules corresponds very well to the close correlation between the development of biliary and vascular structures [29]. This architecture of the intraparenchymal ductules and the escorting vessels is in full agreement with the proposed model of Matsumoto [18] based on 3D reconstruction of the human liver from thousands of serial sections.
According to the original description [18], the “vascular septum” is not a fibrous septum but a vascular surface from which sinusoids originate. However, we observed a collagenous matrix in this location. Since all the studied liver samples were normal and no fibrosis could be seen by traditional connective tissue staining, we suggest that a minimal amount of matrix material in the vascular septum, which can be visualized only by careful immunohistochemical analysis, is a component of the normal human hepatic tissue. Hepatic lobules are separated by well-defined connective tissue septa in several species [30], and the vascular septum of the human liver with its matrix components can be regarded as a rudimentary interlobular septum.
We have observed a peculiar age dependence of the vascular septa. Obviously, a more detailed age-related analysis of these structures is required. The interlobular bile ducts develop from the ductal plate [21], but ductal plates disappear shortly after birth and new bile ducts/ductules arise from pre-existent ducts by branching and elongation [31]. This “cholangiogenesis” could follow the primitive septa we observed in the liver of the 39-week-old fetus. The postnatal maturation of the biliary tree is well documented in humans [21] and rats [22] as well. Furthermore, the interlobular septum also develops postnatally in pigs [30].
The progressing scarcity of the ductular system with age should also be analyzed in more detail. We do not know if this process is absolute due to the apoptosis of biliary cells or just relative. The size of the hepatic lobules increases during ontogeny [32], and if the growth of the ductules is arrested earlier, it may be responsible for their relative regression.
We were curious if the biliary ductules could be characterized by a special immunophenotype. Three different antibodies were able to distinguish reliably the canals of Hering from larger bile ducts: the canals of Hering were EMA−/CD56+/CD133+; whereas, interlobular bile ducts were EMA+/CD56−/CD133−. All of these markers have already been mentioned in connection with the hepatic progenitor cell compartment. Atypical ductular reactions have been reported EMA−/CD56+, while the typical ones, which are similar to the interlobular bile ducts, are EMA+/CD56− [33]. CD56 has been demonstrated in proliferating ductules, while it could not be observed in normal canals of Hering [34,35], but recent studies found CD56 mRNA and protein in ductules of normal human livers [36,37]. CD133 has originally been described as a hematopoietic stem cell marker, and its mRNA has been detected in the liver by Northern hybridization, but no immunostaining was identified in paraffin sections by Miraglia et al. [38]. However, the protein could be detected by immunohistochemistry in the canals of Hering of normal human liver [39] and in regenerating ductules related to fulminant liver failure [40]. In our present experiments, the distribution of this marker showed an age-dependent change. This is similar to our results on rat liver, where the immunophenotype of the canals of Hering developed postnatally [15]. Interestingly, CD133+ cells isolated from HCC proved to be highly tumorigenic and have been reported as tumor stem cells [41]. Increased expression of CD133 has also been reported in a subset of cholangiocellular carcinomas, which were claimed to have a progenitor cell origin [42].
The combined application of these three antibodies provides an efficient tool for the identification of the canals of Hering in the normal human liver. Furthermore, the distinct immunophenotype of the hepatic ductules supports their different biological potential.
In conclusion, we present a refinement for the widely cited architectural description [16,27,43] of the intraparenchymal biliary ductules in normal human liver. The canals of Hering with escorting vessels are situated in the vascular septum and are components of a rudimentary interlobular septum. They can be distinguished from larger bile ducts by a unique immunophenotype. Better comprehension of canals of Hering’s architecture in normal human liver may promote our understanding of their behavior in various pathological/biological reactions.
Footnotes
Acknowledgments
The authors appreciate the help of Dr. Nóra Szlávik, Dr. Éva Görbe, and Dr. Júlia Hajdú in collecting the samples. The article is supported by OTKA K 67697.
Author Disclosure Statement
We declare that we have no duality of interest.