
Abstract
The success rate of human embryonic stem cell (hESC) derivation depends on both culture conditions and embryo quality and is routinely determined by morphological criteria. However, high incidence of chromosomal abnormality even in high-grade cleavage embryos from in vitro fertilization (IVF) patients suggests that the morphological grade of supernumerary embryos obtained from IVF clinics may not be a good prediction factor for successful hESC derivation. We show here that from one donor under identical derivation conditions 12 karyotypically abnormal post-bioptic embryos did not yield hESC lines, whereas two out of four normal embryos did. This suggests that the capacity of embryos to give rise to hESC line is likely to be influenced by their genetic status.
Introduction
C
A widely accepted procedure of preimplantation genetic diagnosis (PGD) allows detection of specific genetic defects in an in vitro fertilized embryo prior to transfer to the uterus for implantation [8 –10]. Although originally proposed in the 1960s, this technique was first applied in a clinical setting in 1990 to detect the sex of an embryo with X-linked disorder [11]. Diagnosis of a single-gene disorder, cystic fibrosis, followed 2 years later [12].
Detection of aneuploidy using fluorescent in situ hybridization is routinely performed on embryos from women of advanced maternal age or with a history of recurrent abortion. Only embryos that are diagnosed as normal using the biopsied blastomere are selected for transfer. This procedure has resulted in increased success of implantation [5,13] and ongoing pregnancy rates as well as in a reduced rate of spontaneous abortions [14].
On the other hand, data collected from number of studies suggested that most of the abnormal human preimplantation embryos analyzed were likely to be mosaics and not truly aneuploid (reviewed in [15]). When at least two cells were analyzed per embryo, overall lower rates of aneuploidy and higher rates of mosaicism were reported [16]. Subsequent studies have also shown a high degree of mosaicism at cleavage stage embryos regardless of the technique employed (eg, [17,18]). Furthermore, derivation of seven normal euploid hESC lines from 74 fresh PGD-defined aneuploid embryos has been reported [19]. The data suggested that: (1) the euploid hESC lines originated from mosaic embryos consisting of aneuploid and euploid cells and (2) in vitro selection favored euploid cells. Since the ratio of true aneuploid and mosaic embryos in this study is unknown, it is impossible to predict efficacy of normal hESC derivation from mosaic embryos and the actual percentage might be even higher than it appears. Also, since the embryos were from multiple donors and were subjected to hESC derivation at various time points, the influence of differences in genetic background and variations in culture conditions may play a role.
Recently we had an opportunity to shed more light on this issue by minimizing differences in genetic background and variations in culture conditions. A couple undergoing IVF wanted normal euploid male embryos to be kept, all abnormal, male and female, as well as normal female embryos were donated for research with their informed consent.
Using these embryos, we found that among four normal and 12 aneuploid morphologically good-quality embryos (grade I and II) only normal ones generated hESC lines lending support to the suggestion of a marked effect of an embryo’s genetic status on efficiency of hESC derivation.
Materials and Methods
hESC derivation and culture
All embryos used in these experiments were transported fresh from the IVF laboratory to the derivation laboratory in Human Tubal Fluid medium (Irvine Scientific, Santa Ana, CA) supplemented with 12 mg/mL human serum albumin (Irvine Scientific). Embryos used in these experiments were embryos donated by consenting patients according to the Institutional Review Board and Stem Cell Research Oversight Committee approved protocol.
Donated post-PGD embryos were cultured in 35 µL drops of G-2 v3 Plus medium (Vitrolife, Goteborg, Sweden), four per drop, under layer of oil (Irvine Scientific) for 2 days. At day 6 after fertilization, the zona pellucida was removed using Acid Thyrode (Irving Scientific) where required. Whole hatched embryos were seeded onto γ-irradiated human foreskin fibroblasts (HFF), population doubling (PD) 14, in hESC medium (80% KO-DMEM, 20% KO-SR, 25 ng/mL bFGF) and incubated at 37°C, in 5% CO2 and 8% O2. The medium was refreshed every 48–72 h. Undifferentiated hESC colonies were manually dissected and propagated on HFF PD14 in hESC medium until freezing. To examine their pluripotency, cells were cultured in hESC medium supplemented with 5% fetal calf serum (Hyclone) and without bFGF, for 2–14 days. The medium was replaced on a daily basis and the cells were not subcultured during the differentiation period.
Alkaline phosphatase assay
The assay was performed according to manufacturer’s instructions (ATCC).
Immunostaining
Undifferentiated or in vitro differentiated hESC cultures were fixed in 90% acetone or 3.8% paraformaldehyde and then permeabilized in 90% acetone and incubated with the primary antibodies recognizing: SSEA-4, TRA-1-60,TRA-1-81 (Millipore, Billerica, MA), nanog (Abcam, Cambridge, MA), Oct-3/4 (Santa Cruz Biotechnology, Santa Cruz, CA), smooth muscle actin (SMA), βIII-tubulin, or α-fetoprotein (αFP; Sigma). All secondary antibodies were purchased from Jackson ImmunoResearch, while Hoechst 33342 was obtained from Invitrogen.
Methylation-specific PCR
Bisulfite conversion of genomic DNA was performed using EZ DNA Methylation Kit (Zymo Research, Orange, CA). Primers were designed to discriminate between directly methylated and unmethylated alleles after treatment with sodium bisulfite. Normal and bisulfite-converted human placental DNA (Sigma) was used as a control. Primer sequences and PCR have been described previously [20,21].
Karyotyping
Aneuploidy screening was performed at PGD Science Inc., Woodland Hills, CA. Karyotypic analysis of early and later passages of each hESC line was performed by certified Cytogenetics Laboratory, Children’s Hospital and Research Center Oakland, CA.
DNA profiling
DNA profile [22] for each hESC line was determined by DNA testing laboratory Identigene, Houston, TX.
Results
Twenty-five embryos were screened for chromosome 13, 18, 21, X, and Y aneuploidy. Among 16 unwanted post-PGD embryos (Fig. 1) donated for research, four were normal (25%), while 12 (75%) were diagnosed as aneuploid. All four normal embryos were female. Four aneuploid embryos were male (33.3%) and eight female (66.7%) (Table 1). After 3-day culture in blastocyst G-2 v3 medium, all aneuploid embryos remained developmentally arrested or deteriorated showing extensive cell fragmentation and death. Three out of four embryos with a normal karyotype developed further (75%). Embryo R5.3 hatched spontaneously after PGD, embryo R5.2 started hatching, and embryo R5.5 remained within zona pellucida (Fig. 2A). Two days after transferring onto γ-irradiated HFF PD14 feeder layer, all three embryos attached and formed outgrowths (Fig. 2B). Initial trophectoderm outgrowths were disaggregated with a fine point needle on day 4 upon plating [23]. After the dissection of R5.2 colony, none of the small clumps proliferated further indicating that all of the cells had differentiated into trophectoderm. However, 9 to 11 days after seeding, clumps from R5.3 and R5.5 colonies generated outgrowths that resembled hESC colonies (Fig. 3A,B). On day 16 upon plating, hESC-like outgrowths were dissected again and pieces were transferred into a new feeder-containing well. The majority of the cell clumps attached and started to grow forming typical hESC colonies several days later (Fig. 3C).
Abnormalities are indicated with boldface.
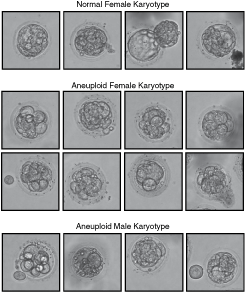
Sixteen unwanted post-preimplantation genetic diagnosis embryos donated for research by a consenting couple.
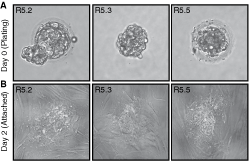
Hatching and plating of embryos. (
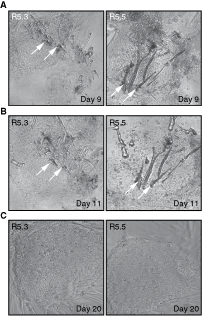
Forming initial human embryonic stem cell (hESC) colonies. Initial trophectodermal outgrowths were disaggregated with a fine point needle on day 4 upon plating [17]. After dissection of R5.2 colony, none of small clumps proliferated further, indicating that all cells were trophectoderm. However, 9 to 11 days upon plating, clumps from R5.3 and R5.5 colonies generated outgrowths that resembled hESC colonies (
The newly derived cell lines had a characteristic morphology of hESC. We used enzymatic assays (alkaline phosphatase/AP activity) and immunostaining (Oct-3/4, nanog, SSEA-4, TRA-1-60, and TRA-1-81) to confirm expression of pluripotency markers (Fig. 4). Differentiation into derivatives from all three embryonic germ layers has been confirmed by immunostaining of in vitro differentiated cultures. SMA was used as a marker of mesoderm, αFP as a marker of endoderm, and βIII-tubulin as a marker of ectoderm derivatives (Fig. 5). Pluripotency of the derived lines was also validated and confirmed in vivo. Both R5.3 and R5.5 hESC differentiated into a variety of cells of the three germ layers and yielded teratomas in immunocompromised mice (Fig. 5).

Both R5.3 and R5.5 human embryonic stem cell (hESC) lines express markers of pluripotency. AP activity and immunostaining (Oct-3/4, Nanog, SSEA-4, TRA-1-60, and TRA-1-81) confirm pluripotency of both R5.3 and R5.5 hESC lines.
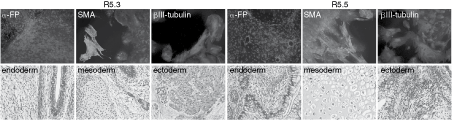
Both R5.3 and R5.5 human embryonic stem cell (hESC) lines can differentiate into all three germ layers in vitro (upper panels) and in vivo (lower panels). Upper panels: differentiation by αFP expression, Mesodermal differentiation was confirmed by demonstrating expression of SMA, endodermal, and ectodermal differentiation by βIII-tubulin expression. Lower panels: areas of teratomas in which cells differentiated into tissues of endodermal, mesdoermal, and ectodermal origin.
During gametogenesis maternal and paternal chromosomal alleles undergo differential epigenetic modifications, including DNA methylation that inactivates one or both copies of certain genes. The resulting unique inheritable inactivation pattern of each allele is termed genomic imprinting. Therefore, allelic expression of an imprinted gene depends upon whether the particular allele originates from a male or female gamete [24]. Inaccurate DNA inactivation during imprinting leads to imprinting disorders [25,26]. While the absolute risk of an imprinting disorder after assisted reproductive technology (ART) is very small, it is about 9-fold higher than in normal population [27]. Moreover, several studies have shown that genomic imprinting might be vulnerable in some hESC lines [20,21,28 –30]. Therefore, using methylation-specific PCR, we assessed the epigenetic stability of R5.3 and R5.5 hESC lines for three different regions: H19 and Oct-3/4 promoters, and DLK1/MEG3 region (Fig. 6). We detected amplicon using primers specific for either methylated or unmethylated DNA in both H19 promoter and DLK1/MEG3 region. Our data showed that both H19 promoter and DLK1/MEG3 region were heavily methylated on the paternal and unmethylated on maternal homolog, thus confirming their normal imprinting status. Furthermore, Oct-3/4 promoter was exclusively unmethylated in undifferentiated hESC from both R5.3 and R5.5 lines. Differentiation in both lines resulted in its methylation, although the DNA band representing unmethylated/active promoter still persisted. The pattern suggested normal and stable epigenetic regulation during differentiation.
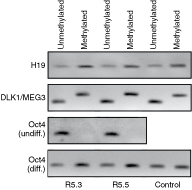
Methylation-specific PCR analysis of H19 promoter, DLK1/MEG3 region and Oct-3/4 promoter in R5.3 and R5.5 human embryonic stem cell (hESC) lines. In both lines H19 promoter and DLK1/MEG3 region showed normal methylation pattern: unmethylated maternal (137 bp band for H19 and 120 bp band for DLK1/MEG3) and methylated paternal (136 bp band for H19 and 160 bp band for DLK1/MEG3) allele. Oct-3/4 promoter showed unmethylated (181 bp) allele in undifferentiated and predominantly methylated (181 bp) allele in differentiated hESC. Normal and bisulfite-converted human placental DNA was used as a control.
As determined by GTG-banding, both R5.3 and R5.5 hESC lines have shown a stable normal female karyotype (46, XX) over 2.5 months of continuous culture (Fig. 7). Analyses of 15 short tandem repeat (STR) loci indicated unique DNA profiles of both lines eliminating any chance of cross-contamination with other hESC lines present in the laboratory (Table 2).
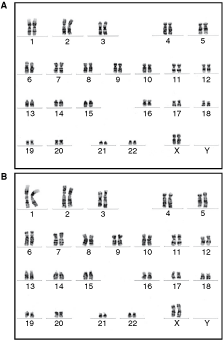
Cytogenetic analysis. Both R5.3 and R5.5 human embryonic stem cell lines showed normal stable female karyotype over 2.5 months of continuous culture.
Discussion
PGD-tested embryos were preferentially used to produce genetically distinct hESC lines with monogenic disorders [31]. hESC lines carrying mutations for myotonic dystrophy 1, Huntington disease, and cystic fibrosis have been derived and characterized [32,33]. Derivation, but not full characterization, of multiple hESC lines with various genetic disorders including adrenoleukodistrophy, fragile-X syndrome, Marfan syndrome, Duchenne and Becker muscular dystrophy, Fanconi anemia, complementation group A disorder, neurofibromatosis type I, and thalassemia has also been reported [34].
Although none of the aneuploid embryos in this particular set of embryos progressed further in development, we cannot negate a probability of hESC derivation from embryos carrying chromosomal abnormalities as determined with PGD aneuploidy screening (PGD-AS). At least two groups of researchers have reported derivation of normal euploid hESC lines from PGD-diagnosed aneuploid embryos [35,36]. High frequencies of mosaicism of up to 70% have been reported in 2–8 cell preimplantation embryos and therefore, PGD-AS diagnosis performed on a single blastomere clearly cannot distinguish between mosaic and nonmosaic embryos [15,37 –42]. In the most cases, under increasing cell cycle control of the developing embryo, abnormal cells undergo mitotic arrest and cell death. As a result, mosaicism disappears in >90% of mosaic embryos that reach the morula stage [42]. Keeping this in mind, it is easy to explain successful derivation of six normal hESC lines out of nine embryos with chromosomal aneuploidies according to PGD-AS [35,42]. As aneuploid blastomeres are highly likely to undergo cell cycle arrest and normal ones likely to continue to divide, the most advanced post-PGD embryos from multiple donors (>20) used in the study might be mosaics with the highest likelihood of having lost their aneuploid blastomeres. Concordantly, the authors reported a higher hESC derivation rate from PGD-analyzed, aneuploid, developmentally advanced embryos than from clinically unsuitable non-PGD embryos.
Approximately 2% frequency of mosaicism encountered at prenatal diagnosis in chorionic villi of vital pregnancies indicates that all aneuploid blastomeres do not undergo cell cycle arrest and that mosaic embryos may develop beyond 2–8 cell stage. However, mosaicism or homogeneous chromosomal abnormalities are in some cases present only in the extraembryonic tissues, with the fetus being normal [43 –47]. Pregnancy in which aneuploid cells are part of the fetus is apparently spontaneously aborted within the first trimester.
In our experiments, we could not exclude a possibility that one or more of embryos diagnosed by PGD to be either normal or aneuploid were mosaics. However, since all PGD-AS-diagnosed aneuploid embryos remained developmentally arrested, it is likely that they were either carrying homogeneous chromosomal abnormalities and thus, were not mosaics, or that they were mosaics with aneuploidy present in the majority of cells that caused developmental arrest of the whole embryo.
Since all embryos employed in our study were from the same donor, harvested, and processed together at the same time, the variabilities between individual embryos that may be caused by differences in genetic background as well as in culturing conditions were reduced to a minimum. Based on the outcome of such a unique set of embryos, one could reasonably argue that there is a dramatic difference in efficacy of hESC derivation from normal and from aneuploid embryos. Given the fact that more than half of 2–8 cell stage embryos harbor chromosomal abnormalities [1 –3,15] and that there is no morphological distinction between normal and aneuploid embryos of the same grade, the efficacy of hESC derivation from grade I and II embryos is actually much higher than calculated based on the total embryo number. In fact, it matches the chance of a live birth following conception, which is estimated to be between 22% and 50% in humans [4,48 –50].
Although we were unable to derive hESC line from developmentally arrested embryos, at least two groups have demonstrated that it is possible to obtain hESC lines even from such embryos [51,52]. The efficacy of derivation reported by both groups was very low (<1%) and it is likely that if we used comparable large number of developmentally arrested embryos, we would also achieve similar results. Zhang et al. derived under identical conditions six hESC lines from a total of 29 nonarrested embryos (20.7% efficacy) and only one hESC line, NCL9, from 132 developmentally arrested embryos [51]. Feki et al. described derivation of one hESC line from 125 arrested embryos. The embryo from which a line has been derived contained only one live blastomere. However, karyotype analysis of the line showed abnormalities in chromosomal number (61, XX) and the presence of isochromosomes [52]. The authors hypothesized that polyploidy was acquired very early, was possibly already present in the surviving blastomere, because the first karyotype was performed at passage 3. In that context, it is worth mentioning that it is also possible to derive normal hESC from single blastomeres as we and others have reported [53 –55].
In conclusion, previous publications show that it is feasible to derive genetically normal hESC lines from poor quality embryos or those deemed abnormal by cleavage stage PGD-AS. Our results suggest that efficiency of such derivations might depend on genetic status of the embryo. Taken together, these data implicate that none of the surplus embryos donated for research should be discarded, regardless of their quality or PGD-determined aneuploidy status. Even poor quality or PGD-aneuploid embryos may yield hESC lines either genetically normal or those bearing genetic abnormalities. In the process we will also enhance our understanding of genetic aberrations and their possible involvement in inducing embryo arrest.