
Abstract
Resveratrol is a natural polyphenol antioxidant that has been shown to facilitate osteogenic differentiation. A recent breakthrough has demonstrated that ectopic expression of four genes is sufficient to reprogram murine and human fibroblasts into induced pluripotent stem (iPS) cells. However, the roles of resveratrol in the differentiation and cytoprotection of iPS cells have never been studied. In this study, we showed that, in addition to cardiac cells, neuron-like cells, and adipocytes, mouse iPS cells could differentiate into osteocyte-like cells. Using atomic force microscopy that provided nanoscale resolution, we monitored mechanical properties of living iPS cells during osteogenic differentiation. The intensity of mineralization and stiffness in differentiating iPS significantly increased after 14 days of osteogenic induction. Furthermore, resveratrol was found to facilitate osteogenic differentiation in both iPS and embryonic stem cells, as shown by increased mineralization, up-regulation of osteogenic markers, and decreased elastic modulus. Dexamethasone-induced apoptosis in iPS cell-derived osteocyte-like cells was effectively prevented by pretreatment with resveratrol. Furthermore, resveratrol significantly increased manganese superoxide dismutase expression and intracellular glutathione level, thereby efficiently decreasing dexamethasone-induced reactive oxygen species (ROS) production and cytotoxicity. Transplantation experiments using iPS cell-derived osteocyte-like cells further demonstrated that oral intake of resveratrol could up-regulate osteopontin expression and inhibit teratoma formation in vivo. In sum, resveratrol can facilitate differentiation of iPS cells into osteocyte-like cells, protect these iPS cell-derived osteocyte-like cells from glucocorticoid-induced oxidative damage, and decrease tumorigenicity of iPS cells. These findings implicate roles of resveratrol and iPS cells in the stem cell therapy of orthopedic diseases.
Introduction
A
Resveratrol (trans-3,5,4′-trihydroxystilbene), a natural polyphenol found largely in the skins of red grapes, nuts, pomegranates, and red wine [8], has several health-promoting effects, including cardiac and neural protection, anti-inflammatory and anti-aging activities, and lifespan extension [8 –10]. Recent studies have demonstrated that resveratrol could enhance osteoblast differentiation [11]. However, the role of resveratrol in promoting osteogenic differentiation in iPS cells needs to be determined. Moreover, resveratrol possesses the ability to scavenge oxidatively generated free radicals, and can either protect cells from apoptosis or induce apoptosis [12 –14]. The exact roles of resveratrol in the survival of iPS cell-derived osteocyte-like cells remain to be clarified.
Atomic force microscopy (AFM) has extremely high-resolution topography and is a powerful tool for the precise measurement of mechanical properties of materials, including cells [15]. Over the past 2 decades, AFM has been widely applied to biological samples [16]. Moreover, AFM provides dramatic improvements over other techniques for measuring the physical properties of biological systems in their natural conditions, making it useful for researching protein or DNA structures, molecular interactions, and biomechanical characteristics [17]. The mechanical elasticity of materials, generally measured as stiffness, is one of the most essential parameters of almost every material and can be determined at the nanoscale by AFM indentation methods [18,19]. A recent study has suggested that AFM can be used to acquire detailed surface images of living cells and materials with nanoscale resolution [15].
In this study, we successfully induced murine iPS cells to differentiate into ectodermal and mesodermal lineage cells, including osteocyte-like cells. The AFM was used to acquire surface images of living iPS cells at nanoscale resolution, as well as to measure the mechanical properties of these cells throughout osteogenic differentiation. The effects of resveratrol on osteogenic differentiation in iPS cells were then determined, followed by examination of the cytoprotective roles of resveratrol in these iPS cell-derived osteocyte-like cells.
Materials and Methods
Culture of iPS and ES cells
Murine germ line-competent iPS cells were kindly provided by Kyoto University (Dr. Shinya Yamanaka) and RIKEN BRC, Japan [6]. The iPS cells were cultured as previously described [5,6]. In brief, iPS cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen, Carlsbad, CA) containing 15% fetal bovine serum (FBS; HyClone, Logan, UT, USA), 1% nonessential amino acids, 1 mmol/L sodium pyruvate, 5.5 mmol/L β-mercaptoethanol, 50 U/mL penicillin, and 50 mg/mL streptomycin (all from Invitrogen) on feeder layers of mouse embryonic fibroblasts that had been treated with 10 µg/mL mitomycin C for 3 h. All experiments were performed with the use of iPS cell line with Nanog promoter-driven green fluorescent protein (GFP). The murine ES cell line derived from BALB/c mouse blastocyst (ESC26GJ constructed by Lee and others at Animal Technology Institute Taiwan) was used in the present study. This line had been transfected with pCX-EGFP to express green fluorescence constitutively [20]. Undifferentiated ES cells were cultured on mitomycin C-treated (10 µg/mL, for 3 h) STO (mouse embryonic fibroblast cell line, ATCC CRL-1503) in DMEM supplemented with 15% FBS, 1% nonessential amino acid, 0.1 mmol/L β-mercaptoethanol, 100 U/mL penicillin, 100 µg/mL streptomycin, and 100 U/mL leukemia inhibitory factor (R&D Systems, Minneapolis, MN, USA).
Osteogenic differentiation
For osteogenic induction, ES or iPS cells were cultured in DMEM-LG (Invitrogen) supplemented with 15% FBS, 50 µg/mL ascorbate-2-phosphate, 10 nmol/L dexamethasone, and 10 mmol/L β-glycerophosphate (Sigma, St. Louis, MO) for 2 weeks. At the end of osteogenic induction, cells were washed twice with PBS, fixed for 10 min at room temperature with 3.7% paraformaldehyde, and stained with von Kossa stain and Alizarin red to assess osteogenic differentiation [3,4].
Real-time RT-PCR
Real-time RT-PCR was performed as previously described [21]. In brief, total RNA (1 µg) of each sample was reverse-transcribed using 0.5 µg oligo dT and 200 U Superscript II RT (Invitrogen). The primer sequences for real-time RT-PCR were listed in Table 1. The amplification was carried out in a total volume of 20 µL containing 0.5 µmol/L of each primer, 4 mmol/L MgCl2, 2 µL LightCycler™–FastStart DNA Master SYBR green I (Roche Molecular Systems, Alameda, CA), and 2 µL of 1:10 diluted cDNA. PCRs were prepared in duplicate and performed using the following program: 95°C for 10 min, followed by 40 cycles of denaturation at 95°C for 10 s, annealing at 55°C for 5 s, and extension at 72°C for 20 s. Standard curves (cycle threshold values versus template concentration) were prepared for each target gene and for the endogenous reference gene (GAPDH) for each sample. Quantification of unknown samples was performed using LightCycler Relative Quantification Software version 3.3 (Roche).
T
Measurement of cell hardness and elasticity by AFM
For AFM studies, iPS or ES cells were cultured in 6-cm culture dishes. At indicated intervals of osteogenic induction, cell morphology was examined using AFM adapted to biological research (Solver BIO, NT-MDT Co., Moscow, Russia) in combination with an inverted microscope (IX70, Olympus, Japan). An active antivibration system (TS-140, HWL Scientific Inst. GmbH, Ammerbuch, Germany) was used, along with a triangular silicon probe with spring constant of 0.1 N/m (CSG 11 type B, NT-MDT Co., Moscow, Russia) in semi-contact mode (resonance = 20 kHz) under a liquid environment. The relationship between force (F) and cantilever deflection (d c) generally follows the function: F = −k × d c (Hooke’s Law, k = spring constant of the cantilever). Therefore, d c is correlated with F. Hardness of the cells was measured using the same type of tip in contact mode to acquire the force–distance curve. The tip-sample distance is equivalent to the displacement in the z-piezo position. The absolute value of the slope (−nA/nm) in this curve is known to indicate the different levels of force (nA) applied over the same distance (approaching the cell), and thus can be understood as a qualitative measure of stiffness and elasticity.
Cell survival analysis by MTT assay and detection of apoptotic cells with annexin V staining
For evaluation of cell survival, cells were seeded on 24-well plates at a density of 2 × 104 cells/well, followed by the addition of methyl thiazol tetrazolium (MTT; Sigma) at the end of cell culture. The amount of MTT formazan product was determined using a microplate reader at an absorbance of 560 nm (SpectraMax 250, Molecular Devices, Sunnyvale, CA, USA). Annexin V staining was used to determine the percentage of apoptotic cells. Cells were harvested and stained with fluorescein isothiocyanate-labeled annexin V (BD Biosciences, San Diego, CA, USA) and propidium iodide (PI; Sigma) in the dark at room temperature for 30 min. The percentage of annexin V-positive and PI-negative cells were determined by flow cytometry.
Western blotting
At the end of cell culture, the cell lysates were collected and the protein concentrations were determined using the Protein Assay kit (Bio-Rad, Hercules, CA, USA). Cell extracts with sample buffer were boiled for 5 min and then separated by 10% SDS-PAGE gel. After electrophoresis, the gel was transferred onto a PVDF membrane for immunoblotting. The membrane was blocked by incubation with 5% nonfat milk in Tris-buffered saline Tween-20 (TBST) at room temperature for 2 h, and then incubated with anti-cleaved caspase 3 antibody (Cell Signaling Technology, Beverly, MA, USA), anti-cleaved poly (ADP-ribose) polymerase (PARP) antibody (Cell Signaling Technology), anti-Bcl-2 antibody (Upstate Biotechnology, Waltham, MA, USA), anti-manganese superoxide dismutase (MnSOD) antibody (Upstate Biotechnology), and anti-actin antibody (Chemicon International, Temecula, CA) for 2 h at room temperature or overnight at 4°C, followed by washing for 5 times with TBST and incubation at room temperature with horseradish peroxidase-conjugated secondary antibody for 2 h. The membrane was then washed for 6 times by TBST and specific bands were visualized by chemiluminescence (Santa Cruz Biotechnology, Santa Cruz, CA, USA).
Determination of intracellular reactive oxygen species (ROS) production and glutathione (GSH) content
The measurement of intracellular reactive oxygen species (ROS) production by the probe 2′,7′-dichlorofluorescein diacetate (DCFH-DA; Molecular Probes, Eugene, OR, USA) was mentioned previously [22]. In brief, cells were incubated with 5 µmol/L DCFH-DA in culture medium for 30 min at 37°C, followed by washing with PBS and flow cytometric analysis. The intracellular GSH content was detected by colorimetric assay using the GSH-400 kit (OXIS International, Portland, OR, USA). In the GSH-400 assay, 4-chloro-1-methyl-7-trifluromethyl-quinolinium methylsulfate was added to react with all mercaptans in the sample, leading to the formation of substitution products, thioesters. Then, 30% sodium hydroxide was used to mediate a β-elimination reaction and specifically transform GSH-thioester into a chromophoric thione with a maximal absorbance wavelength at 400 nm that was detected by a spectrophotometer.
In vivo analysis of cell growth and green fluorescence protein imaging
All procedures involving animals were performed in accordance with the institutional animal welfare guidelines of Taipei Veterans General Hospital. A total of 2 × 106 cells were injected into subcutaneous tissue of back skin in nude mice (BALB/c strain) aged 6 weeks and resveratrol was supplemented per os at 30 µg/30 g animal/day. In vivo GFP imaging was visualized and measured by an illuminating device (LT-9500 Illumatool TLS equipped with excitation illuminating source [470 nm] and filter plate [515 nm]). The integrated optical density of green fluorescence intensity was captured and then analyzed by Image Pro-plus software.
Statistical analysis
The results are expressed as the mean ± SD. Statistical analyses were performed by 1-way or 2-way ANOVA, followed by Tukey’s test, as appropriate. A P value <0.05 was considered as statistically significant.
Results
Potential for cardiogenic and neuronal differentiation in iPS cells
Undifferentiated murine iPS cells were cultured on inactivated mouse embryonic fibroblasts (Fig. 1A). These iPS cells expressed GFP driven by the Nanog promoter in undifferentiated conditions (Fig. 1A). Germ-line transmission properties indicated that the integrity of pluripotency was preserved in these cells, as reported previously [5]. In order to drive iPS cells to enter multilineage differentiation in vitro, the hanging drop method was used for embryoid body (EB) formation (Fig. 1A; Supplementary Materials and Methods; Supplementary materials are available online at http://www.liebertpub.com/). Following 6 days of culture in cardiogenic induction medium, iPS EBs began to exhibit spontaneous beating, which was recorded by calcium channel efflux (Fig. 1B). The neuroectodermal differentiation in iPS cells was also investigated. After 14 days of neurogenic induction, neuron-like cells with neurite formation were observed. Immunofluorescence staining confirmed the presence of neurofilament protein in the neurites of these iPS cell-derived neuron-like cells (Fig. 1C; Supplementary Materials and Methods).
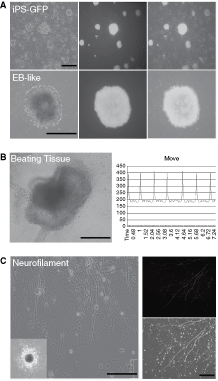
Characterization of mouse induced pluripotent stem (iPS) cells. (
Potential for adipogenic and osteogenic differentiation in iPS cells
In previous studies including ours, mesenchymal stem cells (MSCs) were successfully isolated from bone marrow and induced to differentiate into mesodermal lineages [1,3,4]. Following the protocols used in the aforementioned studies, we were able to differentiate iPS cells into adipocytes after 14 days of culture in adipogenic medium, as confirmed by positive staining with Oil red O and up-regulated expression of PPARγ2 mRNA (Fig. 2A; Supplementary Materials and Methods). To test the potential for osteogenic differentiation, iPS EBs were cultured in osteogenic medium containing 10 nmol/L dexamethasone, and 10 mmol/L β-glycerophosphate. After osteogenic induction for 14 days, the iPS cells exhibited osteocyte morphology and formed a mineralized matrix, as evidenced by von Kossa and Alizarin red staining (Fig. 2B). As measured by real-time RT-PCR, the expression of Runx2 (an osteoblast marker) and osteopontin (OPN; an osteogenesis-related gene) increased significantly after 7 and 14 days of osteogenic induction in iPS cells, as compared with undifferentiated iPS cells (Fig. 2B). In contrast, Oct-4 and Nanog were highly expressed in undifferentiated iPS cells, but their expression was significantly reduced in iPS cells after 7 and 14 days of osteogenic induction (Fig. 2C).
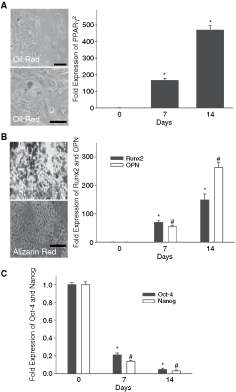
Potential for adipogenic and osteogenic differentiation in induced pluripotent stem (iPS) cells. (
Investigation of the role of resveratrol in osteogenic induction of iPS and ES cells using AFM
To examine the effect of resveratrol on osteogenic induction in iPS and ES cells, 20 µmol/L resveratrol or the corresponding vehicle was added to the osteogenic medium. The addition of resveratrol to the osteogenic medium significantly increased calcium accumulation in iPS and ES cells, as shown by Alizarin red staining (orange-red areas; Fig. 3A and 3B). After 7 or 14 days of induction, the expression of Runx2, OPN, and IBSP (integrin-binding sialoprotein, an osteogenesis-related gene) was significantly higher in iPS and ES cells treated with resveratrol than in those without resveratrol treatment (Fig. 3C). Furthermore, the contact mode of AFM was used to monitor the morphology and hardness of cells during osteogenic induction of iPS cells with or without resveratrol (Fig. 4). Live cells were scanned in a liquid environment using AFM scanning with a triangular silicon probe (Fig. 4A). Both AFM images and force–distance curves (FDC) of iPS cells were acquired at various stages of osteogenic differentiation. In the early stage of induction (Day 1), attached iPS cells showed fibroblast-like morphology and exhibited a flat slope of FDC, as recorded from the AFM and the three-dimensional topography images (Fig. 4B). After 14 days of osteogenic induction, the differentiated iPS cells had a more flattened shape and larger cell size with irregular borders (Fig. 4B), as compared with iPS cells at induction Day 1. At Day 14 of osteogenic induction, the differentiated iPS cells exhibited a sharper FDC slope and significantly greater stiffness as compared with iPS cells at Day 1 of induction (Fig. 4B and 4C). Moreover, the AFM results demonstrated that the elastic moduli of iPS and ES cells treated with resveratrol were significantly lower than those of iPS and ES cells without resveratrol treatment after 7 or 14 days induction (P < 0.05; Fig. 4D). In addition, there was no significant difference in the levels of mineralization, osteogenesis-related gene expression, and mechanical stiffness between iPS and ES cells (P > 0.05; Figs. 3B, 3C, and 4D).
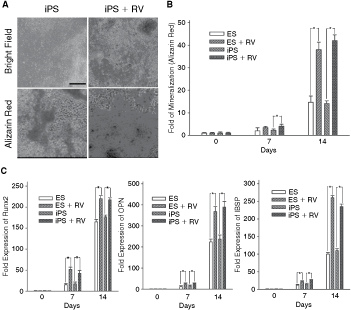
Effects of resveratrol (RV) on osteogenic differentiation in induced pluripotent stem (iPS) and embryonic stem (ES) cells. (
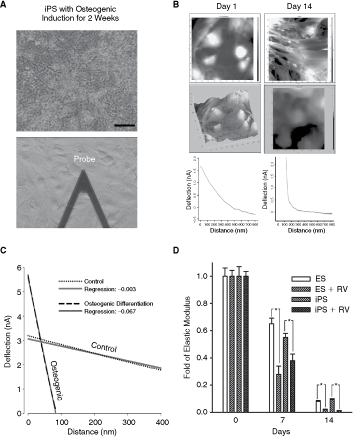
Evaluation of mechanical properties in induced pluripotent stem (iPS) cell-derived osteocyte-like cells by atomic force microscopy (AFM): effects of resveratrol. (
Effects of resveratrol on the survival of iPS cell-derived osteocyte-like cells
Since previous reports indicated that resveratrol was capable of either protecting cells from apoptosis or inducing apoptosis [12,13,23,24], the effects of resveratrol on the survival of iPS cell-derived osteocyte-like cells were examined, using high concentrations of dexamethasone and etoposide as pro-apoptotic stimuli. After 14 days of osteogenic induction of iPS cells with or without 20 µmol/L resveratrol, an equal number of cells were transferred to each well of 24-well plates, and resveratrol or the corresponding vehicle was replenished as in the induction period. Then the cells were treated with 50 µmol/L dexamethasone, 1 µmol/L etoposide or vehicle, followed by survival analysis 6 h later. Although resveratrol had little effect on cell survival in the absence of pro-apoptotic stimuli, it significantly prevented dexamethasone- or etoposide-induced death in iPS cell-derived osteocyte-like cells (Fig. 5A). Pretreatment of cells with resveratrol before dexamethasone or etoposide stimulation also significantly reduced the percentage of Annexin V-positive cells, that is, apoptotic cells (Fig. 5B). Furthermore, resveratrol could efficiently block the dexamethasone- or etoposide-induced cleavage of PARP and caspase 3 (Fig. 5C). In addition, the dexamethasone- or etoposide-induced down-regulation of Bcl-2 was prevented by pretreatment with resveratrol (Fig. 5C). These data suggested that resveratrol had an anti-apoptotic effect on iPS cell-derived osteocyte-like cells.
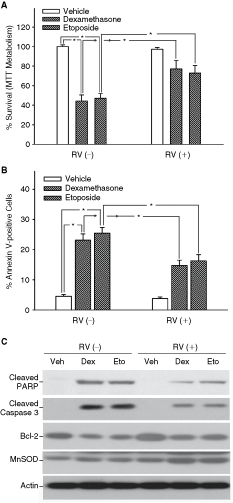
Effects of resveratrol on dexamethasone- and etoposide-induced apoptosis in induced pluripotent stem (iPS) cell-derived osteocyte-like cells. After 14 days of osteogenic induction of iPS cells with or without 20 µmol/L resveratrol (RV), an equal number of cells were treated with 50 µmol/L dexamethasone (Dex), 1 µmol/L etoposide (Eto) or vehicle (Veh) for 6 h. (
Roles of antioxidant effects of resveratrol in its cytoprotection of iPS cell-derived osteocyte-like cells
To evaluate the roles of antioxidant effects of resveratrol in its cytoprotective ability, MnSOD expression, glutathione (GSH) level, and ROS production were measured. Pretreatment of cells with resveratrol before dexamethasone or etoposide treatment significantly up-regulated the MnSOD expression (Fig. 5C) and intracellular GSH level, and decreased ROS production (Fig. 6A and 6B; P < 0.05). The antioxidant and cytoprotective effects of resveratrol were confirmed by challenging cells with an endogenous ROS generator

Roles of antioxidant effects of resveratrol in its cytoprotection of induced pluripotent stem (iPS) cell-derived osteocyte-like cells. (
In vivo effects of resveratrol on osteogenic differentiation of iPS cells
To investigate the effects of resveratrol on osteogenic differentiation in vivo, iPS cells were additionally infected by a lentivector combined with GFP. iPS cells were cultured in osteogenic medium for 7 days (Fig. 7A), and then injected into subcutaneous sites of nude mice at 2 × 106 cells/mouse. The mice were fed with resveratrol (30 µg/30 g animal/day, per os) or control vehicle. Six weeks after transplantation, iPS cells were found to proliferate (Fig. 7B) and GFP signals were detected in the viable transplanted graft (Fig. 7B, arrows). Immunofluorescent assay showed that the expression of OPN was significantly higher in grafts from mice fed with resveratrol than in those from mice fed with vehicle (Fig. 7C). Previous studies have shown that transplanted iPS cells are likely to form teratomas in vivo, a feature also found in ES cells [25]. In the present study, teratoma-like tissues with higher expression of Oct-4 were still detected in grafts from mice transplanted with osteogenic medium-treated iPS cells and fed without resveratrol (Supplementary Fig. 1). No teratoma-like formation was noted in grafts from mice transplanted with osteogenic medium-treated iPS cells and fed with resveratrol (Fig. 7C and Supplementary Fig. 1). To evaluate teratoma formation quantitatively, we measured the expression of embryonic cell stemness/tumorigenicity-related genes and neuroectoderm and endoderm differentiation markers in the grafts. Results of quantitative RT-PCR showed that the expression of Oct-4, Nanog, Klf-4, and c-Myc, genes related to embryonic cell stemness and tumorigenicity, was significantly suppressed in grafts from mice transplanted with osteogenic medium-treated iPS cells and fed with resveratrol (Supplementary Fig. 2). Furthermore, the expression of Sox2 (an ES cell/neural stem cell marker), Neurogenin2 (a brain development-related gene), HNF3β (a liver development-related gene), and PDX-1 (a pancreatic progenitor marker) was also significantly down-regulated in grafts from mice transplanted with osteogenic medium-treated iPS cells and fed with resveratrol (Supplementary Fig. 3). Taken together, these results of transplantation experiments demonstrated that resveratrol can effectively promote osteogenic differentiation and significantly inhibit tumorigenicity in vivo.

In vivo effects of resveratrol on osteogenic differentiation of induced pluripotent stem (iPS) cells transplanted to nude mice. (
Discussion
Mineralization by bone-lining cells and osteocytes plays an important role in osteogenic formation, tissue stiffness, and clinical outcome of bone healing. The recent progress in the measurement of nano-indentation modulus with AFM has been applied to examine the elastic properties of individual human bone lamellae under dry and physiological conditions [26], and to investigate demineralization/remineralization cycles at human tooth enamel surfaces [15,27]. Recently, researchers expanded the use of this technique to measure mechanical properties in bone and osteocytes, and even to monitor osteogenic differentiation in MSCs and osteoblasts [28,29]. In this study, we investigated the osteogenic potential of iPS cells, and AFM was used to monitor the changes in stiffness and elasticity in living iPS cells undergoing osteogenic differentiation. We found that, during the process of osteogenic induction, the stiffness of differentiating iPS cells (Fig. 4) rose concomitantly with increased osteogenic gene expression and mineralization (Figs. 2 and 3). Recently, Bäckesjö et al. demonstrated that resveratrol as a SirT1 agonist markedly decreased adipocyte development and promoted osteoblast differentiation in MSCs [11]. In accordance with the aforementioned report, we found that resveratrol exhibited a potent capacity to facilitate osteoblast differentiation in both iPS and ES cells, as evidenced by increased mineralization, osteogenic gene expression, and stiffness (Figs. 3 and 4). Furthermore, our data suggested that iPS cells were indistinguishable from ES cells in the osteogenic differentiation capability. Importantly, we demonstrated that resveratrol given per os markedly enhanced osteogenic differentiation of transplanted iPS cells (Fig. 7). To our knowledge, this is the first study to report that resveratrol could induce differentiation of iPS cells into osteocyte-like cells with mechanical properties similar to osteocytes, and also promote osteogenic differentiation in vivo.
Glucocorticoids have been extensively used for the treatment of immune and inflammatory conditions [23]. However, long-term use of glucocorticoids is associated with severe adverse effects on several organs, including the skeleton [23,30]. Glucocorticoid-induced osteoporosis is the most common form of secondary osteoporosis [23]. In the present study, we demonstrated that resveratrol could protect iPS cell-derived osteocyte-like cells from glucocorticoid-induced cytotoxicity. The addition of resveratrol before dexamethasone treatment significantly reduced the percentage of apoptotic cells and the level of activated caspase 3, and up-regulated the expression of Bcl-2 (Fig. 5). Our results were supported by a recent study showing that resveratrol was able to increase Bcl-2 levels in endothelial cells and prevent peroxynitrite-triggered caspases-3 and -9 activation and apoptosis [31]. Dudley et al. also demonstrated that resveratrol at low dose provided cardioprotection through up-regulating Akt and Bcl-2 protein [32]. Whether resveratrol can prevent glucocorticoid-induced osteoporosis needs further research.
ROS play a critical role in limiting lifespan of organisms as well as initiating the process of cellular senescence [33]. Increasing levels of ROS have also been shown to restrict the lifespan of hematopoietic stem cells [34]. Recently, Robb et al. have demonstrated that resveratrol can dramatically induce mitochondrial MnSOD expression and activity in MRC-5 cells as well as in mouse brain tissues [35,36]. Moreover, resveratrol has also been shown to protect ROS-induced cell death by activating AMP-activated kinase in cardiac muscle cells [37]. In this study, we found that resveratrol could dramatically reduce glucocorticoid-induced ROS accumulation and up-regulate intracellular GSH levels and MnSOD expression in iPS cell-derived osteocyte-like cells (Fig. 6A and 6B). Since resveratrol rescued cells from glucocorticoid-induced damage and the cytoprotective ability of combined resveratrol and superoxide scavenger Tempol treatment was not superior to that of Tempol alone, we proposed that resveratrol promoted survival in iPS cell-derived osteocyte-like cells largely by its antioxidant effects (Figs. 5 and 6). Future studies are required to examine the role of ROS in the self-renewal and differentiation of iPS cells and the involvement of ROS reduction in resveratrol-promoted osteogenic differentiation.
The ability to form teratomas in vivo has been a landmark and routine assay for evaluating the pluripotency of ES as well as iPS cells [25]. However, teratoma formation from pluripotent stem cells is considered as an unacceptable obstacle for the application of stem cell therapy in regenerative medicine. Therefore, measures to overcome the tumorigenicity of iPS cells are crucial for successful treatment of patients with iPS cells. Reservatrol has been found to exhibit the anticancer potential and/or prevent tumorigenesis through activation of SirT1, which inhibits the activity of surviving [38,39]. Our previous study has demonstrated that reservatrol can effectively induce CD133-positive cancer stem-like cells to differentiate into CD133-negative cancer cells, and further enhance the chemoradiotherapeutic effects on malignant brain tumors [40]. In the present study, we found that resveratrol treatment in iPS cell-bearing mice could significantly down-regulate the mRNA expression of ES stemness genes in the graft and effectively blocked the teratoma formation (Fig. 7 and Supplementary Fig. 2). Further studies are required to examine the roles of resveratrol in the inhibition of tumorigenicity in various models of transplantation therapy using iPS cells. In this study, we also demonstrated that resveratrol greatly enhanced osteogenic differentiation of iPS cells and effectively protected iPS cell-derived osteocyte-like cells from glucocorticoid-induced cytotoxicity. In the future, iPS cells may be applied to the treatment of osteoporosis and other orthopedic diseases by resveratrol-induced osteogenic differentiation and cytoprotection.
Footnotes
Acknowledgments
The authors thank the RIKEN BRC, Japan, for providing the biological resources used in this study. This study was supported by research grants from NSC (97-3111-B-075-001-MY3), Taipei Veterans General Hospital (V97B1-006, E1-008, ER2-018, ER3-005, F-001), the Joint Projects of UTVGH (VGHUST 98-G6-6), Yen-Tjing-Ling Medical Foundation, National Yang-Ming University (Ministry of Education, Aim for the Top University Plan), and Technology Development Program for Academia, Department of Industrial Technology, Ministry of Economic Affairs, Taiwan.
Author Disclosure Statement
The authors declare no conflict of interest.