
Abstract
Lectins form an important constituent of our daily diet, and thus, it is essential that their effect(s) on various tissues be examined systematically in order to assess whether they are beneficial or detrimental to human health. We examined the effect of oral administration of two dietary lectins that were isolated from banana (BL) and garlic (GL)—two quite commonly consumed food items—on the hematopoiesis of mice. Balb/c mice were fed weekly with lectins and their marrow mononuclear cells (MNCs) were subjected to various hematopoietic stem/progenitor (HSPC)-specific phenotypic and functional assays. It was observed that the lectin-fed mice harbored a considerably increased HSPC pool in their marrow. Marrow-derived MNCs isolated from these lectin-fed mice gave rise to large-sized colony-forming unit-fibroblast (CFU-F) colonies indicating that the lectins had a salutary effect on the stromal compartment. The molecular mechanisms involved in the process were examined by using a stromal cell line model, M210B4. The lectins pulled down pro-insulin and insulin receptors in an immunoprecipitation experiment and activated extracellular signal-regulated kinase (ERK) signaling in the treated cells, in a manner comparable to insulin, both in terms of kinetics as well as extent. M210B4 cells incubated with BL, GL, or insulin showed reduced levels of reactive oxygen species, suggesting that perhaps the lectins protected the stem cell pool of mice by activating ERK signaling and reducing the oxidative stress in the niche. Our data suggest that these lectins may serve as micronutrients for therapeutic purposes in hematological deficiencies.
Introduction
D
Lectins are ubiquitously occurring, carbohydrate-binding proteins that serve several biological functions in animals, ranging from the regulation of cell adhesion to glycoprotein synthesis, and participate in the immune function [6] by recognizing specific carbohydrates on pathogens (the lectin complement activation pathway) or on host cells (mannose-binding lectin) [7]. The function of lectins in plants is still uncertain, but studies suggest a role in plant germination, seed survival, and defense against pathogens [8]. Purified lectins are important in a clinical setting because they are used for blood typing. The crystal structures of legume lectins have led to a detailed insight of the atomic interactions between carbohydrates and proteins [9].
The resistance of the lectins toward the proteolytic activities of digestive enzymes and their ability to enter systemic circulation and tissues, taken together with their ability to bind to cell surface molecules containing specific carbohydrate moieties, indicate that they may affect the functions of various tissues [5]. Dietary lectins like kidney bean lectin (Phaseolus vulgaris) when fed to rats resulted in abnormal microvilli development [4]. Similarly, higher content of wheat germ agglutinin though useful against pests was found to affect the growth of the rats with atrophy of the tissues [10]. Accordingly, it is of considerable importance to establish whether a given lectin has deleterious or beneficial effects for mammals [11].
We used lectins isolated from banana (Musa paradisiaca; BL) and garlic (Allium sativum; GL)—food items that are consumed in plenty and, quite commonly, in an unprocessed form—in the experiments to examine the effect of their oral administration on the hematopoiesis of mice. We selected these lectins because of several reasons: both lectins have well-defined mannose-binding specificities [12 –14]; mannosyl residues play an important role in stem cell functions like homing and engraftment [15]; mannose-specific lectins have been shown to preserve hematopoietic cells in vitro [16 –18]; both of them do not agglutinate RBCs [14,19], thus were not likely to induce tissue toxicity. Mice were weekly fed with small amounts of either BL or GL and the effect on their hematopoiesis, especially on the hematopoietic stem/progenitor compartment (HSPC), was assessed using various in vitro and in vivo HSPC-specific parameters. We show here that the bone marrow population of lectin-fed mice contains a substantially larger pool of primitive HSPCs, as was evident by the output of various phenotypic and functional assays.
Our results suggest that the lectins protect the HSPC pool probably through activation of extracellular signal-regulated kinase (ERK) signaling and also through antioxidant mechanisms evoked in the stromal compartment. These lectins may serve as micronutrients for therapeutic purposes.
Materials and Methods
Lectins
Isolation, purification, binding specificities, and crystal structure of both GL and BL have been published before [14,20 –25]. The isolation and purification steps are as follows: The dry garlic (A. sativum) bulbs were homogenized in 20 mM phosphate buffer (pH 7.4) containing 150 mM NaCl (PBS). The extract was filtered and centrifuged at 10,000 rpm. The supernatant was subjected to (NH4)2SO4 cut (70%), and was centrifuged. The protein pellet was resuspended in PBS and was dialyzed extensively against PBS. The lectin was purified by affinity chromatography on mannose-sepharose column followed by gel filtration on Bio-Gel P-200 column. Purified garlic lectin has a molecular weight of 25 kDa and is a heterodimer of 12.5- and 11.5-kDa subunits.
Ripe mashed banana fruit was added to 150 mM acetic acid solution (30 bananas into 2 L of 150 mM acetic acid) and was incubated at 4°C for 12–18 h. The solution was stirred with an overhead stirrer rotating at 10 rpm. The extract thus obtained was filtered with muslin cloth wetted with deionized water, centrifuged at 7,000 rpm at 4°C for 20 min and the cell debris was discarded. The supernatant was subjected to (NH4)2SO4 cut (65%) while stirring at 4°C overnight. The precipitate was dissolved in minimum volume of ice cold PBS (pH 7.4) and was dialyzed exhaustively for 48 h against the same buffer at 4°C. The dialyzed solution was centrifuged and the clear supernatant was loaded onto a mannose-sepharose column pre-equilibrated with PBS at 4°C. The column was washed with PBS till the OD at 280 nm was <0.005. The lectin bound to mannose-sepharose was eluted with 0.3 M α-methyl-
The purity of both the lectins was checked by SDS-PAGE [20,23]. The lectin preparations were exhaustively dialyzed against PBS to remove bound sugar and were sterilized by filtering through 0.2 µm filters prior to performing the experiments. The protein content of the lectin preparations was estimated using a micro BCA kit (Pierce, Rockford, IL, USA).
Cell lines
M210B4 cell line was purchased from American Tissue Culture Collection (ATCC, Manassas, VA, USA) and was maintained as per their guidelines.
Feeding
All the protocols used in animal experimentation have been approved by the institutional animal ethics committee. Balb/c mice (6–8-weeks old; five mice/group) were fed weekly with 10 ng of either BL or GL in 25 µL of sterile phosphate-buffered saline (PBS) through a feeding needle. PBS-fed mice were used as controls. Ten nanograms of BL correspond to ∼1 mg of banana and 10 ng of GL corresponds to ∼0.33 mg of garlic bulbs per mouse per week. The mice were sacrificed at 12 weeks (early) and 28 weeks (late) time points and the mononuclear cells were isolated from the marrow flushed out from the femoral bones (BM MNCs) using Ficoll-Hypaque (Histopaque; Sigma, St. Louis, MO, USA) density gradient separation.
In vitro assays
Colony-forming unit (CFU) assay. 1–2 × 104 marrow mononuclear cells (MNCs) were seeded in 1% methyl cellulose containing murine recombinant growth factors, namely, interleukin-3 (IL3)—10 ng/mL, stem cell factor (SCF)—10 ng/mL, granulocyte-monocyte colony-stimulating factor (GM-CSF)—5 ng/mL, and erythropoietin (EPO)—2 U/mL (Peprotech, Rocky Hill, NJ, USA) [26,27]. Colonies belonging to blast-forming unit erythroid (BFU-E), granulocyte-monocyte (GM), and granulocyte-erythroid-monocyte-megakaryocyte (GEMM) were scored after 14 days. The experiments were repeated thrice (n = 3) and five mice were kept in each group. Six replicate plates for each set were scored for the analysis.
Long-term culture initiating (LTC-IC) assay. M210B4 cells were seeded on collagen-coated wells of a 24-well plate and were γ-irradiated (8,000 rad) [28]. One milliliter of Myelocult medium (Stem cell Tech, Vancouver, CA) containing freshly prepared hydrocortisone (10−6 M; Sigma) was added to each well. 1 × 105 MNCs were seeded per well of a 24-well plate and the cultures were maintained with a weekly demi defoliation. After 5 weeks, the cultures were harvested and colony-forming unit (CFU) assay was carried out as described earlier.
In some experiments, LTCs were established with BM cells from control (unmanipulated) mice with the addition of BL or GL (20 ng/mL). The lectins were added from the day of initiation of LTC and were replenished every week. The assay was carried out as described earlier.
Extended LTC-IC assay. This assay was carried out like the LTC-IC assays described earlier, except that in these assays, after every 4 weeks the feeder layers were changed [29]. The cultures were harvested after 12 weeks and the cells were subjected to CFU assay using thrombopoietin (TPO)—25 ng/mL, FMS-like tyrosine kinase 3 (FLT3)—25 ng/mL, SCF—25 ng/mL, IL3—10 ng/mL, GM-CSF—5 ng/mL, EPO—5 U/mL.
CFU-Fibroblast (CFU-F) assay. 3 × 106 BM MNCs from the lectin-fed mice (28 weeks) were suspended in Iscove’s modification of Dulbecco’s medium (IMDM) with 20% FBS (Stem Cell Tech) and were incubated in 60 mm dishes for 12 days without disturbing [30]. Nonadherent population was removed by washing and the adherent colonies were fixed with 1% buffered paraformaldehyde. The plates were stained with 0.1% crystal violet and the number of colonies was counted. The plates were imaged and size of the colonies was measured by Image-J software (NIH, USA). The data are represented as mean ± SD.
Similar experiments were carried out on marrow cells from unmanipulated mice. The BM MNCs were treated in vitro with BL, GL (both at 20 ng/mL), or insulin (2 µg/mL) in the presence or absence of mitogen-activated protein kinase kinase (MEK) inhibitor, U0126 (10 µM; Calbiochem, Darmstadt, Germany) and the plates were kept undisturbed for 12 days.
In vivo assays
CFU-S assay. BM MNCs of lectin-fed or PBS-fed mice were intravenously infused (105 MNCs/mouse) into lethally irradiated syngeneic mice (8- to 10-weeks old, six mice/group) [31]. The irradiation dose used was 9.5 Gy, given in two split doses 4 h apart (Co60 gamma chamber 5000; BRIT, Navi Mumbai, India). The mice were sacrificed on 12th day. The spleens were weighed and imaged. The colony size was measured by analyzing the images of the colonies using Image-J software (NIH, USA). The macroscopic colonies formed on the spleen (CFU-S) were counted under a dissection microscope after fixation in Tellyesniczky’s fixative (70% ethanol:formalin:glacial acetic acid in 20:2:1 parts). The BM MNCs (CFU-S marrow; Supplementary Fig. 1; Supplementary materials are available online at http://www.liebertpub.com/) were infused in secondary recipients for pre-CFU-S assay.
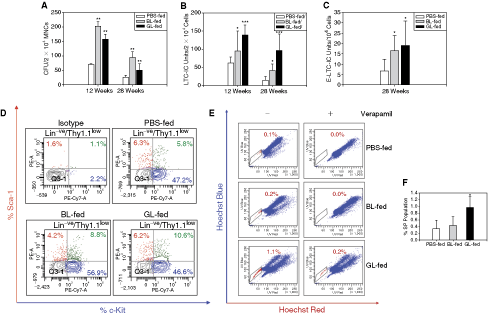
Intake of mannose-specific lectins, BL and GL, stimulates hematopoiesis. Marrow cells of PBS- or lectin-fed mice (five mice/set) were analyzed for CFU, LTC-IC, and E-LTC-IC contents. A 2- to 3-fold increase in the CFU content (
Pre-CFU-S assay. CFU-S marrow cells were infused into lethally irradiated (9.5 Gy) secondary recipients (105 MNCs/mouse, four mice/group). The mice were sacrificed on 12th day and the colonies formed on the spleens (pre-CFU-S) were counted. The BM MNCs (pre-CFU-S-marrow) were subjected to phenotypic characterization to detect stem cell population (flow chart—Supplementary Fig. 1).
Phenotypic analysis
BM MNCs of 2 × 106 were suspended in IMDM containing 20% FBS. Fc receptor was blocked by incubating the cells with CD16/CD32 (BD-Pharmingen, San Diego, California, USA) antibody at 4°C for 10 min. The cells were washed and were suspended in PBS containing 0.1% BSA and 0.1% sodium azide. They were further stained with c-Kit (CD117, clone-2B8)-PE-Cy7, Thy1.1 (CD90.1, clone-OX-7)-FITC, lineage marker cocktail (CD3e, CD11b, CD45R/B220, Ly-76, Ly-6G, and Ly-6C)-APC, Sca-1(Ly-6A/E, clone-D7)-PE (BD) at 4°C for 45 min with frequent mixing. The cells were washed with PBS and were fixed in 1% buffered paraformaldehyde. Appropriate isotype controls were used. Fifty thousand events in the lineage negative gate were acquired for each sample (FACS Canto II; BD Bioscience, San Diego, USA). The flow data were analyzed using FACS Diva™ (BD) software. Thy-1.1low population was gated in the lineage negative cells and the Lin− Thy-1.1low population was analyzed for c-Kit and Sca-1 double positive population (KTLS).
Side population (SP) analysis
SP analysis was carried out as described by Eaker and colleagues [32]. In brief, 106 MNCs of lectin-fed or PBS-fed mice were stained with Hoechst 33342 (Sigma), with or without 50 µM verapamil (Sigma), for 90 min at 37°C and were analyzed on a flow cytometer using UV laser (FACS Aria, BD; excitation λ: 350 nm, emission λ: 450 nm—blue and 650 nm—red).
Reactive oxygen species (ROS) detection
Cells, either M210B4 or BM MNCs, were incubated with lectins for 72 h in RPMI 1640 medium with 10% FBS and stem span serum-free medium (Stem cell Technology, Vancouver, CA), respectively [33]. The ROS levels were determined by staining them with 5 µM of 2′,7′-dichlorodihydrofluorescein diacetate (DCHFDA; Molecular Probes, Carlsbad, CA, USA) at 37°C for 30 min and the fluorescence levels were analyzed on a flow cytometer (Canto II, BD). Propidium iodide (PI) was added to exclude dead cells. The MNCs were further incubated with antibodies to Sca-1, c-Kit, and Lineage markers (Lineage cocktail; BD) to determine the ROS levels in the stem cell compartment. The flow data were analyzed using FACS Diva™ (BD) software. The overlays of the flow data were prepared using Cell Quest Pro software (BD).
Western blot and immunoprecipitation
M210B4 cells were treated with BL, GL (20 ng/mL), or insulin (2 µg/mL), with or without the presence of U0126 (10 µM) or α-methyl-
Statistical analyses
The data were analyzed by one-way repeated measure analysis of variance (one-way RM ANOVA) using Sigma Stat software (Jandel Scientific Corporation, San Raphael, CA). The values were represented as mean ± SD. P value ≤ 0.05 was considered as statistically significant. The symbols used were: *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001.
Results
Lectin-fed mice have higher progenitor cell content in their marrow
The mice were sacrificed after 12 and 28 weeks of feeding and the BM MNCs were subjected to CFU assay [26,27]. As seen in Figure 1A, the BM MNCs from BL- as well as GL-fed mice formed 2- to 3-fold higher number of colonies (P < 0.01) than those from the control mice (PBS-fed) at both the time points examined, indicating that the oral administration of both lectins stimulated the hematopoiesis of mice. The enhancement in colony formation was seen in all types of colonies, including the GEMM type (Supplementary Fig. 2A, indicating that the effect was not restricted towards a particular lineage. Since there was no significant difference between the yields of total bone marrow cells (data not shown) from the mice of various groups, the difference in the colony formation truly reflects the increase in the progenitors.
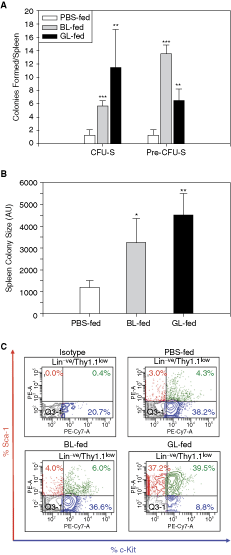
HSPCs of lectin-fed mice have better in vivo attributes. (
A direct addition of lectins in the assay plates, however, did not stimulate colony formation (data not shown), indicating that their effect was not mediated through secretion of colony-stimulating growth factors from the accessory cells like monocyte-macrophages, but probably involved other mechanism(s). Addition of lectins in the long-term cultures established using bone marrow cells from unmanipulated mice, however, significantly improved the LTC-IC unit output (Supplementary Fig. 2B), indicating that the effect was perhaps mediated through the stromal compartment.
Since several lectins have been suspected to exert anti-nutritional effects [5], we monitored the weight of the experimental animals. We did not observe any significant difference between the weights of control and lectin-fed mice (Supplementary Fig. 2C) indicating that the lectins were not causing nutritional deficiency.
Salutary effect of lectins on the primitive HSPCs
We carried out LTC-IC and E-LTC-IC assays to examine the effect of lectins on the primitive HSPCs. As seen in Figure 1B, the marrow of the lectin-fed mice contained a significantly higher number of LTC-IC units as compared to the marrow from the control animals at both the time points examined. E-LTC-IC assay is known to estimate the SRC-equivalent primitive stem cells [29]. The lectin-fed mice (28 weeks) showed the presence of 2- to 3-fold higher E-LTC-IC units as compared to the controls (Fig. 1C).
Presence of higher number of primitive stem cells in the marrow of the lectin-fed mice
Primitive stem cells are known to be c-Kit+ Thy1.1low Sca-1+Lin− (KTLS) [35,36]. As is clearly seen in Figure 1D, marrow cells of the lectin-fed mice (28 weeks) showed 1.5- to 1.8-fold higher (BL and GL sets, respectively) percentage of KTLS cells as compared to the controls (N = 2).
SP population is known to harbor the long-term populating stem cells that are critically important for giving a sustained multilineage hematopoiesis [37]. As seen in Figure 1E and 1F, the lectin-fed (28 weeks)—especially the GL-fed—mice had significantly higher percentage of SP population as compared to the PBS-fed mice (N = 3; *P < 0.05).
Thus, the results clearly show that the lectin-fed mice had a higher population of primitive stem cells in their marrow as compared to the controls.
Better in vivo functional attributes of HSPCs in lectin-fed mice
We carried out in vivo assays, namely CFU-S and pre-CFU-S, using BM MNCs of lectin- or PBS-fed mice (12 weeks). As seen in Figure 2A, the marrow cells isolated from the lectin-fed mice had a significantly higher number of CFU-S as well as pre-CFU-S units as compared to those from the control mice, underscoring their better functional attributes.
There was no significant difference in the weight of the spleens in BL-fed group when compared with the control, but the weight of spleens in GL-fed group was significantly high (P < 0.01; data not shown). The size of the colonies was significantly larger in the lectin-fed set as compared to the PBS-fed set (Fig. 2B; P < 0.05 for BL-fed and P < 0.01 for GL-fed).
We isolated the marrow cells of these secondary recipients (pre-CFU-S marrow) and analyzed them for the presence of KTLS population (flow chart—Supplementary Fig. 1). As seen in Figure 2C, the marrow from the recipients of CFU-S marrow of lectin-fed animals had a much higher percentage of KTLS cells as compared to the marrow of those receiving CFU-S marrow of control mice (1.4-fold higher in BL and 9-fold higher in GL). The results indicated that the lectin-fed mice harbored higher number of HSPCs in their marrow. A competitive repopulation assay needs to be carried out to enumerate the stem cells having a long-term engraftment capacity [38] in the marrow of the lectin-fed mice and to detect the subtle differences in them, if any.
Lectins stimulate stromal progenitors
Stromal cells create a microenvironment that fosters the growth of primitive stem cells. Therefore, the increased pool of HSPCs in the lectin-fed mice prompted us to examine whether the lectins had any effect on the stromal compartment. In order to examine this issue, we carried out CFU-F assay on the BM MNCs of the lectin-fed mice [30]. As seen in Figure 3A-a, the lectins did not have a significant effect on the number of CFU-F formed (P = 0.305), but the size of the colonies formed from the MNCs of GL-fed mice was significantly larger (Fig. 3A-b and c; ***P < 0.001) than the controls. The colonies formed from the MNCs of BL-fed mice were also larger than controls, but the data did not reach statistical significance due to a variation in the colony size. The results indicated that the lectins stimulated the growth of stromal progenitors. The effect of GL was more striking.
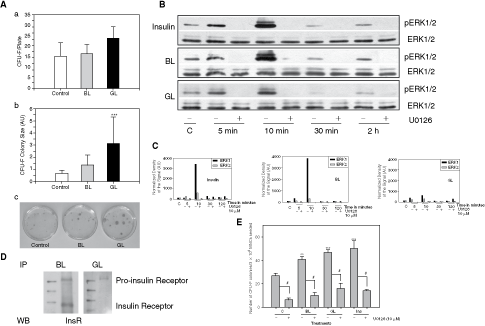
Lectins stimulate stromal progenitors via ERK signaling. (
Lectins initiate ERK signaling in the stromal cells
Stimulation of CFU-F growth by the lectins indicated that they may be inducing active signaling mechanisms in the stromal cells. In order to avoid the inherent heterogeneity of primary marrow-derived stromal cultures, and the consequent possible difficulty that may arise in the interpretation of the results, we used a well-characterized stromal cell line, M210B4, for the purpose. This cell line has been extensively used as stromal cells for experimentations and has been shown to support both murine as well as human HSCs [31,39
–42]. The serum-starved M210B4 cells were treated either with the lectins (BL or GL; 20 ng/mL) or with insulin (2 µg/mL, used as a positive control—known to activate ERK in M210B4 from our earlier experiments) for various time points, in presence or absence of an MEK inhibitor, U0126. The lysates were subjected to western blot analysis using antibodies that recognize the phosphorylated or the native forms of ERK. Both lectins induced a rapid phosphorylation of ERK in these cells and this activation was sensitive to the presence of U0126 (Fig. 3B) and α-methyl-
Lectins interact with insulin and pro-insulin receptors
The striking similarity in the kinetics of the MEK-dependent ERK activation by the lectins with that by insulin strongly suggested a possibility that the lectins may be signaling through insulin receptor. We, therefore, carried out an immunoprecipitation experiment with M210B4 cell lysate using biotin-labeled BL and GL and probed the blot of the immunoprecipitate with an antibody specific to insulin receptor β. As seen in Figure 3D, both lectins pulled down the pro-insulin receptor and BL pulled down both pro-insulin as well as insulin receptor β. The data show that perhaps BL signals through both pro-insulin as well as insulin receptor β, but GL signals only through pro-insulin receptor. Thus, the milder ERK activation induced by GL, as compared to that induced by BL or by insulin, could be due to its action through pro-insulin receptor, rather than through insulin receptor. The lectins did not compete with each other for binding in a flow cytometric analysis, indicating that perhaps they recognize separate, but closely spaced, epitopes of pro-insulin receptor (Supplementary Fig. 4).
In order to examine whether insulin also showed a salutary effect on the CFU-F formation, we incubated marrow MNCs from unmanipulated mice in the presence of BL, GL, and insulin. As seen in Figure 3E, the number of CFU-F formed in the lectin- or insulin-treated dishes were significantly higher than the untreated MNCs (**P > 0.01, ***P < 0.001) and the effect was sensitive to the presence of U0126 (#P < 0.001). These results indicate that insulin receptor-mediated signaling indeed stimulates stromal cells.
Lectins reduce oxidative stress in the M210B4 stromal cells
Results of all the experiments carried out so far clearly indicated that the marrow of the lectin-fed mice contained higher HSPC pool. Oxidative stress plays an important role in the onset of aging [44]. We, therefore, hypothesized that the lectins perhaps protected the HSPC pool by preventing their aging through the reduction of ROS levels in them. We examined the antioxidant effect of lectins on MNCs. As seen in Figure 4A-b and 4B, both % DCHFDA+ve and the mean fluorescence intensity (MFI) in the KLS cells from the MNCs that were incubated with GL were about 20% lower than those from the control MNCs (N = 3; P < 0.05). The % of DCHFDA+ve cells in the BL-treated KLS population was higher and their MFI was only marginally lower than those of controls. The results indicate that only GL reduced the oxidative stress in the stem cells to some extent.
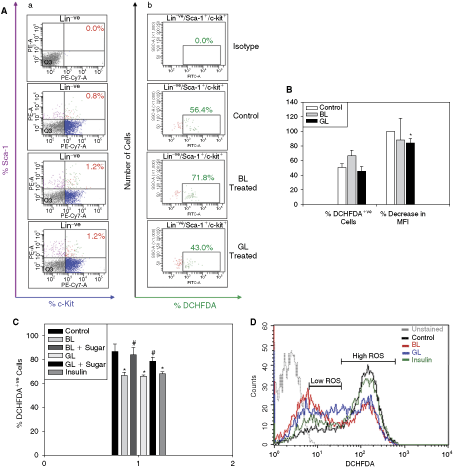
Lectins protect the HSCs in vitro and GL reduces oxidative stress in HSCs. Mouse BM MNCs incubated with BL or GL in a serum-free medium for 72 h were stained with DCHFDA followed by antibodies to Sca-1, c-Kit, and lineage cocktail (N = 3). As seen in the panel
Interestingly, the lectin-incubated MNCs contained about 50% more KLS cells (1.2% in both BL and GL sets; Fig. 4A-a) than the control MNCs (0.8%), indicating that perhaps both BL and GL protect the stem cells under in vitro conditions as well [16 –18].
We examined ROS levels in M210B4 cells incubated with lectins or insulin. As seen in Figure 4C, the ROS levels in the treated cells were significantly less as compared to the control cells and the effect was sensitive to the presence of 20 mM α-methyl-
Discussion
Lectins have been used as cell separation and characterization reagents [45 –49] as they often bind only to restricted cell surface markers. Lectin microarrays have been shown to be an effective tool for analyzing diverse cell processes including cell development and differentiation, cell–cell communication, pathogen–host recognition, and cell surface biomarker identification [4,50]. Their usefulness as anticancer and antitumor agents by both oral and parenteral administration has been recently recognized [51 –53]. For several years, the lectins have been considered as toxic substances causing allergies, nutritional deficiencies, and other discomforts [5,11], yet they form an integral part of the vegetarian diet. Since one cannot avoid their exposure, it becomes imperative to examine their effect on various tissues and make a distinction between the ones having beneficial effects and the ones that could be potentially harmful.
Hematopoietic tissue serves as a sensitive indicator of health. This tissue harbors stem cells, whose function is indispensable for tissue regeneration and repair throughout the life of an individual. In the present work, we have made a systematic effort to examine the effect of two lectins that have been isolated from commonly consumed food items, namely banana and garlic, on the hematopoiesis of mice. Importantly, both these items are commonly consumed in an unprocessed form. Our results clearly show that both lectins have a salutary effect on the hematopoiesis. Since the doses used in the present study are very low, these lectins seem to have a potential to act as “micronutrients” for hematological disorders.
Survival and proliferation of the cells have been linked to activation of ERK pathway in several cell systems [34,54 –56]. The larger size of the CFU-F colonies formed from the marrows of the lectin-fed mice indicates that the lectins stimulated the stromal progenitors in vivo. The marrow MNCs incubated with lectins or with insulin also showed significantly higher CFU-F formation in an MEK-dependent way, indicating that insulin receptor signaling has a salutary effect on the stromal cells. In M210B4 cells, BL showed almost identical pattern of ERK activation—both in terms of kinetics and extent—to that of insulin, while GL showed a milder activation in terms of the extent, though the kinetics was similar to that of insulin. The most notable point was the transient nature of the activation (Fig. 3B), indicating that the lectin-induced signaling was amenable to feedback regulation. In several instances, the resistance of the lectin-induced signaling to feedback regulation has been shown to cause a problem in the clinical application of lectins [57].
ERK activation has been linked to both osteogenic as well as to adipogenic differentiation of marrow-derived stromal cells [58,59]. It will be interesting to examine whether the lectins induce a lineage-specific gene expression in the stromal compartment in vivo. Such studies would show whether the salutary action of the lectins on HSPC pool is a consequence of their ability to target the niche.
Yet another interesting observation that we made in this study was the differential interaction of these lectins with insulin receptors. While BL interacted with both pro-insulin as well as with insulin receptor β of M210B4 cells, GL interacted with only pro-insulin receptor. Since GL showed better effect on the primitive HSPCs, it appears that a milder level of ERK activation in the stromal cells as evoked by GL through pro-insulin receptor, rather than a high-level activation as evoked by BL and insulin through insulin receptor, may be more supportive of the primitive HSPC expansion. Such stem cell regulation by a differential insulin receptor signaling may have a bearing on stem cell dysfunction seen in various pathological conditions, including diabetes. This possibility needs to be formally addressed.
Aging is linked to the exposure to ROS and it has consequences in the stem cell regenerative function, including their homing and engraftment [60]. It is realized that the microenvironmental cells or “niche” regulate stem cell aging [44] and a healthy niche fosters the development of superior stem cell pool. In our experiments the antioxidant effect of the lectins on the stromal cells was quite striking, and it was not mediated via p38 pathway (data not shown), indicating that a p38-independent pathway was involved in the process. GL exerted a more direct effect on the ROS levels in the KLS population. Our data have unraveled a hitherto unrecognized antioxidant property of the lectins.
Diet restriction has been shown to delay HSC senescence [61]. Though long-term studies will be necessary to confirm the effects of these lectins on stem cell aging, our data suggest a possibility that the dietary components like BL and GL may delay the aging of HSPCs through the reduction in the ROS levels, and thus, “diet selection” or “diet enrichment” may be a helpful approach to protect the stem cells from aging. It would be indeed extremely important to see the effect of these lectins on other tissue-specific stem cells [62]. Pro-insulin receptor-mediated signaling has been shown to prevent apoptosis in embryonic stages [63,64], thus it would be important to examine whether this is also true for the stem cells.
In summary, our results clearly show that an oral administration of banana lectin and garlic lectin has a salutary effect on the HSPCs and suggest that this effect could be mediated by activation of ERK signaling and reduction of oxidative stress in the stromal cells.
Footnotes
Acknowledgments
The authors wish to thank the Department of Biotechnology, New Delhi, India, for the research grants, Ms. Nikhat Siddiqui for technical help in the animal experiments, and Mr. Swapnil Walke for help in acquisition of samples on the flow cytometer.
A part of the data was presented in the form of a poster by A.H. in 4th ISSCR Annual Meeting held at Toronto, Canada (June 29–July 1, 2006).