
Abstract
In adult cardiomyocytes (CMs), the Na+/Ca2+ exchanger (NCX) is a well-defined determinant of Ca2+ homeostasis. Developmentally, global NCX knockout in mice leads to abnormal myofibrillar organization, electrical defects, and early embryonic death. Little is known about the expression and function of NCX in human heart development. Self-renewable, pluripotent human embryonic stem cells (hESCs) can serve as an excellent experimental model. However, hESC-derived CMs are highly heterogeneous. A stably lentivirus-transduced hESC line (MLC2v-dsRed) was generated to express dsRed under the transcriptional control of the ventricular-restricted myosin light chain-2v (MLC2v) promoter. Electrophysiologically, dsRed+ cells differentiated from MLC2v-dsRed hESCs displayed ventricular action potentials (AP), exclusively. Neither atrial nor pacemaker APs were observed. While ICa-L, If, and IKr were robustly expressed, IKs and IK1 were absent in dsRed+ ventricular hESC-CMs. Upon differentiation (7+40 to +90 days), the basal [Ca2+]i, Ca2+ transient amplitude, maximum upstroke, and decay velocities significantly increased (P < 0.05). The I Ca-L antagonizer nifedipine (1μM) decreased the Ca2+ transient amplitude (to ∼30%) and slowed the kinetics (by ∼2-fold), but Ca2+ transients could still be elicited even after complete I Ca-L blockade, suggesting the presence of additional Ca2+ influx(es). Indeed, Ni2+-sensitive I NCX could be recorded in 7+40− and +90-day dsRed+ hESC-CMs, and its densities increased from −1.2 ± 0.6 pA/pF at −120 mV and 3.6 ± 1.0 pA/pF at 60 mV by 6- and 2-folds, respectively. With higher [Ca2+]i, 7+90-day ventricular hESC-CMs spontaneously but irregularly fired transients upon a single stimulus under an external Na+-free condition; however, without extracellular Na+, nifedipine could completely inhibit Ca2+ transients. We conclude that INCX is functionally expressed in developing ventricular hESC-CMs and contributes to their excitation–contraction coupling.
Introduction
L
Sarcolemmal Na+/Ca2+ exchanger (NCX), a bidirectional transporter that catalyzes the exchange of 3 or 4 Na+ ions for one Ca2+ ion, is a well-defined determinant of Ca2+ homeostasis [4]. NCX is responsible for extruding the elevated Ca2+ during contraction, thereby restoring a low resting [Ca2+]i and a high excitation–contraction (E–C)-coupling gain. As one of the earliest functional gene products, NCX is known to be involved in embryonic heart development and function. Developmentally, global knockout of NCX in transgenic mouse models has been reported to lead to abnormal myofibrillar organization, apoptosis, electrical defects, and early embryonic death [5,6]. However, little is known about the expression and functional profiles of NCX in human heart development. We have previously reported that NCX is expressed at the protein level by western blot analysis in mixed hESC-CMs directly differentiated from hESCs by embryoid body formation [7]. But it remains uncertain whether NCX is functional. As such, their biophysical properties and contribution to E–C coupling are unknown. Since I NCX measurements require the inhibition of most ion channels (such as K+, Ca2+, and Cl− channels) and chamber-specific CMs differ significantly in their ion channel expression profiles, it will be necessary to identify and select the ventricular derivative for experiments to avoid ambiguities. In the present study, we generated an engineered hESC line whose ventricular derivatives were fluorescently labeled, followed by patch-clamp recording of I NCX and functionally assessing its contribution to cytosolic Ca2+ transients and E–C coupling.
Materials and Methods
hESC culturing and differentiation
The H1 (WiCells, Madison, WI) hESC line (NIH code: WA01) chosen for this study were cultured and differentiated as we previously described [2,7]. In brief, H1 cells were grown on irradiated mEFs from 13.5-day embryos of CF-1 mice and trypsin-propagated. The culture medium consisted of 80% DMEM, 20% knockout serum replacement, 4 ng/mL basic fibroblast growth factor (b-FGF), 1 mmol/L glutamine, 0.1 mmol/L β-mercaptoethanol, and 1% nonessential amino acid solution (all from Invitrogen, Carlsbad, CA). For differentiation, hESCs were induced to form embryoid bodies (EBs). Undifferentiated cells were detached using 1 mg/mL type IV collagenase (Gibco-BRL, Carlsbad, CA) and transferred to Petri dishes with differentiation media containing 80% DMED, 20% fetal bovine serum defined (HyClone, Logan, UT), 1 mmol/L glutamine, and 1% nonessential amino acid without b-FGF. The aggregates were cultured in suspension for 7 days, followed by plating on gelatin-coated (0.1%; Sigma-Aldrich, St. Louis, MO) 60-mm dishes to form hESC-CMs.
Lentivirus-mediated gene transfer
For stable genetic modification, lentivirus (LV)-mediated gene transfer was performed as we previously reported [2]. To generate pLV-MLC2v-dsRed, a 250-bp fragment of the MLC2v promoter carrying the homologous sequence also found in rat and human, which has been shown to sufficiently recapitulate the cardiac-specific expression pattern [8 –10], along with a 406-bp region of the human cytomegalovirus (CMV) enhancer and the dsRed gene, were inserted into the 3′ end of the plasmid pLV-EF1α-GFP at the BamHI and ClaI restriction sites. EF1α-GFP was kept for selecting positively transduced undifferentiated hESCs. For generating LV particles, the plasmids pδ8.91, pMD.G, and pLV-MLC2v-dsRed (3:1:2 mass ratio) were co-transfected into HEK293T cells seeded at a density of 6 × 106 cells per 10-cm dish 24 h prior to transfection. LV particles were harvested from the supernatant at 24 and 48 h post-transfection and stored at −80°C before use. For transduction, LV particles with a titer of 106 and an MOI of 3 and polybrene (6 μg/mL) were incubated with cells. A single round of LV transduction typically yields an efficiency of ξ50%.
Isolation of hESC-CMs
For isolating hESC-CMs, dsRed+ outgrowths were microsurgically dissected from EBs at 2 stages, 7+30-to-40 days (40-day) and 7+80-to-90 days (90-day), by a glass knife [7,11] (cf. Fig. 1). The dissected clusters were digested with collagenase II (1 mg/mL) at 37°C for 30 min. The isolated cells were incubated with KB solution containing (mM): 85 KCl, 30 K2HPO4, 5 MgSO4, 1 EGTA, 2 Na2-ATP, 5 pyruvic acid, 5 creatine, 20 taurine, 20
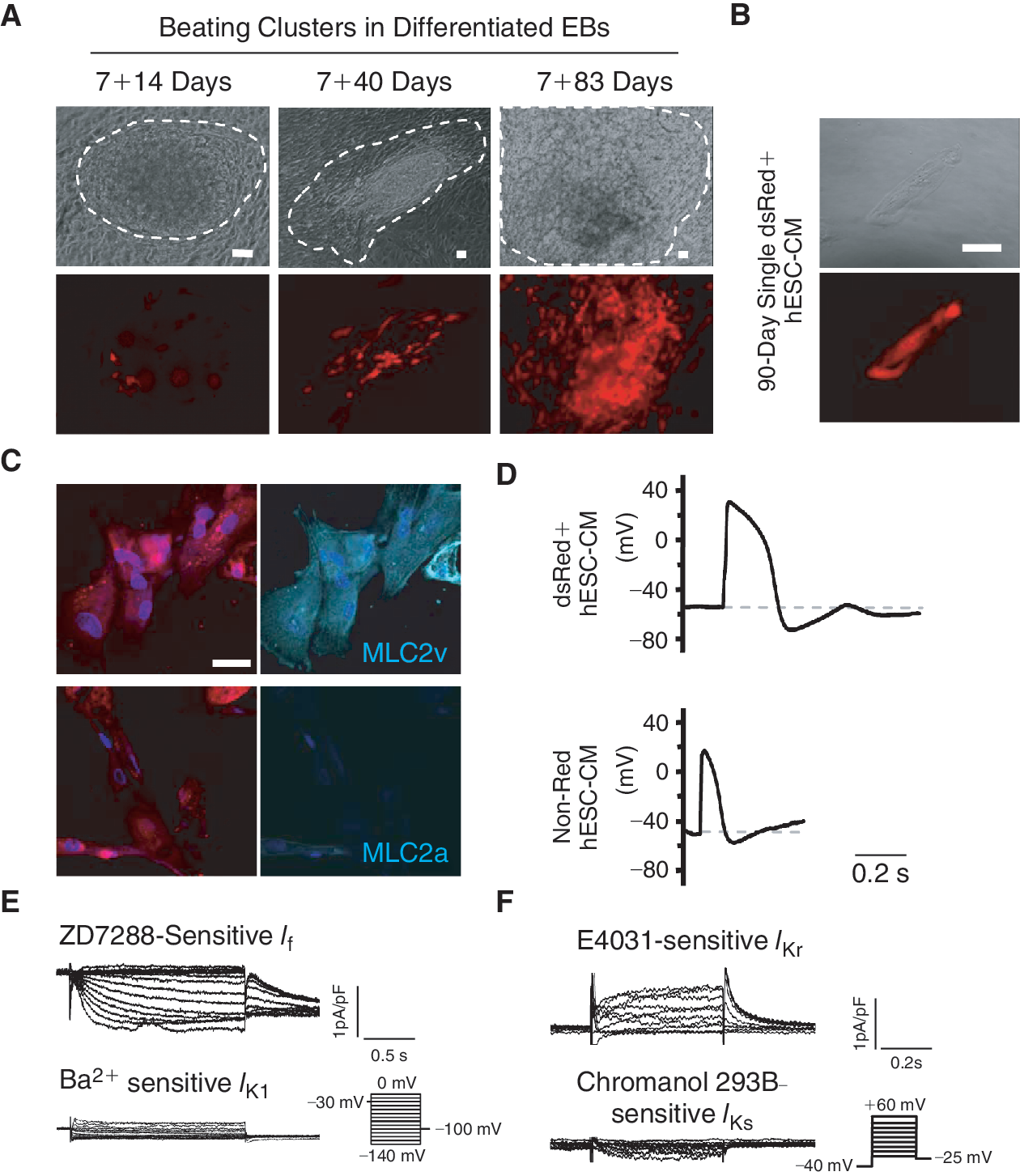
DsRed+ cells differentiated from stably LV-MLC2v-dsRed-transduced human embryonic stem cells (hESCs) displayed a ventricular phenotype. (
Isolation of human fetal ventricular cardiomyocytes (FLV-CMs)
Human fetal ventricular cardiomyocytes (FLV-CMs) were isolated and experimented according to protocols approved by the UC Davis IUPAC and IRB (Protocol #200614787-1). In brief, fetal human hearts (16–18 weeks, Advanced Bioscience Resources, Inc. Alameda, CA) were perfused with enzymatic solutions using a customized Langendorff apparatus as previously described [7]. FLV-CMs were cultured in laminin-coated 24-well dishes with a density of ∼5 × 105 cells/well with media containing: 5 mM carnitine, 5 mM creatine, 5 mM taurine, 100 μg/mL penicillin–streptomycin, and 10% fetal bovine serum in Medium 199 (Sigma-Aldrich Corp., St. Louis, MO).
Immunostaining
Cells were fixed for 15 min at room temperature with 4% paraformaldehyde in PBS. After washing with PBS, cells were permeabilized in PBS containing 0.2% Triton X-100, then incubated with primary mouse anti-MLC2v or anti-MLC2a monoclonal antibody. Alexa Fluor-647 anti-mouse IgG (Invitrogen) was the second antibody used for fluorescence imaging. Hoechst 33342 (H3570; Invitrogen) was used to stain the nuclei. Coverslips were mounted onto glass slides in Prolong Gold antifade reagent (Invitrogen). Samples were imaged on a confocal laser scanning microscope (C1si, Nikon, Japan).
Electrophysiology
Electrophysiological experiments were performed using the whole-cell patch-clamp technique with an Axopatch 200B amplifier and the pClamp9.2 software (Axon Instruments Inc., Foster City, CA) as previously described [3]. A xenon arc lamp was used to view the dsRed fluorescence at 560/590 nm (excitation/emission). Patch pipettes were prepared from 1.5 mm thin-walled borosilicate glass tubes using a Sutter micropipette puller P-97 and had typical resistances of 4–6 Mω The micropipettes were filled with an internal solution containing (mmol/L): 110 K+ aspartate, 20 KCl, 1 MgCl2, 0.1 NaGTP, 5 MgATP, 5 Na2-phospocreatine, 1 EGTA, 10 HEPES, pH adjusted to 7.3 with KOH. The external Tyrode's bath solution consisted of (mmol/L): 140 NaCl, 5 KCl, 1 CaCl2, 1 MgCl2, 10 glucose, 10 HEPES, pH adjusted to 7.4 with NaOH. For AP recording, current-clamp recordings were performed at 37°C within 24 to 48 h after hESC-CM isolation. The hESC-CMs were given a stimulus of 0.1–0.5 nA for 1–5 ms to elicit AP. Voltage-clamp recordings of ionic currents were performed using standard electrophysiological and pharmacological isolation.
For measuring the L-type Ca2+ currents (I Ca-L), the internal solution contained (in mM): 130 CsCl, 1.0 MgCl2, 0.1 NaGTP, 5 MgATP, 10 HEPES, and 10 EGTA. pH was adjusted to 7.2 with CsOH. The external solution contained (in mM): 136 NaCl, 5.4 CsCl, 1.0 MgCl2, 2.0 CaCl2, 2.0 NaH2PO4, 10 glucose, and 10 HEPES. pH was adjusted to 7.4 with NaOH, TTX (0.05 mM) and TEA-Cl (20 mM) were added during the recording, and I Ca-L was elicited by stepping to various voltages (−50 to+10 mV, 10 mV increments, 0.2 Hz) for 100 ms from a holding potential of −30 mV, and defined as 5 mM nifedipine-sensitive currents. Steady-state inactivation was determined by stepping to various pre-pulse voltages (−50 to +10 mV, 10 mV increments) for 1 s prior to depolarization to a fixed 250-ms test pulse of +10 mV every 10 s. Peak currents obtained at all voltages were normalized to the maximal value.
NCX current (I NCX) density was determined using the whole-cell patch-clamp technique as described previously [12]. [Ca2+]i was buffered to 150 nM with BAPTA (calculated using the Maxchelator program [13]). The external solution was K+-free and contained (in mmol/L): Na+ 135, Ca2+ 2, MgCl 1, glucose 10, HEPES 10, CsCl 10 (to block the inward rectifier K+ current, I K1, and the Na+/K+ pump), (in μmol/L): niflumic acid 100 (to block Ca2+-activated Cl− current), ouabain 10 (Na+/K+ pump inhibitor), and verapamil 10 (dihydropyridine antagonist), adjusted to pH 7.4 (CsOH). The internal solution contained (in mmol/L): CsCl 136, NaCl 10, aspartic acid 42, MgCl2 3, HEPES 5, tetraethylammonium (TEA) 20, MgATP 10, and 150 mM free [Ca2+]i, adjusted to pH 7.4 (CsOH). The holding potential was −30 mV to inactivate the T-type Ca2+ and Na+ channels. Slow-ramp pulses were applied (+60 to −120 mV, 0.09 V/s) at 10 s intervals to construct the current–voltage (I/V) relationships. I NCX was measured as the bidirectional Ni2+ (5 mM)-sensitive current.
Measurements of cytosolic Ca2+
Intracellular Ca2+ transients were recorded from dsRed+ hESC-CMs as identified by fluorescence microscope, within 48 h after plating. A spectrofluorometric method with Fura-2/AM as the Ca2+ indicator was used for measuring [Ca2+]i. dsRed+ hESC-CMs were incubated with 5 μM Fura-2/AM and 0.2% pluronic F-127 for 30 min at 37°C. Fluorescent signals obtained upon excitation at 340 nm (F340) and 380 nm (F380) were recorded from cells perfusing with Tyrode solution unless otherwise indicated. For Na+-free solution, LiCl (140 mM) was used to replace NaCl (140 mM). The F340/F380 ratio was used to represent cytosolic [Ca2+]i. To elicit cytoplasmic Ca2+ transients, hESC-CMs were electrically pulsed (0.1–0.5 Hz). Ca2+ transients were recorded and analyzed after a series of depolarization that enabled each transient to fully decay so as to establish a steady state. Data were analyzed using the Ionwizard software (Version 5, IonOptix) to generate the Ca2+ transient parameters reported [7,11,14].
Statistical analysis
All data were expressed as means ± SEM. One-way ANOVA followed by Newman-Keuls multiple comparison tests or paired t-test was carried out to test for differences between the mean values within the same study. A difference of P < 0.05 was considered significant.
Results
dsRed+ cells differentiated from stably LV-MLC2v-dsRed-transduced hESCs displayed a ventricular phenotype
To confirm the efficacy of LV-MLC2v-dsRed for identifying ventricular CMs, we first transduced and compared human fetal left ventricular (FLV) CMs and human embryonic kidney (HEK) 293 cells. FLV-CMs (but not HEK293 cells) uniquely expressed dsRed after LV-MLC2v-dsRed transduction (Supplementary Fig. 1A; Supplementary materials are available online at
For functional characterization, we dissected from EBs beating clusters that contained dsRed+ cells followed by dissociating them into single cells for electrophysiological recording. After FACS sorting, a typical single 90-day dsRed+ cell after attachment is shown in Figure 1B. As anticipated, dsRed+ cells were stained positively for the ventricular isoform (MLC2v), but not the atrial isoform (MLC2a), of the myosin light chain (Fig. 1C). Figure 1D further shows that functionally only ventricular-like action potentials (AP) could be recorded from 40-day dsRed+ cells (n = 15 of 15). In contrast, atrial-like APs were observed only in non-dsRed cells (5 of 7) isolated from the same beating clusters. The hyperpolarization-activated pacemaker current (I f) and the rapid component of the delayed rectifier (I Kr) were expressed in dsRed+ hESC-CMs; but the slow component (I Ks) and the inward rectifier (I K1) were not (Fig. 1E and 1F). These results were consistent with wild-type hESC-CMs that displayed ventricular APs. Therefore, dsRed+ cells differentiated from MLC2v-dsRed hESCs belonged to the ventricular lineage. Since cardiac differentiation of hESCs is known to generate a heterogeneous population of ventricular, atrial, and pacemaker derivatives [1,3], selection of dsRed+ hESC-CMs for experiments will enable us to study the ventricular lineage without ambiguities.
Development of Ca2+ transients during ventricular hESC-CM differentiation
Ca2+ homeostasis is crucial for E–C coupling and subsequently, the contractile properties of functioning CMs. Figure 2 summarizes the Ca2+ transient properties recorded from dsRed+ (ventricular) hESC-CMs at different time points upon differentiation. From 7+40 to 7+90-day post-differentiation, both the basal [Ca2+]i and Ca2+ transient amplitude significantly increased (P < 0.05) (Fig. 2A–2C). Ca2+ transients of the 7+90-day group also had faster kinetics with higher maximum upstroke (V max-upstroke; Fig. 2D) and maximum decay (V max-decay; Fig. 2E) velocities (P < 0.05). Taken together, these results demonstrate that Ca2+ transients of ventricular hESC-CMs appeared to time-dependently mature in culture, although the adult level was still not reached even after almost 100 days.
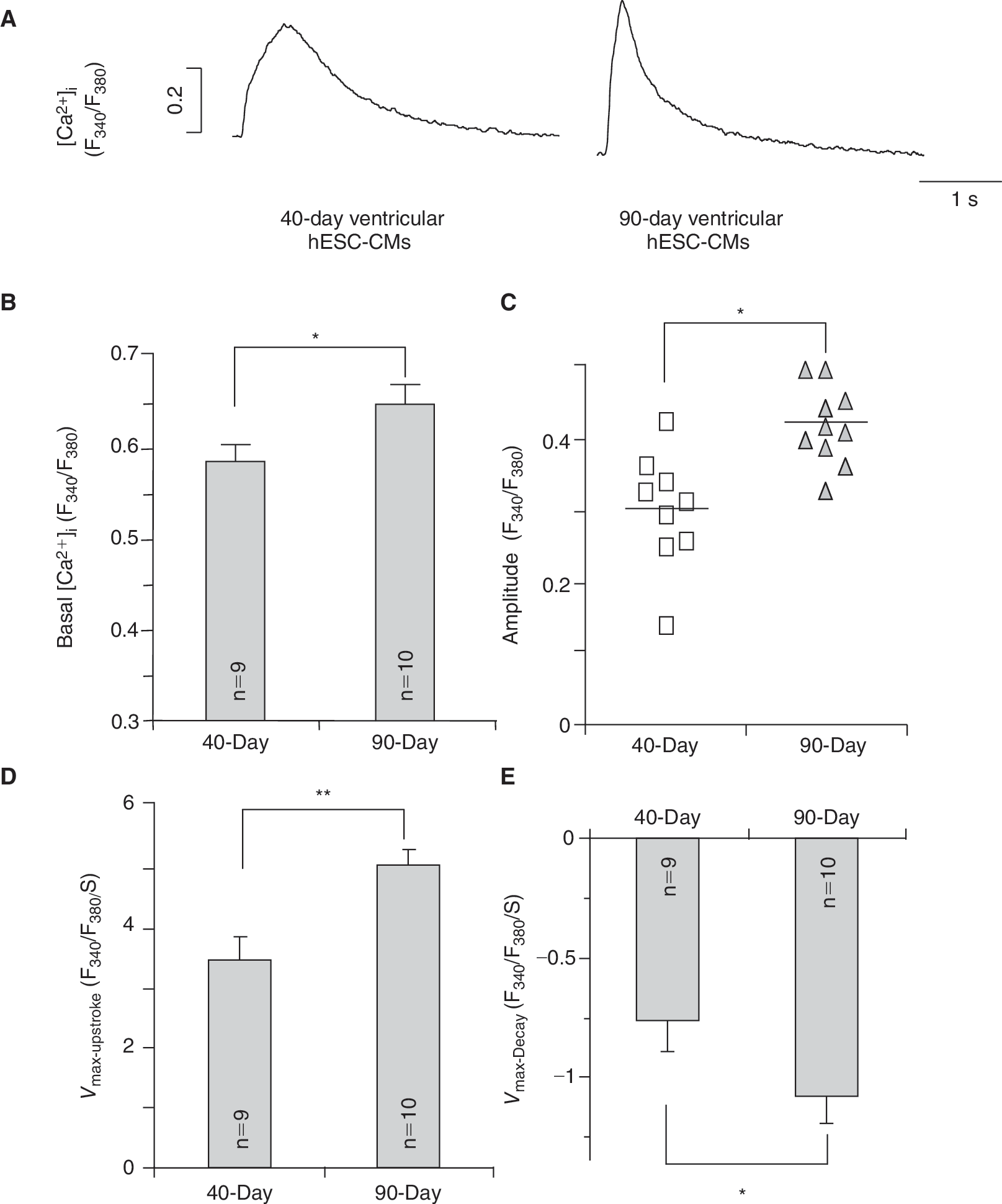
Ca2+ transients recorded from 7+40- to 7+90-day dsRed+ ventricular hESC-CMs. (
Contribution of L-type Ca2+ channels to Ca2+ transients
In adult CMs, L-type Ca2+ (I Ca-L) channels mediate the transport of trans-sarcolemmal Ca2+ influx for activating Ca2+ release from the sarcoplasmic reticulum (SR) via a process known as Ca2+-induced Ca2+ release (CICR) during E–C coupling. In murine ESC-CMs, I Ca-L plays a similar functional role but the same in ventricular hESC-CMs has not been defined. Figure 3A and 3B shows that I Ca-L was robustly expressed in 7+40-day dsRed+ ventricular hESC-CMs (2.24 ± 0.54 pA/pF at +10 mV; n = 4). The steady-state activation (V 1/2 = −10.9 ± 0.6 mV, K = 4.6 ± 0.3) and inactivation (V 1/2 = −27.1 ± 2.0 mV, K = 7.6 ± 1.3) curves are given in Supplementary Figure 2. Upon infusion of nifedipine (1 μmol/L), a selective antagonist of I Ca-L, the Ca2+ transient amplitude decreased to 33.5% ± 9.2% in 40-day (n = 4) and 28.0% ± 4.6% in 90-day (n = 4) dsRed+ ventricular hESC-CMs (Fig. 3C and 3D). The upstroke velocities of Ca2+ transients were significantly slowed (Fig. 3E). However, the inhibitory effect of nifedipine were not significantly different between 7+40-day and 7+90-day dsRed+ ventricular hESC-CMs (P ξ 0.05). Interestingly, Ca2+ transients, albeit small in the amplitude (Fig. 3C), could still elicited in both 7+40- and 7+90-day ventricular hESC-CMs even after I Ca-L was completely blocked by 10 μmol/L nifedipine (Fig. 3B). These results raise the possibility that additional source(s) of Ca2+ other than I Ca-L is present in ventricular hESC-CMs that suffice to initiate CICR.
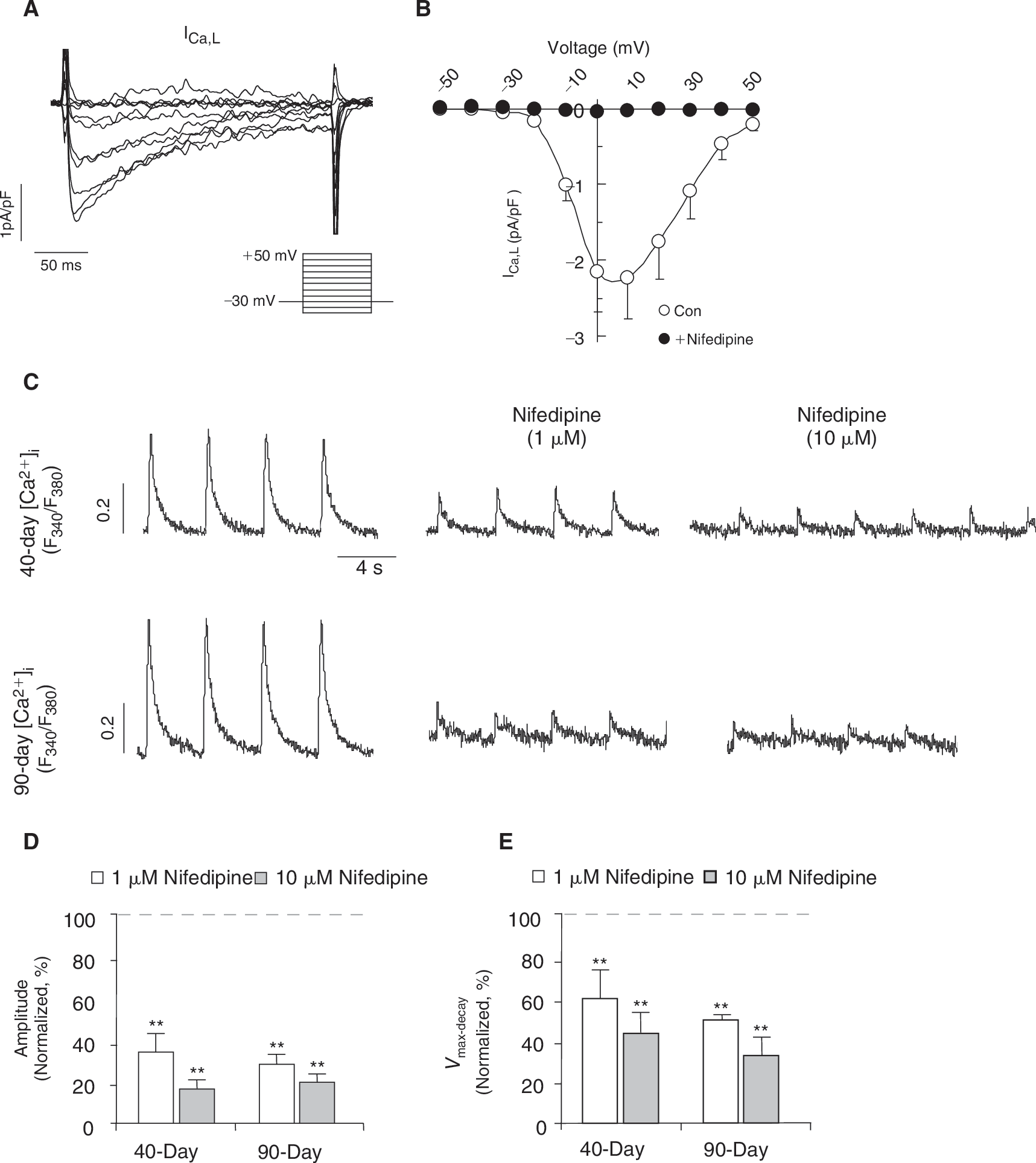
The inhibitory effect of nifedipine on Ca2+ transients during ventricular hESC-CM differentiation. (
Contribution of NCX to Ca2+ transients during human ventricular differentiation
To obtain molecular insights, we next analyzed the transcriptional expression levels of the three NCX isoforms (ie, NCX1, 2, and 3) in undifferentiated hESCs, hESC-derived ventricular (V) CMs, human fetal VCMs, and adult VCMs. Figure 4A shows that only the expression of NCX1, but not the other isoforms, increased during cardiac differentiation from hESCs to hESC-VCMs, peaked in fetal VCMs then decreased in adult VCMs. In our previous studies, we have also reported that the NCX1 is abundantly expressed in unsorted human ESC-CMs at the protein level [7]. However, it remains unknown whether I NCX is functionally expressed in ventricular hESC-CMs. When operating in the reverse mode, the NCX current (I NCX) can contribute to the elevation of cytosolic Ca2+ in early developing murine ESC-CMs [14] and fetal CMs [15]. Figure 4B shows that Ni2+ (5 mM), an inhibitor of NCX, completely abolished Ca2+ transients of dsRed+ ventricular hESC-CMs. To test for the functional expression of I NCX, we voltage-clamped and compared 7+40- and 7+90-day dsRed+ ventricular hESC-CMs. Similar to previous studies, blockers for K+, Ca2+-activated Cl−, and L-type Ca2+ currents, as well as the Na+/K+ pump, were added to isolate I NCX [12]. Figure 4C shows that I NCX, defined as Ni 2 + (5 mM)-sensitive currents, was indeed functionally and robustly expressed in 7+40- and 7+90-day dsRed+ ventricular hESC-CMs. A voltage ramp from +60 to −120 mV elicited an almost linear current-voltage relationship of I NCX (Fig. 4D), similar to those previously described for fetal and adult mouse CMs [15]. The I NCX densities in 40-day ventricular hESC-CMs were −1.2 ± 0.6 pA/pF (n = 4) at −120 mV (Ca2+ outward mode) and 3.6 ± 1.0 pA/pF at 60 mV (Ca2+ inward mode). Significant increases of I NCX, particularly in the Ca2+ outward mode, were observed in 90-day ventricular hESC-CMs (−6.9 ± 1.3 pA/pF or ∼6-fold increase at −120 mV n = 5 and 7.9 ± 1.3 pA/pF or ∼2-fold at 60 mV; P < 0.05).
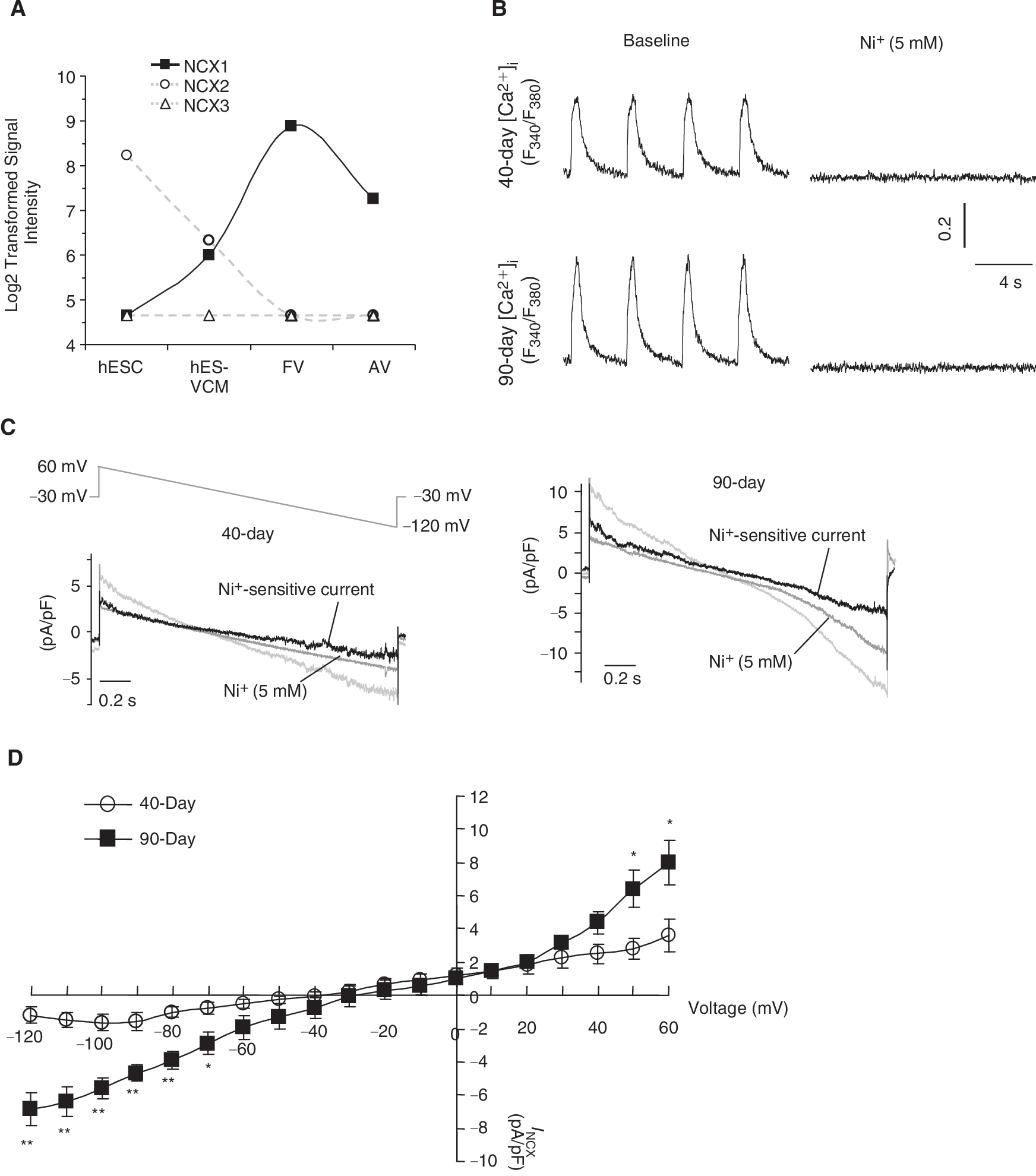
Developmental changes of NCX current (I
NCX) in dsRed+ ventricular hESC-CMs. (
To explore the functional role of I NCX in Ca2+ transients of ventricular hESC-CMs, we next applied a Na+-free external solution to prevent NCX from extruding Ca2+ [14,16]. Upon its perfusion, the basal [Ca2+]i increased significantly in 7+90-day, but not 7+40-day, dsRed+ ventricular hESC-CMs (Fig. 5A and 5B). With a higher cytosolic [Ca2+]i, 7+90-day ventricular hESC-CMs spontaneously but irregularly fired Ca2+ transients upon a single stimulus (Fig. 5A). Detailed analysis of the first Ca2+ transient elicited revealed that both its Ca2+ transient amplitude and decay velocity reduced to 76.8% ± 6.2% (n = 4, Fig. 5C) and 38.0% ± 13.9% (n = 4, Fig. 5D), respectively. Similarly, the Na+-free solution decreased the amplitude (to 66.3% ± 5.8%, n = 4) and slowed the decay velocity (to 51.6% ± 16.4%, n = 4) of 7+40-day ventricular hESC-CMs, although irregular firing was not observed. Consistently, Ca2+ transients were completely blocked in both 7+40- and 7+90-day dsRed+ ventricular hESC-CMs cells only when both nifedipine and Na+-free conditions were used. Upon reperfusion of the normal Tyrode solution (containing 140 mM Na+) even in the presence of nifedipine (10 μmol/L), small Ca2+ transients could be elicited (Fig. 5E). Taken collectively, our results strongly suggest that the reverse mode of I NCX contributes significantly to the Ca2+- handling properties of developing ventricular hESC-CMs.
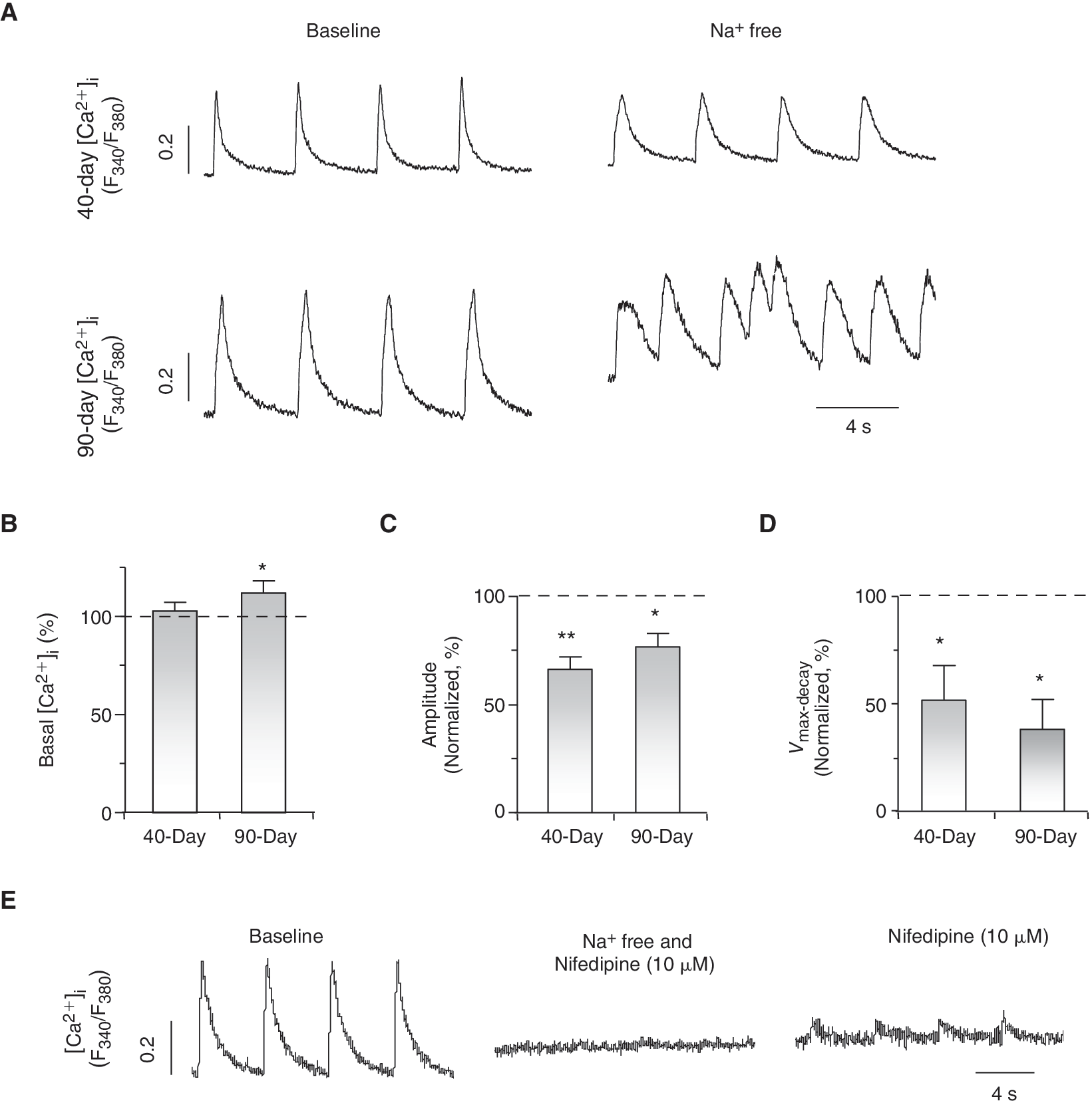
I
NCX contributes to E-C coupling of ventricular hESC-CMs. (
Discussion
It is now well-accepted that hESC-CMs differentiated from hESCs are highly heterogeneous consisting of ventricular-, atrial-, and pacemaker-like derivatives, fibroblasts, as well as extracardiac cell types. Technically, this heterogeneity has largely limited their functional characterization and therefore, our understanding of their biology. Adding an extra level of complexity is the fact that although hESCs are by definition pluripotent, different lines have distinct cardiogenic potentials to become early ventricular-, atrial-, and pacemaker-like derivatives [3]. For instance, we have reported that HES2 cells have a high likelihood of differentiating into ventricular-like hESC-CMs while the atrial-like phenotype predominates in H1. Pacemaker-like hESCs were always the minority (<5%) [3]. Despite the different distributions, HES2-derived ventricular-, atrial-, and pacemaker-like derivatives display electrophysiological properties comparable to those of H1 once they are differentiated. As such, ambiguities exist if chamber-specific cells are not selected for experiments. In the present study, we employed the classical strategy that has been proven for mESCs [9,17] for purifying out ventricular hESC-CMs for experiments. Expression of dsRed under the transcriptional control of MLC2v appeared upon the occurrence of spontaneously beating activities and increased time-dependently; dsRed+ cells could be isolated and FACS-sorted into single cells. Patch-clamp recordings revealed an AP profile that is consistent with the ventricular phenotype. dsRed+ hESC-CMs were positively immunostained for the cardiac-specific proteins cTnI and MHC (data not shown). Indeed, Huber et al. recently reported a similar strategy to identify hESC-derived ventricular derivatives by expressing GFP under the transcriptional control of the MLC2v promoter although the functional properties of their labeled derivatives were not studied and reported [18].
Compared to adult CMs, fetal CMs have smaller Ca2+ transient amplitudes and slower dynamics, and display developmental changes during the maturation process [11]. In rat and rabbit, cardiac sarcolemmal NCX is abundantly expressed and functionally well-developed in late fetal, reaches a maximum at perinatal, followed by declining at adult stages [19,20]. In developing mouse embryonic CMs, functional NCX operates in both Ca2+ inward and outward modes to maintain intracellular Ca2+ homeostasis also with a significantly higher basal NCX current (I NCX) density at early (10.5 dpc) than late (16.5 dpc) stages [15]. Haddock et al. report that Ca2+ entry via outward I NCX directly supports contraction of newborn rabbit ventricular myocytes [21]. Along the same line, both NCX transcript and protein levels have been recently shown to increase from 10-week to 20-week gestation during human heart development, then decrease at neonate and further diminish at adult stages [22]. Consistently, the same expression pattern was observed in our own study. We have previously reported that the cardiac NCX1 protein is abundantly expressed in human fetal ventricular and hESC-CMs [7]. Using MLC2v-dsRed hESCs, here we further showed that NCX is indeed functionally expressed in ventricular hESC-CMs. Developmentally, I NCX, particularly the outward mode, increases time-dependently, consistent with the higher NCX transcript and protein expression levels in human fetal heart [7,22]. Given the relatively low NCX level in adult, it is reasonable to extrapolate from these findings that NCX continues to increase during cardiogenesis and declines only after the fetal stage to reach the adult level as part of the maturation process. Taken collectively, our results and those of others have revealed significant evolutionary footprints, as well as differences, between human and various mammalian species.
In adult CMs, E-C coupling is initiated primarily by CICR. ICa-
Pluripotent hESC-like iPSCs have been recently derived by reprogramming adult somatic (fibroblast) cells [32,33]. Using protocols originally developed for maintaining and cardiac differentiation of hESCs, iPSC can be similarly cultured and differentiated into CMs. iPSC-CMs express key cardiac-specific proteins, and spontaneously contract in a manner similar to hESC-CMs, and are also heterogeneous with nodal-, atrial-, and ventricular-like phenotypes [34]. Our MLC2v-dsRed-based selection strategy will be useful for characterizing and comparing ventricular iPSC- and hESC-CMs. A better understanding of the basic biology of ventricular derivatives is crucial for their ultimate clinical application in myocardial repair. Additionally, although the Ca2+-handling properties of ventricular hESC-CMs appear to mature in culture in vitro, they did not reach the adult level even after 100 days. The results further implicate that alternative protocols need to be developed for facilitating functional maturation to improve both the safety and efficacy of hESC-CMs.
Conclusion
We have generated the stably LV-transduced MLC2v-dsRed hESC line and electrophysiologically characterized their dsRed+ ventricular derivatives. Based on our present results, we conclude that NCX is functionally expressed in, and contributes to their Ca2+-handling properties of ventricular hESC-CMs.
Footnotes
Acknowledgments
This work was supported by grants from the National Institutes of Health—R01 HL72857 (to R.A.L.), the Stem Cell Program of the University of California (to R.A.L.), the California Institute for Regenerative Medicine (to J.D.F. and R.A.L.), and the CC Wong Foundation Stem Cell Fund (to R.A.L.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.