
Abstract
Attempts to repair the central nervous system damaged as a result of trauma or disease will depend on the ability to restore the appropriate neuronal connectivity. This will rely on establishing appropriate chemical and physical environments for supporting neural cells and their processes and in this regard, engineering of biomaterials is of increasing interest. It will be important to understand how cells behave on these biomaterials in vitro, prior to future in vivo application. We reveal that modification of 3-dimensional (3D) electrospun poly-ε-caprolactone (PCL) nanofiber scaffolds by fiber alignment and aminolysation is superior to classical 2-dimensional (2D) culture-ware in promoting in vitro proliferation and differentiation of cortical cells. Many studies have examined the importance of exogenous soluble factors to promote cell fate specification. Here, we demonstrate that tethering the neurotrophin, brain-derived neurotrophic factor (BDNF), onto modified nanofibers is superior to culturing in the presence of soluble BDNF. Functional immobilization of BDNF to polymer nanofibers enhances neural stem cell (NSC) proliferation and directs cell fate toward neuronal and oligodendrocyte specification, essential for neural tissue repair. These findings indicate that modified PCL nanofibrous 3D scaffolds are capable of supporting NSCs and their derivatives and may present a new avenue for encouraging neural repair in the future.
Introduction
R
Previously we, along with others, have tested a number of different biomaterials, including hydrogels and electrospun nanofibers, in an attempt to model the native 3D in vivo environment [1 –6]. Electrospun nanofibers are superior to hydrogels in simulating the 3D in vivo environment of neural tissue because fiber alignment, diameter, and inter-fiber distance can be regulated to generate a surface more permissive for neural stem cell (NSC) adhesion and axon support [6]. Furthermore, nanofibers can be modified to provide 3D trophic support to cells. Functionalization with surface-bound amines on electrospun polymer nanofibers has previously been demonstrated to promote cell adhesion of both adult NSCs and hematopoietic stem cells. The mechanism for these superior characteristics is unclear, but enrichment with supportive proteins, binding and activation of negatively charged proteins, and changes in surface hydrophilicity have been proposed [4,7]. The biodegradability of PCL along with our previous preliminary findings [4], illustrating that neural cells were supported by electrospun PCL, has lead us to further examine the properties of various modified PCL scaffolds, using the support of NSCs and their derivates as a marker of biocompatibility.
Functional ligands are also important in tissue repair. While it is widely accepted that many of these ligands act at the surface rather than in a soluble form, until recently their actions in this context could not be studied. Examination of immobilized proteins was previously hindered by difficulties in stably attaching ligands to a surface while maintaining cellular accessibility. Consequently, these studies relied upon continual addition of the ligand, either to the culture media in vitro, or via invasive implantation of cannulas and osmotic pumps in vivo. Ligand immobilization onto a surface prevents endocytosis of the molecule, thereby prolonging the period of stimulation [8 –10].
There are now several examples of immobilized ligands promoting cell fate. Covalent attachment of mitogens (epidermal growth factors, EGF, or leukemia inhibitory factor) onto glass or thin polymer coatings promoted cell proliferation and maintained embryonic stem cells [11,12]; basic fibroblast growth factor gradients covalently attached to hydrogels promoted cell migration [13]; and the attachment of EGF to hydrogels promoted survival of mesenchymal stem cells [14].
The neurotrophins, including nerve growth factor (NGF), neurotrophin-3 (NT3), NT4/5 and brain-derived neurotrophic factor (BDNF) [15], are of particular interest with regards to promoting neuronal survival and connectivity in the context of tissue repair. Attempts have been made to immobilize neurotrophins onto glass or to incorporate them into hydrogels or microtubules; however, these surfaces lack the architectural benefits of nanofibrous scaffolds such as high surface areas for high-density presentation of ligands and structural similarities to the native basement membrane [16 –19]. The few attempts to immobilize ligands to nanofibers [20 –22] have focused predominantly on the engineering of the material itself with little attention to potential biological applications. In this regard, we wish to examine the effects of BDNF-nanofiber immobilization on cortical NSCs as a proof of concept of ligand immobilization for neural cells.
The reason BDNF was chosen for immobilization onto the nanofibers was its role in promoting survival of cortical neurons in vitro and in vivo and its well-known role in development of the CNS and amelioration of CNS and PNS injuries [23 –27]. In addition, BDNF is important for regulating cell proliferation and survival during oligodendrogenesis [28,29]. These oligodendrocyte progenitors are responsible for the production of myelin, critical for the rapid propagation of action potentials along nerve fibers and the repair of neural connections. Together, BDNF and NSCs (including neural and oligodendrocyte progenitors) can enhance CNS and PNS repair [27, 30 –32].
We compared the proliferation and fate specification of cortical NSCs on a number of modified PCL electrospun nanofibers, including nanofibers with tethered BDNF, with conventional 2-dimensional (2D) in vitro assay systems. We showed that 3D PCL nanofibers are capable of supporting NSCs and their derivative by enhancing cell numbers (presumably by increased proliferation) and differentiation. Most notable characteristics that supported neural cells were aminolysis of the PCL scaffold combined with immobilization of BDNF. These findings illustrate the ability of PCL nanofibrous scaffolds to support neural cells and illustrate the benefits of a number of modifications, including BDNF immobilization. These findings provide important insights into the potential use of functionalized biomaterials for future applications in tissue engineering to repair the injured nervous system.
Materials and Methods
Preparation of scaffolds
Electrospinning refers to a nanofabrication process whereby an electric field causes a droplet of a polymer solution at a tip of a needle to deform into a cone. At a critical voltage, the charge build up on the surface of the solution overcomes the surface tension and a fine jet forms. The jet of polymer solution undergoes instabilities caused by electrostatic repulsion that further elongates the stream. During this process, the solvent evaporates yielding a polymer fiber that can be deposited onto a rotating mandrel to provide 3D nanofibrous scaffolds [33]. Using this method, we generated 3D nanofibrous scaffolds from PCL.
PCL was obtained from Absorbable Polymers, Inc. (Alabama). Polymer solutions of 13% w/v were prepared by dissolving PCL in 2 mL of chloroform (Merck Pty Ltd., Australia) and methanol (Merck Pty Ltd., Australia) at a ratio of 3:1 (v/v). One mM of dodecyl trimethyl ammonium bromide was also added as a surfactant to facilitate the spinning process. The solutions were placed into a glass syringe (10 mL) with a 21-gauge needle for electrospinning and a flow rate of 0.794 mL/h was used. A 15-kV accelerating voltage was applied by a high-voltage power supply and a 15-cm working distance was utilized. The scaffolds were deposited on an aluminum mandrel with a diameter of 5 cm. Speeds of 200 rpm were used to fabricate randomly orientated fibers and 4,000 rpm to fabricate aligned fibers. The scaffolds were then examined using scanning electron microscopy (SEM). They were gold-coated using a sputter coater (Balzers SCD-005, BAL-TEC) with a 25-mA current for 180 s prior to imaging at 15 kV with the SEM (S-570 Hitachi Tokyo, Japan). The resultant electrospun fibers had a mean fiber diameter of 550 ± 100 nm.
Some scaffolds were subsequently aminolysed in 0.05 M ethylenediamine (ED) (Sigma-Aldrich Pty Ltd., St. Louis, MO) in 2-isopropanol (IPA; Sigma, Sydney, Australia) and allowed to react at 20°C for 10 min. For samples with polyethyleneimine (PEI) coatings, the scaffolds were submerged in 20 mg/mL of PEI (Mw 2,000; Sigma, Australia) in water for 1 h. The treated samples were then washed 3 times in ice water and soaked for 1 h in distilled water on ice.
Punch biopsies (0.6 cm in diameter) were made from each electrospun scaffold, with a material thickness of 150 ± 50 μm for aligned and 250 ± 40 μm for random scaffolds. BDNF was either added directly to the culturing media (30 ng/mL; R&D Systems, Minneapolis, MN) or anchored to the scaffolds prior to cell plating. For attachment of BDNF, sterilized ED-coated scaffolds were incubated in succinimidyl 4-(N-maleimidomethyl)-cyclohexane-1-carboxylate (SMCC) 5 mM, dissolved in PBS for 1 h for 2 h with agitation. Subsequently, the solution containing the scaffolds was replaced with BDNF (10 μg/mL) in PBS at 4°C for 10 h. Scaffolds were then washed 3 times for 10 min in PBS prior to seeding of NSCs.
Apart from the previously sterilized scaffold with BDNF tethered to the fibers, all remaining scaffolds were sterilized in 70% ethanol for 5 min followed by 3 washes in PBS (5 min), prior to seeding of the NSCs. Triplicates of each scaffold material were prepared, placed into the bottom of a single well of a 96-well culture-ware plate, and anchored to the bottom of the well using a perspex cylindrical insert (see Fig. 1A).

Modification of electrospun poly(ε-caprolactone) (PCL) nanofibers and brain-derived neurotrophic factor (BDNF) immobilization. (
Cortical cultures
All methods conformed to the Australian National Health and Medical Research Council published code of practice for the use of animals in research and were approved by the Florey Neurosciences Institute animal ethics committee. Embryonic day 14.5 (E14.5) pregnant mice were sacrificed by anesthetizing with isoflurane followed by cervical dislocation. The uterine horn was rapidly removed, placed on ice-chilled PBS, and sprayed with 70% ethanol. Subsequently, the embryos were removed and transferred to a fresh plate of chilled PBS. The cortex was isolated and meninges removed in serum free N2 medium [consisting of a 1:1 mixture of F12 and MEM, 15 mM HEPES buffer, 1 mM glutamine, 6 mg/mL glucose (Sigma), 3 mg/mL Albumax, and N2 supplement (all purchased from Invitrogen)] and containing 0.03% glucose. The isolated cortices were dissociated with polished Pasteur pipettes and seeded at a density 1.25 × 106 cells/mL in N2 medium containing epidermal growth factor (EGF, 10 ng/mL) and fibroblast growth factor-2 (FGF2, 20 ng/mL) to form neurospheres. After 5 days of expansion, the spheres were trypsinized into a single cell suspension and plated onto scaffolds or poly-
Immunocytochemistry
Immunocytochemistry was performed on 4% paraformaldehyde-fixed cultures. The following primary antibodies were used: mouse anti-βIII-tubulin, IgG (Tuj1, 1:2,000, Promega, Madison, WI; neuronal marker), rabbit anti-NG2 (1:500, Chemicon, Copenhagen, Denmark; a marker of neural progenitors commonly used to label oligodendrocyte progenitors [34]), rabbit anti-glial fibrillary acidic protein (GFAP, 1:800, DAKO, Carpinteria, CA), mouse anti-nestin (1:200, Chemicon), and rabbit anti-cleaved caspase-3 (cell death marker, 1:1,000, Cell Signaling, Danvers, MA).
Appropriate fluorophore-conjugated or biotinylated secondary antibodies (Alexa-488 and Alexa-594) were used for visualization. For chromogenic histochemistry, following incubation in the secondary antibody, coverslips/scaffolds were washed in PBS (3 times 5 min), incubated in avidin peroxidase (1:5,000, 1 h), and then reacted with cobalt-and nickel-intensified diaminobenzidine (DAB, Sigma Chemicals, St. Louis, MO). Hoechst nuclear stain (5 mg/mL, 1:5,000; Sigma) was performed for visualization of all cells in culture. Hoechst-labeled cells were scored as either live or dead based on nuclear morphology. Dead, apoptotic cells, whose nuclei were shrunken and contained collapsed chromatin were excluded from total cell counts.
For cell counts, triplicate glass or PCL scaffolds of each treatment were included for each immunological marker (nestin, TUJ1, GFAP, NG2, caspase-3) and repeated on quadruplicate-independent cultures. Stained scaffolds were mounted onto glass slides and coverslipped using DAKO-mounting media. Cell counts were performed on a Leica DM LB2 microscope (Wetzlar, Germany) using a 40× objective and equipped with Optronics color digital camera. To avoid counting bias, on each scaffold, the first counting field was taken as the third field of view from the left-hand side of the scaffold material. Five fields were sampled from each scaffold. Furthermore, the observer was blinded to the treatment groups.
BDNF ELISA
In parallel to scaffold preparation for in vitro culturing of cortical cells, additional scaffolds were prepared to confirm the attachment of BDNF. In brief, Random_ED, Align_ED, Random_ED_X-BDNF, and Align_ED_X-BDNF scaffolds were placed into individual well of a 96-well plate and washed 3 times in PBST (PBS containing 0.05% Tween-20) prior to blocking in 5% normal goat serum (20 min). The scaffolds were then incubated in rabbit anti-BDNF antibody (1:750 in PBST; Abcam, Cambridge, MA) for 2 h at 37°C. The wells were washed 4 times with PBST and incubated in 150 μL/well of horseradish peroxidase (HRP, 1:1,000)-labeled anti-rabbit immunoglobulin (DAKO, Denmark) for 1 h at 37°C. Bound HRP activity was assayed by color development using TMB microwell peroxidase system. The reaction of HRP was stopped using 0.3 M HCl acid, and absorbance at 450 nm was measured with a microtitre plate reader (SpectraMax). For ELISA, triplicate scaffolds of each treatment were included and repeated on 3 independent cultures.
Statistical analysis
Data was analyzed using Sigma Stat software (version 3.0). For analyses comparing 2 groups such as relative amounts of bound BDNF (Fig. 1), Student's t-tests were used. Where greater than 2 groups were compared (Figs. 2 –4), Holm-Sidak multiple comparison tests were used. Each graph value is the mean ± SEM. Statistical significance was set at the level of 0.05 (*P < 0.05, **P < 0.01, ***P < 0.001).
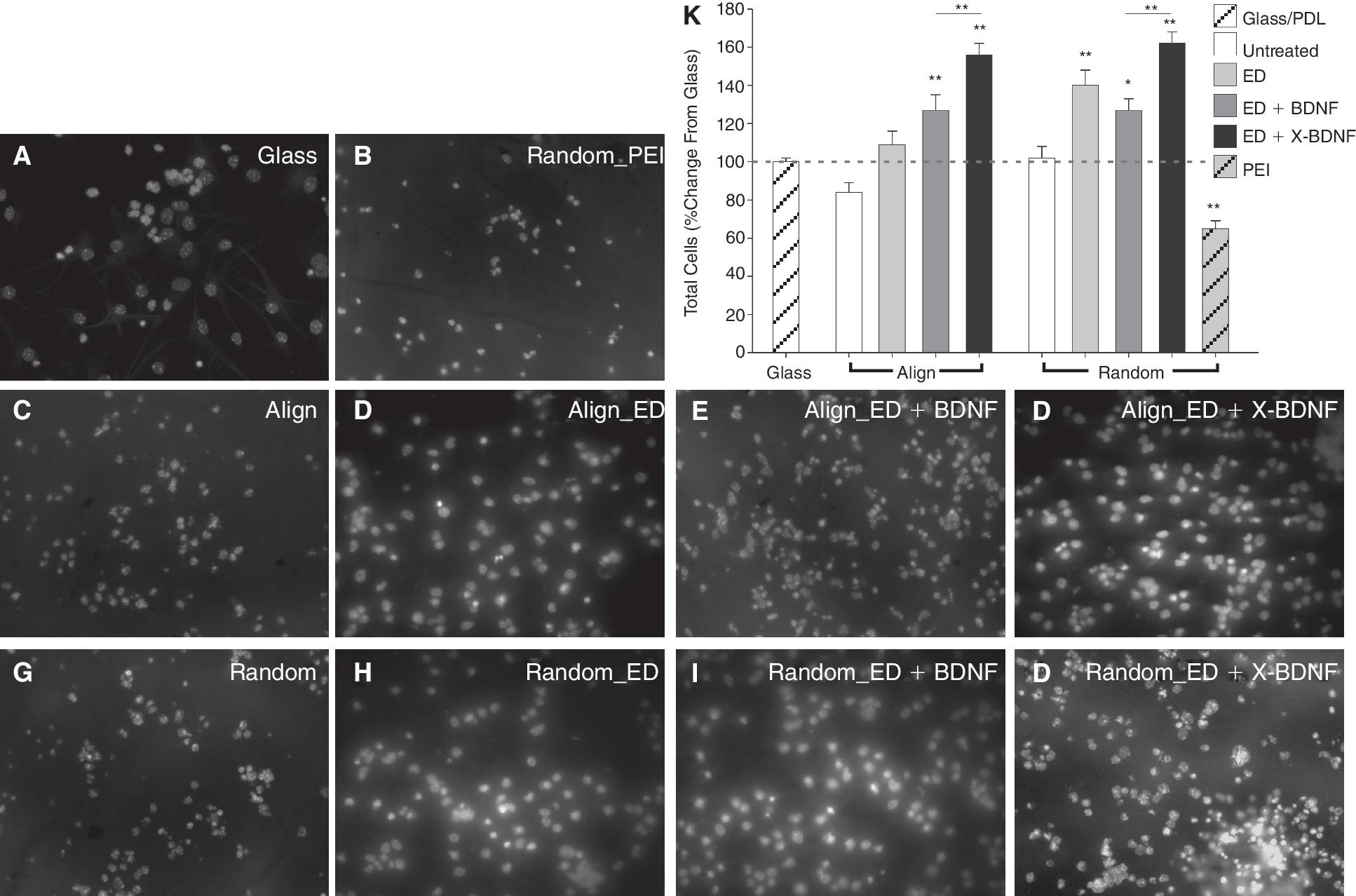
Modified electrospun nanofibers and immobilized brain-derived neurotrophic factor (BDNF) enhance cell number in vitro. (
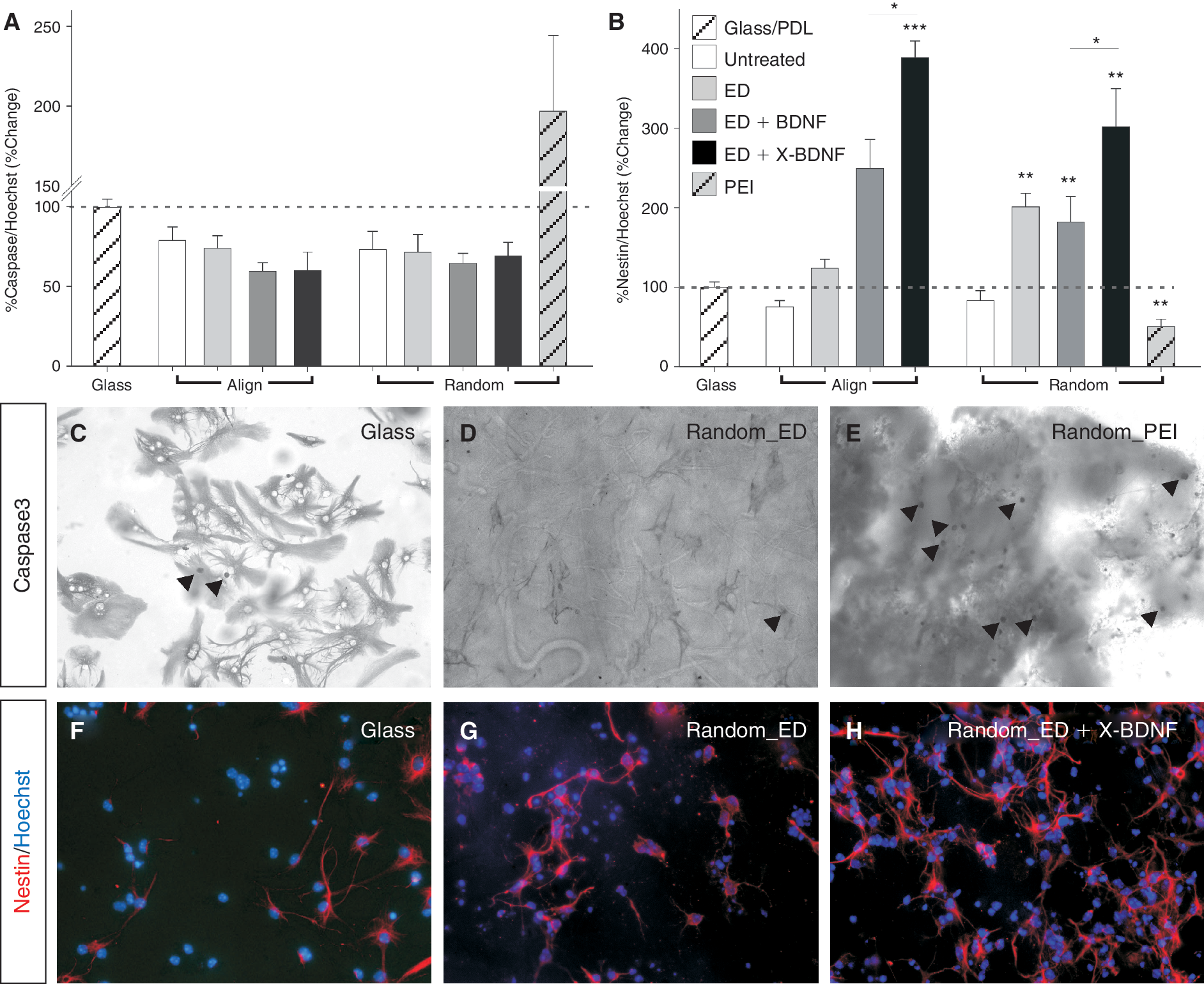
Immobilized brain-derived neurotrophic factor (BDNF) on nanofibers promotes neural stem cell (NSC) proliferation and survival. Quantification of (
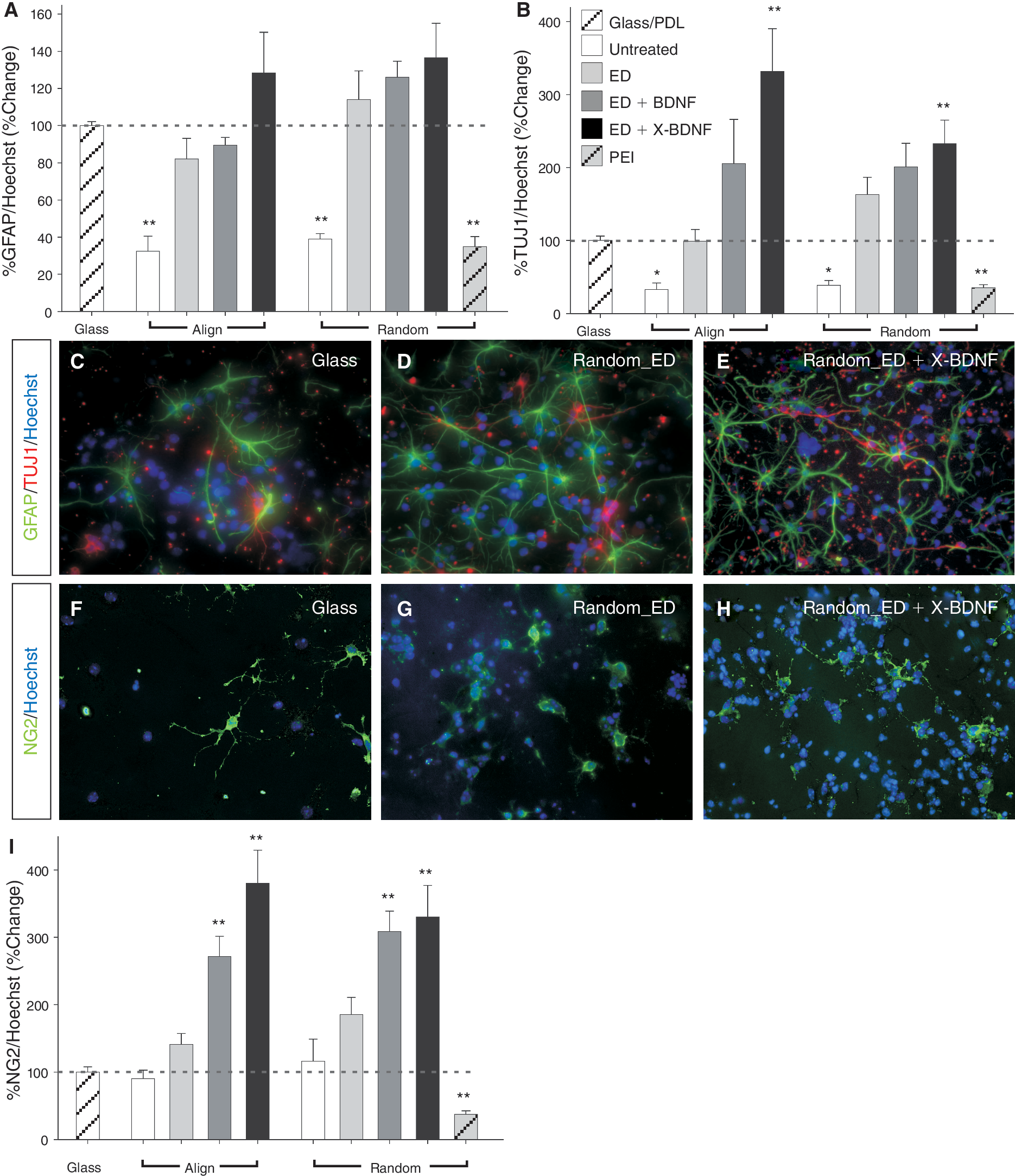
Immobilized brain-derived neurotrophic factor (BDNF) on nanofibers alters trilineage potential of cortical neural stem cell (NSC), promoting oligodendrogenesis. Quantification of (
Results and Discussion
We assessed cortical NSC adhesion, proliferation, and differentiation in order to determine the ability of modified PCL electrospun nanofibers, including ligand immobilization (BDNF), to support cortical NSCs and their derivatives.
Scanning electron microscopy (SEM) examination of the electrospun PCL scaffolds confirmed that randomly orientated and aligned nanofiber architectures were achieved (Fig. 1B and 1C). Henceforth, the scaffold architectures are described as random or aligned. Aminolysation was conducted as previously described [4] and characterized fluorescently using 3-(4-carboxybenzoyl)-quinoline-2-carboxaldehyde. The aminolysation produced 0.1 nmol of NH2/g of PCL polymer. Importantly, aminolysation of the scaffolds allowed sites for subsequent BDNF immobilization. BDNF was bound to both random and aligned PCL scaffolds after aminolysis (ethylene diamine, ED, treatment) as schematically represented in Figure 1D. An ELISA assay using BDNF antibody demonstrated that significantly more BDNF was attached by SMCC-BDNF linkage onto random PCL-ED fibers (0.726 ± 0.15) than to untreated random PCL-ED scaffolds (0.094 ± 0.006), (Fig. 1E). There was also a strong trend for more BDNF to be attached to aligned PCL-ED fibers (P = 0.057) than to untreated aligned PCL-ED scaffolds (0.519 ± 0.14 and 0.142 ± 0.024, respectively, Fig. 1E). Values represent the mean relative amount of bound BDNF ± SEM, Student's t-test, *P < 0.05. Future studies will require that the amount of covalently immobilized ligand on these scaffolds be determined [11,12]; however, the purpose of this study was to first establish whether BDNF could be tethered to these nanofibers, and subsequently, whether immobilized BDNF was superior to soluble BDNF for culturing cortical cells. Our findings establish that BDNF can be successfully tethered to PCL nanofiber scaffolds.
Modified electrospun nanofibers (ED) and immobilized BDNF enhance cell number
Nanofiber scaffolds promote cell adhesion in vitro and in vivo through their high porosity, enhanced surface area-to-volume ratio, and by mimicking components of the extracellular matrix [1, 6, 35, 36]. Hence, we compared cell numbers (total number of Hoechst-labeled nuclei) on a number of modified electrospun PCL nanofiber scaffolds with cell numbers on PDL-coated glass coverslips.
Cultured cells remained predominantly on the surface of the scaffold with few cells penetrating into the scaffold, presumably the result of the small PCL interfiber distances and the large cell size. We observed that 3D nanofibrous scaffolds altered the morphology of the cortical cells, with cells adopting a smaller, rounder morphology compared to the large, flat morphology adopted by cells grown under 2D conditions on plastic culture-ware (data not shown) or glass. Presumably, this change in morphology is due to the altered environment including the 3D exposure of the cells to nutrients and BDNF. A number of neurite fibers infiltrating random scaffolds were also observed, as previously described [1].
Quantitatively, cell numbers remained impartial to culturing on unmodified random or aligned nanofibers (Fig. 2A, 2C, 2G, and 2K); however, aminolysis of the scaffolds (by addition of ED) enhanced cell numbers on aligned fibers scaffolds (9.3% ± 5.0%) with a pronounced effect on random fibers (39.5% ± 9.1%) compared to control cultures (expressed as % change from PDL-coated glass coverslip) (Fig. 2A, 2D, 2H, and 2K).
In comparison, cell adhesion was impaired by attachment of positively charged polyamine chains (PEI) onto scaffolds (35.4% decrease in cell numbers, Fig. 2B and 2K) possibly caused by the high charge density disrupting the cell membrane [37].
Addition of BDNF to the culture media significantly increased cell numbers (random: 26.6% ± 5.4% increase, aligned: 27.8% ± 5.6%, Fig. 2K). More noteworthy, however, were the effects of BDNF immobilization. Anchoring BDNF to PCL-ED scaffolds resulted in significantly greater cell numbers in culture than ED + soluble BDNF (random: 56.4% ± 6.2% and aligned: 64.6% ± 3.7%, Fig. 2E, 2F, and 2I–2K).
We then addressed whether the augmented cell numbers on modified nanofibers were a result of enhanced attachment, cellular proliferation, and/or cell survival. While there was a trend toward increased survival on PCL scaffolds (± modifications), the proportion of caspase-3 immunoreactive cells was not significantly reduced (Fig. 3A). Therefore, in the present study, the increased cell numbers on modified scaffolds is not likely to be due to decreased caspase-3-mediated apoptotic cell death; however, the activation of other death cascades, including autophagy or necrosis, cannot be excluded.
In contrast, there was significantly more cell death on random scaffolds aminolysed with PEI (197.2% ± 47.5%) than on PDL-coated glass (100.0% ± 3.3%, Fig. 3A and 3E), suggesting not only were these scaffolds less amenable to cell adhesion, but more likely reduced cell numbers could be attributed to the toxicity from the highly charged PEI [37].
Although proliferation and self-renewal of mouse embryonic stem cells was enhanced by activation of GTPases when cultured on nanofibers (Rac and PI3K) [38], we could not find an effect on proliferation of NSC grown on untreated nanofibers. However, there was a trend toward greater numbers of nestin+ cells in culture on aligned nanofibers (with ED: 22.5% ± 10.8%) and significantly increased NSC numbers (nestin+ cells) on random_ED fibers (101.75% ± 17.5%) than cultures on their respective untreated scaffolds (Fig. 3B and 3F–3H). There are several plausible explanations for the difference in cell numbers (Fig. 2K) and nestin+ cells (Fig. 3B) on random_ED and aligned_ED scaffolds. Aligned fibers may cause cells to polarize, and subsequently differentiate [2]. This increase in differentiation would result in fewer nestin+ (and consequently total cells). The consequences of cell polarization by aligned fibers is evident by comparing the cell organization in Figure 1F, where nestin+ can be seen aligned with βIII-tubulin+ cells, with the cell organization on random nanofibers (Fig. 1G). In addition, the alignment of PCL nanofibers results in dramatically reduced pore size and interfiber distance, and therefore these scaffolds may in fact better reflect a 2D textured PCL surface (aligned scaffold thickness: 150 ± 50, random scaffold thickness: 250 ± 40). As a result of the reduced available surface area presented to the cells, it is likely that fewer cells adhere. Moreover, in the present study and our previous observation [1], cells cultured on random PCL scaffolds more frequently extended processes (nestin and/or βIII-tubulin) into the scaffold compared to aligned PCL scaffolds, where cell processes predominantly remain on the surface and may thereby influence the ability of cells to adhere to the scaffold.
BDNF promotes proliferation of NSCs [39,40] and in many instances, including in the cortex [23 –26], acts as a pro-survival factor. In our cultures, treatment with soluble BDNF increased the proportion of nestin+ NSCs (align: 149% ± 46.6%, random: 82.1% ± 32.0%), these effects being significantly enhanced by immobilization of the ligand onto both random and aligned nanofibers (289.0% ± 20.6% and 201.8% ± 47.9%, respectively, Fig. 3B), suggesting that the increase in cell numbers observed in Figure 1 may be attributable to proliferation.
Immobilized BDNF on nanofibers alters trilineage potential of cortical NSC
Finally, we examined the effects of nanofiber modification on the differentiation and trilineage potential of cortical NSCs. Cultures were stained for neurons (βIII-tubulin), astrocytes/radial glia (GFAP), and oligodendrocyte progenitors (NG2) and compared to cortical cultures differentiated on PDL-coated glass coverslips. All 3 neural lineages were present under all culturing conditions. Under conventional 2D culturing conditions (on PDL-coated glass coverslips), the proportion of each neural lineage were as follows: %βIII-tubulin/Hoechst = 9.95 ± 2.31, %GFAP/Hoechst = 25.16 ± 6.82, %NG2/Hoechst = 4.29 ± 1.77, mean ± SEM. Note, %nestin+/Hoechst cells = 16.02 ± 2.77.
While astrocytes/radial glia numbers were significantly reduced on unmodified scaffolds, modification of the nanofiber scaffolds offered little benefit to astroglia (Fig. 4A and 4C–4E). Similarly, differentiation of neurons was significantly inhibited by culturing on unmodified nanofibers.
There was a trend toward increased differentiation of neurons cultured in the presence of aminolysis (ED) or ED + soluble BDNF. However, immobilization of BDNF on ED-treated fibers significantly enhanced neuron numbers on random and aligned scaffolds (132.6% ± 32.4% and 231.9% ± 58.3%, respectively, Fig. 4B). These findings are important in the context of tissue repair, implying that immobilizing BDNF on 3D PCL scaffolds can enrich the differentiation of NSCs.
Most noteworthy, with regards to differentiation, were the effects on oligodendrocyte maturation. Consequential benefits from enriching oligodendrocyte numbers may be achieved through aminolysation of fibers and the addition of BDNF, with BDNF immobilized on aligned and random ED fibers increasing oligodendrocyte numbers by 295.2% ± 32.5% and 230.1% ± 47.8%, respectively (Fig. 4I). This data confirm previous findings regarding the ability of BDNF to promote cell survival and proliferation during early stages of oligodendrocyte maturation and reiterate the benefit of ligand immobilization.
Aminolysing nanofiber scaffolds with ED appears to provide multiple benefits including significantly enhancing cell numbers, proliferation, and differentiation. While aminolysis may increase the hydrophilicity of the scaffold, it is also likely that the amine groups present on the surface of the fibers facilitate greater protein adsorption that will subsequently affect the cellular response [4,7].
Past comparisons between ligand immobilization and soluble protein have provided conflicting conclusions. While EGF or transforming growth factor β immobilized onto hydrogels was superior to saturated concentrations of soluble ligand in promoting cell survival [14,41], NGF photochemically bound to hydrogels and soluble NGF resulted in similar proportion of cells extending neurites [19]. It is not clear whether these discrepancies arise from differences between the trophic factors per se or from the bioassay. Regardless, in the present study we attribute at least some of the increase in proliferation and differentiation to BDNF immobilization, given that similar trends (although less significant) were seen in response to soluble BDNF. Further studies, however, will be required to confirm these findings, employing other proteins or peptides of similar molecular size.
Typically BDNF signaling requires BDNF to bind 2 cell surface receptors, TrκB and p75 (or LNGFR, low-affinity nerve growth factor receptor). Hence, when a cell contacts the BDNF immobilized to the scaffold surface, there is likely to be persisting interaction between ligand and receptor, whereas soluble BDNF risks endocytosis and subsequent loss of biological activity [8 –10, 12]. In the context of our results, together with the findings of others [14,41], it remains to be determined whether the differences between soluble and immobilization ligands result from differences between ligands, biomaterial properties (surface tension, chemistry, etc.), or immobilization methods.
In addition to promoting cellular support, immobilization of ligands holds benefits for the future application of these biomaterials in vivo, eliminating the reliance on cannulation/infusion pumps while still maintaining prolonged delivery to the repairing nervous system. In this context, it will be important to determine how long these ligands remain active on the scaffolds.
Conclusion
It is probable and likely that in the future, repair of neural circuits, notably those in the spinal cord, will employ strategies providing both physical and trophic support for new and injured neurons. In addition, it will be of equal importance that neural support cells (such as oligodendrocytes) are also incorporated to ensure close interaction and rapid transmission of electrical activity. In this regard, the structural resemblance of electrospun nanofibers to the extracellular matrix is attracting increasing attention.
Given the 3D organization in vivo setting in which cells normally reside, interact with neighboring cells, and are exposed to various extracellular proteins, these 3D scaffolds provide a more relevant culture assay compared to conventional 2D culture-ware to study cell properties as well as the potential application for these biomaterials in tissue repair. Furthermore, PCL polymers are biodegradable and therefore additionally attractive in the context of tissue repair. Based on the present in vitro findings, incorporating various modifications, we can conclude that these nanofibrous PCL scaffolds are suitable for the support of NSCs and their derivatives. It remains to be determined whether these modified scaffolds behave similarly in vivo, and promote neural repair.
Footnotes
Acknowledgments
This work was supported by grants from the Australian Research Council (DP0985433) and National Health and Medical Research Australia (NHMRC). C.L.P is supported by a National Health and Medical Research Australia (NHMRC) C.J. Martin Fellowship and NHMRC Career Development Award. D.R.N is supported by an ARC Postdoctorate Fellowship.
Author Disclosure Statement
All authors declare no conflict of interest.