
Abstract
Studies revealed that PI3K/AKT/mTOR signaling is important in the regulation of human embryonic stem cell (hESC) self-renewal and differentiation. However, its action on osteogenic differentiation of hESCs is poorly understood. We tested the effects of pharmacological PI3K/AKT/mTOR inhibitors on their potential to induce osteogenic differentiation of hESCs. Under feeder-free culture conditions, rapamycin (an mTOR inhibitor) potently inhibited the activities of mTOR and p70S6K in undifferentiated hESCs; however, LY294002 (a PI3K inhibitor) and an AKT inhibitor had no effects. Treatment with any of these inhibitors down-regulated the hESC markers Oct4 and Nanog, but only rapamycin induced the up-regulation of the early osteogenic markers BMP2 and Runx2. We also observed that hESCs differentiated when treated with FK506, a structural analog of rapamycin, but did not exhibit an osteogenic phenotype. Increases in Smad1/5/8 phosphorylation and Id1–4 mRNA expression indicated that rapamycin significantly stimulated BMP/Smad signaling. After inducing both hESCs and human embryoid bodies (hEBs) for 2–3 weeks with rapamycin, osteoblastic differentiation was further characterized by the expression of osteoblastic marker mRNAs and/or proteins (osterix, osteocalcin, osteoprotegerin, osteonectin, and bone sialoprotein), alkaline phosphatase activity, and alizarin red S staining for mineralized bone nodule formation. No significant differences in the osteogenic phenotypes of rapamycin-differentiated hESCs and hEBs were detected. Our results suggest that, among these 3 inhibitors, only rapamycin functions as a potent stimulator of osteoblastic differentiation of hESCs, and it does so by modulating rapamycin-sensitive mTOR and BMP/Smad signaling.
Introduction
H
Progress has been made in the identification of various osteogenic inducers, and a basic understanding of osteogenic signaling has been achieved. However, improvements are still needed to develop a scalable, reliable osteogenic culture system. 17-β-Estradiol [12], leucine-rich amelogenin peptide (LRAP) [13], and soluble extracts from 58S sol-gel bioactive glass [14] have all been demonstrated to have a positive effect on the osteoblastic differentiation of ESCs.
The involvement of fibroblast growth factor (FGF) signaling in the osteogenic differentiation of mouse ESCs (mESCs) was demonstrated by up-regulating FGF receptor (FGFR) 1–4 at both the mRNA and protein levels during osteogenic culture conditions [15]. The osteogenic transcription factor Runx2 is a major target gene for bone morphogenic protein (BMP)/Smad signaling during osteogenic induction in various cell types. In addition, BMPs have been demonstrated to stimulate osteogenic differentiation by inducing the expression of Runx2 in both mESCs [16,17] and hESCs [3]. Homozygous PPAR-γ (a transcription factor that plays a role in the cell-fate decision between adipocytic and osteoblastic lineages)-deficient ESCs [18] and PPAR-γ siRNA-treated ESCs undergo differentiation into osteoblastic lineages [19]. While these findings have contributed to the development of an osteogenic culture system, their utility may depend on cell type and culture conditions.
Studies have shown that the phosphoinositide 3-kinase (PI3K) pathway plays important roles in proliferation, survival, and the maintenance of pluripotency in ESCs [20,21]. mTOR (mammalian target of rapamycin), a major downstream target of PI3K/AKT, is ubiquitously expressed in various tissues and cell types [22,23], including undifferentiated and differentiated ESCs [24]. It regulates various cellular functions required for cell growth and proliferation by phosphorylating a series of substrates that include p70 S6 kinase (p70S6K1) and eukaryotic initiation factor 4E-binding protein 1 (4E-BP1) [22]. An essential role for mTOR in cell growth and proliferation was demonstrated in both early mouse embryos and ESCs by gene targeting technology [24]. Disruption of mTOR’s kinase domain and treatment with rapamycin both resulted in decreased proliferation of mouse ESCs [20,24].
Inhibiting mTOR signaling by rapamycin suppresses the proliferation of mESCs [25,26]. Rapamycin’s contribution to osteogenic differentiation has been demonstrated in various cell types [27,28], but the effect of rapamycin on the osteogenic differentiation of hESCs has not been addressed to date. Thus, we examined the effects of PI3K/AKT/mTOR inhibitors, including LY294002 (a PI3K inhibitor), AKT inhibitor, and rapamycin on hESC differentiation, with emphasis on differentiation into osteoblasts. We report that the mTOR inhibitor rapamycin is capable of differentiating cultured human ESCs toward an osteoblastic phenotype by blocking AKT, mTOR, p70S6K, and 4E-BP1 activation and stimulating BMP/Smad signaling.
Materials and Methods
Cell culture
H9 hESCs add (WiCell Res. Ins., WI) were maintained in an undifferentiated state using mechanical passaging as described previously [29]. After 5–6 days of culturing in hESC medium [80% DMEM/F12 medium, 20% knockout serum replacement (Invitrogen, Carlsbad, CA), 1% nonessential amino acids (Invitrogen), 1 mM
Western blot analysis
Cells were lysed on ice for 10 min using lysis buffer containing 50 mM Tris, pH 8.0, 150 mM NaCl, 1% NP-40, 0.1% SDS, 0.5% deoxycholic acid, 1 mM PMSF, and 1× protease inhibitor cocktail (Roche Applied Science, Indianapolis, IN) and centrifuged at 20,000g for 10 min at 4πC. Protein concentrations were determined using the BCA protein assay kit (Pierce, Rockford, IL). Equal amounts of protein (20 µg) were resolved by SDS polyacrylamide gel electrophoresis (PAGE). Gels were blotted to polyvinylidene fluoride (PVDF) membranes (Millipore Corp, Bedford, MA) and probed with one of the following primary antibodies, AKT antibody, p-Akt (Ser473) antibody, mTOR antibody, p-mTOR (Ser2448) antibody, p70S6K antibody, p-p70S6K (Ser389) antibody, 4E-BP1 antibody, p-4E-BP1 (Thr70) antibody, Smad antibody, p-Smad1/5/8 (Ser463/465, Ser463/465, Ser426/428) antibody, and β-actin antibody (Sigma Aldrich) for 1–2 h. After washing, membranes were probed with the corresponding secondary anti-rabbit HRP-conjugated or anti-mouse HRP-conjugated antibodies (Amersham, Arlington Heights, IL) at 1:10,000. Antibodies were purchased form Cell Signaling Technology (Beverly, MA) unless otherwise indicated. The bands were visualized by chemiluminescence using an ECL Advance kit (Amersham). Bands were analyzed for density with Image Gauge software (Fuji Photo Film GMBH, Düsseldorf) and normalized to loading control (β-actin) bands. All experiments were performed in triplicate. Error bars represent standard error of the mean (n = 3). Statistical significance was calculated using Student’s t-test (*P < 0.05, **P < 0.01).
Osteoblastic differentiation
LY294002 and AKT inhibitor were purchased from Calbiochem (La Jolla, CA). Rapamycin and FK506 were purchased from Sigma. Osteoblastic differentiation of hESCs was tested by culturing hESCs with or without an EB step. hESCs were cultured with MEF-CM in the presence or absence of the small molecular compounds (LY294002, AKT inhibitor, rapamycin, FK506) at various incubation times and concentrations, as depicted in Figure 1A. The medium was changed every 2 days. The osteoblastic differentiation of hEBs by inhibitors was tested as shown in Figure 2A. hEBs formed in suspension culture for 5 days were plated onto Matrigel-coated dishes. Attached hEBs were maintained in the presence or absence of inhibitors for 1–3 weeks with a media change every 2 days. Attached EBs were also cultured separately in medium containing osteogenic supplements as well as 0.1 mM
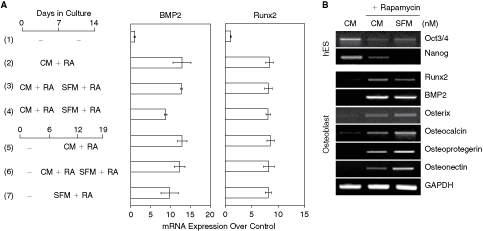
Effects of rapamycin on the osteoblastic differentiation of human embryonic stem cells (hESCs) cultured in serum or in serum-free conditions. (
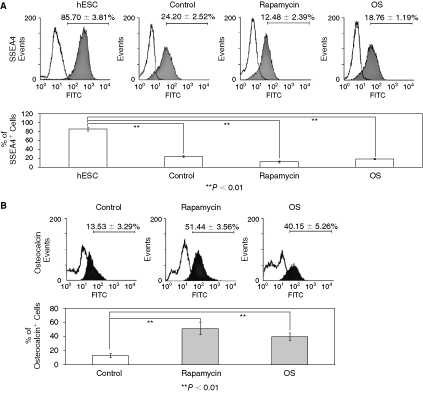
FACS analysis of rapamycin-differentiated human embryonic stem cells (hESCs). Undifferentiated H9 hESCs were maintained in mouse embryonic fibroblast (MEF)-conditioned medium plus 8 ng/mL of bFGF. Cells were incubated with MEF-CM without the bFGF supplementation in the absence (control) or presence of 1 nM rapamycin (rapamycin) or osteogenic supplements containing ascorbic acid (50 µg/mL), β-glycerophosphate (10 mM), and dexamethasone (100 nM) (OS) for 2 weeks. hESCs or hESC-derived differentiated cells were labeled with antibody to SSEA4 (
Analysis of mRNA levels
Total RNAs were extracted using Trizol (Invitrogen) following the instructions of the manufacturer. First-strand cDNAs were synthesized with the SuperScript First-Strand Synthesis System using 2 µg of RNA, oligo(dT), and SuperScript™ II reverse transcriptase according to the manufacturer’s protocol (Invitrogen). Synthesized cDNAs were amplified with 5× Taq-PCR Mix (Genotech, Daejeon, Korea) on GeneAmp PCR System 9700 (Applied Biosystems, Foster City, CA). PCR products were separated on a 1.5% agarose gel containing ethidium bromide and visualized with a Gel Doc EQ System (Bio-Rad, Watford, UK). Quantitative real-time PCR (qRT-PCR) was also performed using Power SYBR Green PCR Master (Applied Biosystems) and the AB 7500 Real-Time PCR System (Applied Biosystems). After Taq polymerase was activated at 95°C for 15 min, the reactions were denatured at 95°C for 15 s, annealed, and elongated at 58°C for 1 min for 50 cycles. All experiments were run in triplicate, and a C T value for each target gene was determined using the software provided by the manufacturer and normalized against the GAPDH expression level. Statistical differences between groups were assessed with Student’s t-test. A P value below 0.05 was considered to be statistically significant (*P < 0.05, **P < 0.01). Each pairs of primers are listed in Supplementary Table 1.
Alkaline phosphatase and alizarin red S staining
Cells were washed twice with PBS and fixed for 10 min by addition of 10% neutral buffered formalin (Accustain; Sigma). Alkaline phosphatase (ALP) activity was assessed using the ALP detection kit (Chemicon). Fixed cells were stained with naphthol/fast red violet solution for 15 min in the dark and then washed with phosphate-buffered saline (PBS) according to manufacturer’s instructions. For alizarin red S staining, fixed cells were incubated with 2% alizarin red S (Sigma) solution (pH 4.2) for 5 min, washed several times with distilled water, and air-dried for microscopic observation. Images were observed under an inverted microscope (Olympus, Tokyo, Japan).
Flow cytometry
hESCs or hESC-derived cells were dissociated into a single-cell suspension with 0.25% trypsin/EDTA solution (Invitrogen). The cell suspension was filtered through a 40-µm nylon cell strainer (BD Biosciences), and resuspended to about 5 × 105 cells in 100 µL PBS/0.1% BSA. For SSEA4 expression analysis, cells were incubated with primary antibodies including IgG isotype control (1 µg/106 cells) and SSEA4 (R&D systems, 1 µg/106 cells), diluted in PBS/0.1% BSA at 4°C for 30 min. For osteocalcin expression analysis, cells were fixed with 70% ethanol and permeabilized in 0.2% Triton X-100 solution. Subsequently, cells were incubated with IgG isotype control (1 µg/106 cells) and anti-osteocalcin antibody (R&D systems, 1 µg/106 cells). After washing, cells were incubated with fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse IgG (Santa Cruz Biotechnology, Santa Cruz, CA) at 4°C for 30 min. Cells were washed and analyzed on FACSCalibur™ flow cytometer (Becton Dickinson, San Jose, CA) using CellQuest™ software. Each analysis included at least 10,000 events. The percentage of SSEA4- or osteocalcin-positive cells was assessed after correction for the percentage of cells reactive to an isotype control conjugated to FITC. Statistical data expressed as mean ± SE. The data from FACS were evaluated by t-test.
Immunocytochemistry
Cells were washed twice with PBS and fixed for 10 min by addition of 10% neutral buffered formalin (Accustain; Sigma). After rinsing with PBST (PBS, pH 7.4, containing 0.05% Tween 20), cell membranes were permeabilized with 0.1% Triton X-100 solution (Sigma) for 30 min. After rinsing with PBST, cells were incubated with 4% BSA/PBST for 60 min and then with primary antibodies for osteocalcin (R&D Systems, Minneapolis, MN, USA) or bone sialoprotein II (Chemicon, Temecula, CA, USA) at a dilution of 1:500 or 1:1,000 in 3% BSA/PBST overnight at 4°C. Cells were washed with PBST, incubated with Alexa Fluor 488 chicken anti-mouse or rabbit IgG (Molecular Probes, Eugene, OR, USA) secondary antibodies in 3% BSA/PBST for 60 min at room temperature. After rinsing with PBST, cells were stained with DAPI (Molecular Probes) for 5 min and then mounted with ProLong Gold antifade reagent (Molecular Probes).
Fluorescence was detected using an Axiovert 200M multifluorescence imaging system (Carl Zeiss, Oberkochen, Germany) with AxioVision software.
Results
Rapamycin, an mTOR inhibitor, inactivates PI3K-AKT and mTOR-dependent pathways in hESCs
We first evaluated the sensitivity of hESCs to inhibitors of PI3K (LY294002), AKT (AKT inhibitor), and mTOR (rapamycin) by monitoring the levels of mTOR and p70S6K phosphorylation. Undifferentiated H9 hESCs were maintained in mouse embryonic fibroblast-conditioned medium (MEF-CM) under feeder-free conditions and treated with increasing concentration of inhibitors for 10, 15, or 30 min. Whole cell lysates prepared from undifferentiated hESCs and inhibitor-treated hESCs were analyzed by western blotting using antibodies against mTOR, phospho-mTOR (p-mTOR), p70S6K, phospho-p70S6K (p-p70S6K), and β-actin (as a loading control). Results showed that undifferentiated hESCs expressed active components of the mTOR pathway, such as p-mTOR, and p-p70S6K (Fig. 3A and 3B). LY294002 (5 µM) and AKT inhibitor (0.5 µM) did not alter the levels of mTOR, and p70S6K phosphorylation, suggesting that these inhibitors have no significant effects on mTOR-dependent pathways in hESCs. After 10 to 30 min in a low concentration of rapamycin (1 nM), the mTOR pathways were potently suppressed in hESCs, as demonstrated by decreases in mTOR and p70S6K phosphorylation (Fig. 3A and 3B). The total protein levels were not significantly changed by inhibitor treatment. We also examined the effects of FK506, a structural analog of rapamycin; as shown in Figure 3B, FK506 (10 nM) did not alter the levels of p70S6K or 4E-BP1 phosphorylation.

Effects of pharmacological PI3K/AKT/mTOR pathway inhibitors on mTOR and p70S6K activities. H9-undifferentiated human embryonic stem cells (hESCs) cultured for 5–6 days were treated either with DMSO (control; 0.05%), LY294002 (PI3K inhibitor; 5 µM), AKTi (AKT inhibitor; 0.5 µM), rapamycin (mTOR inhibitor; 1 nM), or FK506 (a structural analog of rapamycin; 10 nM) in mouse embryonic fibroblast-conditioned medium (MEF-CM) for 10–30 min. (
Rapamycin induces the expression of the early osteogenic markers Runx2 and BMP2 in monolayer-cultured hESCs after 1 week of culture
In an attempt to understand the effects of PI3K, AKT, and mTOR inhibitors on hESCs, hESCs were maintained in MEF-CM supplemented with or without LY294002 (5 µM), AKT inhibitor (0.5 µM), rapamycin (1 nM), or FK506 (10 nM), and hESC morphology and marker expression were monitored. Under all treatments, hESCs lost their normal morphology, displaying a looser colony structure after 5 days (data not shown). At the same time, decreases in the expression of hESC-specific marker genes, including Oct3/4 and Nanog mRNA, were observed by semiquantitative RT-PCR analysis (Fig. 4A). These data indicate that activation of the PI3K-AKT and mTOR pathways is required for the maintenance of hESC self-renewal and that their inactivation leads to their differentiation.
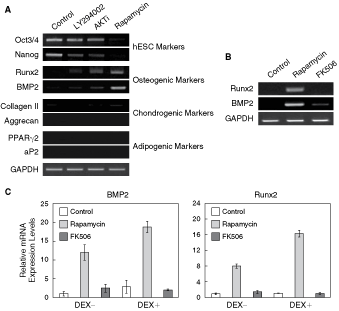
Effects of pharmacological PI3K/AKT/mTOR pathway inhibitors on the osteogenic differentiation of human embryonic stem cells (hESCs). H9 hESCs were cultured in mouse embryonic fibroblast (MEF)-conditioned medium containing DMSO control (0.05%), LY294002 (5 µM), AKTi (0.5 µM), rapamycin (1 nM), or FK506 (10 nM) for 5 days. (
Our observations led us to ask whether there is lineage commitment during hESC differentiation induced by these inhibitor treatments. Stimulatory or inhibitory effects of rapamycin on osteogenic differentiation have been demonstrated in various cell types [25,27,30] but not in hESCs. Accordingly, we examined whether these inhibitors promote differentiation of hESCs into osteogenic lineages by monitoring the appearance of osteoblast phenotypes.
After plating, monolayer-cultured hESCs were incubated in MEF-CM with or without LY294002 (5 µM), AKT inhibitor (0.5 µM), rapamycin (1 nM), and FK506 (10 nM) for 5–7 days. Commitment of hESCs to osteogenic lineages was evaluated by RT-PCR using primers for osteoblast lineage-specific mRNA. As a negative control, we also monitored the expression of chondrogenic and adipogenic markers during inhibitor treatment. Significantly, Runx and BMP2 transcripts, early markers of osteogenic differentiation were up-regulated in rapamycin-differentiated hESCs (Fig. 4A–4C) but not in hESCs treated with PI3K or AKT inhibitors (Fig. 4A). Under these conditions, the expression of chondrogenic or adipogenic markers was not observed, suggesting specific osteogenic induction by rapamycin (Fig. 4A).
Although rapamycin and FK506 bind to the same intracellular protein [termed FK506-binding protein (FKBP12)], they have distinct mechanisms of action and exert diverse cellular effects. When hESCs were treated with 10 nM FK506, BMP mRNA was slightly up-regulated, but Runx2 mRNA was not detected (Fig. 4B). A stepwise increase in concentration of FK506 (10 nM to 1 µM) failed to induce Runx2 mRNA expression. Real-time PCR analysis revealed that induction of Runx2 and BMP2 expression by rapamycin was enhanced by inclusion of 1 nM dexamethasone (DEX), an osteogenic supplement (Fig. 4C). However, FK506 in the presence of DEX did not significantly increase the expression level of BMP2 and Runx2 mRNAs in hESCs (Fig. 4C). These results suggest that rapamycin has the potential to be an osteogenic inducer of hESCs.
Rapamycin induces expression of the late osteogenic markers osteocalcin, osteoprotegerin, and osteonectin in monolayer-cultured hESCs after 2–3 weeks of culture with or without serum
To further confirm and optimize the osteogenic differentiation of hESCs induced by rapamycin, hESCs were plated onto Matrigel-coated culture dishes and treated with 1 nM of rapamycin either in MEF-CM, which contained 20% knockout serum replacement, or in serum-free medium (SFM) for different periods of time (2–3 weeks). Runx2 and BMP2 mRNA expression was significantly elevated from day 5 or day 7 up to day 21, with ∼8- and 12-fold induction, respectively. Treatment with rapamycin 1 day after plating significantly enhanced the osteogenic differentiation of hESCs but generated only a small number of differentiated cells. To obtain a large quantity of rapamycin-differentiated cells, hESCs were treated with 1 nM rapamycin from day 6 after plating up to 2–3 weeks, and expression of osteogenic marker mRNAs was confirmed by semiquantitative RT-PCR and real-time PCR. We found no significant difference in the expression levels of Runx2 and BMP2 between different treatment time periods (Fig. 1B). The mRNA levels of late osteoblast differentiation marker genes, including osteocalcin, osteoprotegerin, and osteonectin, were significantly enhanced after continuous exposure of hESCs to rapamycin both in MEF-CM and SFM (Fig. 1A and 1B). These results suggest that hESCs cultured in rapamycin-supplemented medium with or without serum undergo differentiation into osteogenic lineage without an EB formation step.
Rapamycin, but not FK506, induces in vitro differentiation of hESCs grown in a feeder-free culture into mineralized osteoblasts without an EB step
Osteoblastic differentiation of hESCs was further confirmed by bone-specific alkaline phosphatase (ALP) staining (Fig. 5A) and alizarin red S staining (Fig. 5B). ALP activity in rapamycin-differentiated hESCs was significantly lower after 1 week of culture. However, enzyme activity was markedly enhanced after culturing for >2 weeks (Fig. 5A). In comparison, hESCs treated with FK506, LY294002, or AKT inhibitor from 1 week up to 3 weeks did not display increased ALP activity. To detect mineralization of hESCs, cells were stained with alizarin red S (Fig. 5B). Mineralized bone nodules stained by alizarin red S were detected after 2 weeks in rapamycin-differentiated hESCs incubated either in MEF-CM or in SFM, and stained cells were increased in a time-dependent manner (Fig. 5B). Alizarin red S-stained cells were not seen in hESCs treated either with FK506, LY294002, or AKT inhibitor. FACS analysis of hESC populations showed that 51.44% ± 3.56% of rapamycin-differentiated hESCs and 40.15% ± 5.26% of osteogenic supplements (OS) (ascorbic acid, β-glycerophosphate, dexamethazone)-differentiated hESCs were found to express osteocalcin at 2 weeks of culture (Fig. 2B). However, the osteocalcin expression was not further increased with longer incubation times (3 weeks) (data not shown). Under the same conditions, the population of SSEA4-positive cells markedly decreased in rapamycin-differentiated hESCs (12.48% ± 2.39%), and OS-differentiated hESCs (18.76% ± 1.19%) compared to undifferentiated hESCs maintained in MEF-CM plus bFGF (85.70% ± 3.81%) at 2 weeks of culture (Fig. 2A). These results suggest that rapamycin alone promotes the osteoblastic differentiation of hESCs and, further, that mTOR, a downstream signaling molecule of the PI3K/AKT pathway, plays an important role in the modulation of osteoinductive signal in hESCs.
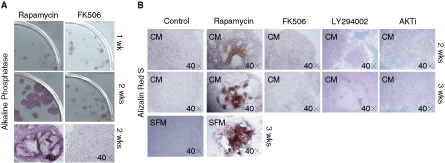
Characterization of human embryonic stem cell (hESC) osteoblastic differentiation after rapamycin induction. H9 hESCs were cultured in the presence of rapamycin (1 nM), FK506 (10 nM), LY294002 (5 µM), or AKTi (0.5 µM) for 2–3 weeks. (
Rapamycin, but not FK506, enhances the osteoblastic differentiation of hEBs
We also evaluated the osteogenic effects of rapamycin, FK506, LY294002, and AKT inhibitor in hEBs. Spherical cell aggregates of hEBs were formed by static and suspended incubation for 5 days (Fig. 6A, Stage I), attached to Matrigel-coated dishes, and grown in the presence or absence of inhibitors (Fig. 6A, Stage II). In the presence of rapamycin, attached hEBs displayed a rougher surface morphology (Fig. 6A), similar to that seen in rapamycin-differentiated hESCs.
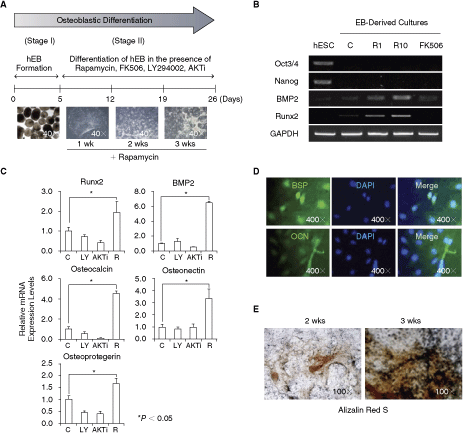
Characterization of osteoblastic differentiation of hESC-derived embroid body (hEB) induced by rapamycin. (
The expression levels of osteoblast marker genes in hEBs after treatment with inhibitors were tracked by RT-PCR or quantitative real-time RT-PCR analysis. Similar to the results observed for monolayer-cultured hESCs, rapamycin significantly enhanced the expression of osteoblast markers such as BMP2, Runx2, osteocalcin, osteonectin, and osteoprotegerin when hEBs were continually treated for 2 weeks (Fig. 6B and 6C). However, FK506, LY294002, and AKT did not alter the mRNA expression of those osteoblast marker genes (Fig. 6C). Along with the increases in mRNA expression of osteoblast marker genes, up-regulation of bone sialoprotein (BSP) and osteocalcin protein expression was determined by immunofluorescent staining with the respective antibodies (Fig. 6D).
Osteoblastic differentiation of rapamycin-induced hEBs was further confirmed by monitoring the formation of mineralized nodules positive for alizarin red S staining (Fig. 6E). Mineralized nodules were detected after 2 weeks of culture, and their formation reached the highest level after 3 weeks of culture (Fig. 6E). No mineralized nodules were observed in untreated control cultures or in FK506-, LY294002-, or AKT inhibitor-treated cultures. These results suggest that rapamycin alone is able to stimulate the differentiation of hEBs into mature osteoblasts.
Rapamycin-mediated osteogenic induction is related to BMP/Smad activity
In the presence of rapamycin, hESCs differentiated into osteoblasts, and this differentiation was accompanied by BMP2 mRNA up-regulation (Fig. 4). Correlatively, cross talk between mTOR and BMP/Smad signaling has been previously demonstrated [31]. Therefore, we speculated that induction of hESC osteogenic differentiation via antagonized mTOR signaling might be linked to Smad activity and transcriptional changes of its target genes. Western blot analysis showed that rapamycin significantly induced the phosphorylation of Smad1/5/8, a downstream target of BMP. Furthermore, addition of Noggin, a BMP antagonist [32], decreased the rapamycin-induced phosphorylation of Smad1/5/8 in hESCs. As a control, we observed that BMP2 significantly elevated the phosphorylation of Smad1/5/8, whereas BMP2-mediated phosphorylation of Smad1/5/8 was inhibited by treatment with Noggin, an antagonist of BMPs (Fig. 7A). Furthermore, RT-PCR analysis revealed that rapamycin induced the up-regulation of Id1–4, a direct target of BMP2 [32] after 2 weeks of culture. Id1–4 mRNAs were elevated after 1 week of hESC culture, but no further increase was observed (Fig. 7B). The expression of Id2 mRNA was increased in a time-dependent manner. Id3 mRNA levels were elevated after 2 weeks of hESC culture (Fig. 7B). These results demonstrate that rapamycin activates the BMP-dependent Smads and modulates their transcriptional activation of target genes, which is important for the osteogenesis of hESCs.

Effects of rapamycin on BMP/Smad signaling in human embryonic stem cells (hESCs). (
Discussion
A role for PI3K/AKT/mTOR signaling in the maintenance of ESC pluripotency and survival has been demonstrated both in mice [20,21,24,33 –37] and humans [38,39]. We have characterized the effect of rapamycin, a selective, small, molecular mTOR (mammalian Target Of Rapamycin) inhibitor, on the differentiation of hESCs into osteoblastic lineages. We found that rapamycin-induced osteoblastic differentiation of hESCs by stimulating BMP/Smad signaling.
Undifferentiated hESCs expressed the active phosphorylated form of mTOR and p70S6K (downstream of mTOR) (Fig. 3A and 3B). Blocking PI3K, AKT, or mTOR activity with their specific small molecular inhibitors resulted in hESC differentiation within a single passage under feeder-free condition. A previous study showed that activating PI3K via leukemia inhibitory factor (LIF), which supports ESC maintenance, is blocked by the PI3K-specific inhibitor LY294002 and leads to down-regulation of mouse ESC self-renewal [36]. Inhibition of PI3K by LY294002 induced inactivation of the downstream effectors including AKT and p70S6K, although only a marginal inhibitory effect on p70S6K activity was observed in undifferentiated mouse ESCs [36,37]. mTOR/p70S6K activities can be blocked by the PI3K inhibitor LY294002, but higher concentrations of LY294002 are usually required for mTOR/p70S6K inhibition than for PI3K inhibition. The mTOR inhibitor rapamycin was able to effectively block activation of the mTOR pathway in undifferentiated hESCs at a low concentration (1 nM) (Fig. 3). It is notable that both LY294002 (5 µM) and AKT inhibitor (0.5 µM) had no effect on mTOR activity in undifferentiated hESCs (Fig. 3A and 3B). Our results indicate that blocking the PI3K-AKT-mTOR pathway either at the PI3K/AKT level with LY294002 or AKT inhibitor or at the mTOR level with rapamycin significantly suppresses hESC self-renewal and promotes differentiation. Our results also indicate that those inhibitors have different impacts on downstream of mTOR-dependent pathway.
Although rapamycin and FK506 bind to the same intracellular protein FKBP12, they have distinct mechanisms of action and exert diverse cellular effects. The osteogenic potential of both rapamycin and FK506 has been demonstrated in various cell types. However, they exert both stimulatory and inhibitory influences on osteogenic differentiation depending on cell type and treatment conditions. Antagonizing mTOR by rapamycin reduced osteoblast-specific osteocalcin, bone sialoprotein, and osterix mRNA expression, ALP activity, and mineralization capacity in differentiating osteoblasts [25]. In contrast, rapamycin, but not FK506, stimulates osteoblastic differentiation in combination with vitamin D3 in rat osteoblast-like osteosarcoma (ROS 17/2.8) cells [27]. In contrast, FK506, but not rapamycin, stimulates osteoblastic differentiation in mesenchymal stem cells [30,40].
We observed that continuous treatment with rapamycin, but not LY294002, AKT inhibitor, or FK506, stimulated the osteogenesis of hESCs cultured for 5–7 days under feeder-free conditions, and this result was confirmed by up-regulation of the early markers for osteoblastic differentiation, Runx2 and BMP2 (Fig. 4A–4C). Conventionally, hESCs undergo differentiation into osteoblast lineages when cultured in the presence of osteogenic culture medium containing dexamethasone (DEX),
Continuous exposure to rapamycin induced the mRNA expression of the early osteogenic transcription factor Runx2 from 5 days up to 2 weeks (Figs. 1B and 3B), after which it declined. These results indicate that, at an early step of the differentiation process, rapamycin directs the differentiation of hESCs into early osteoblast lineage cells via a Runx2-dependent pathway. This pathway’s subsequent down-regulation may then provide favorable conditions for mature osteoblastic differentiation. Supporting this view, other studies showed that Cbfa1 (Runx2), an essential transcription factor for osteoblastic differentiation, promotes osteoblastic differentiation at the early stage and inhibits the late stage of osteoblastic differentiation [41,42].
Osterix is an osteogenic transcription factor that acts downstream of Cbfa1 and is required bone formation [43]. Along with the increase in Runx2 mRNA expression, expression of osterix mRNA was enhanced by rapamycin after 2 weeks of culture in a time-dependent manner. This enhanced Runx2 expression may mediate the induction of osteoblast-specific gene expression required for the production of a bone-specific matrix.
We also tested the osteogenic potential of rapamycin in hEB-derived cultures. Similar to their effects on hESCs, rapamycin, but not LY294002, AKT inhibitor, or FK506, potently stimulated the osteogenic differentiation of hEBs, as evidenced by the appearance of mineralized bone nodules (Fig. 6E) and the expression of the osteoblast-specific markers Runx2, BMP2, bone sialoprotein, OCN, osteoprotegerin, and osteonectin (Fig. 6B–6D). In as little as 2 weeks, bone nodule-like structures were found in hEB cultures but only in the presence of rapamycin. Formation of mineralized nodules increased in a time-dependent manner (Fig. 6E). This observation implies that, with or without the EB step, rapamycin stimulates osteoblastic differentiation of hESCs. Mineralized bone nodules, indicative of mature osteoblastic differentiation, appeared in differentiated cells only after 2–3 weeks of differentiation. In our experimental conditions, incubation with low concentrations of rapamycin (1–10 nM) are effective for osteogenic induction in both hESC monolayer cultures and hEB-derived cultures, while higher concentrations of rapamycin (≥50 nM) produced a significant decrease in cell viability and growth (data not shown).
BMPs, which belong to the transforming growth factor-β (TGF-β) superfamily, serve as osteogenic stimulators in various cell lines, including hESCs [3,16,44 –46]. However, in hESCs, BMP/Smad activity is also required for cardiac differentiation [47] and/or extra-embryonic endoderm differentiation [48] depending on culture conditions. Notably, up-regulation of BMP2 mRNA expression and Smad1/5/8 phosphorylation was observed in rapamycin-treated hESCs (Fig. 7A), indicating that the stimulatory effect of rapamycin on osteoblastic differentiation may be exerted through activation of the TGF-β type I receptor and/or BMP/Smad signaling. Activation of BMP/Smad signaling by rapamycin in hESCs was further evidenced by up-regulation of Id1–4 mRNA expression (Fig. 7B). Consistent with this observation, rapamycin has been shown to regulate TGF-β/BMP signaling by blocking the interaction of FKBP12 and the TGF-β type I receptor [49,50] and inducing BMP4 expression and Smad activity [31]. In addition, p70S6 kinase, a downstream effector of mTOR, is involved in BMP signaling [51], and its regulation is known to be important for the osteoblastic differentiation [26].
We conclude that inhibition of PI3K-AKT-mTOR signaling by rapamycin contributes to hESC commitment into osteoblastic lineages in vitro and, therefore, present rapamycin as a new osteogenic factor that stimulates the osteoblastic differentiation of hESCs. Although further clarification is needed to understand the cellular and molecular mechanisms by which rapamycin induces the osteoblastic differentiation of hESCs, our results provide useful insight into the distinct roles of mTOR signaling in regulating cell-fate decisions of hESCs, as well as into the possible biological responses of rapamycin as an immunosuppressant after allogenic stem cell transplantation.
Footnotes
Acknowledgments
We thank Dr. Hyung Min Chung for kindly providing CF1 mouse for MEF preparation. This work was supported by the KRF/KOSEF Stem Cell Research Program (2009-0084069) and the KRIBB Research Initiative Program.