
Abstract
The recent findings indicate that under conditions of severe tubular injuries, transplantation of mesenchymal stem cells (MSCs) may be a promising treatment in acute kidney diseases; nevertheless, the underling mechanism is still under debate. To investigate the differentiation characteristics and the role of MSCs in renal tubular injury, human adipose-derived MSCs (hAD-MSCs) were transplanted into ischemia–reperfusion (I/R) kidneys in C57BL/6 mouse model. Results showed that hAD-MSCs were able to differentiate toward renal tubular epithelium at an early stage of injuries. The differentiated donor cells replaced the vacant space left over by the dead cells, contributed to maintenance of structural integrity and proceeded to a subsequent tissue repair process. Furthermore, MSCs as supportive cells may promote repair via secreting cytokines. The differentiation and replacement of MSCs at an extremely early stage play important roles for the subsequent self-repair and -renewal of functional cells. Direct differentiation of MSCs, as an important mechanism of injured kidney repair, warrants further investigation.
Introduction
A
Despite recognition of the efficacy of MSCs on kidney injuries, the underling mechanism is still under debate. In general, preliminary studies showed that replacement of the injured tissue by trans-differentiated donor cells contributed to the regeneration [2,3]; however, there are increasing evidences that other differentiation-independent mechanisms of MSCs play a key role in promoting tissue repair [4 –6]. Some studies even showed that MSCs did not differentiate into tissue-specific cells such as tubular epithelial cells during the repair process [4,5].
Abundant works of our laboratory have demonstrated multilineage plasticity potential of MSCs [7], including epithelial tissues such as lung and small intestine [8 –10], and their immune regulatory effect and low immunogenicity [11 –14]. With their low immunogenicity, we had transplanted hAD-MSCs into immunocompetent mdx mice and detected their engraftment and differentiation in vivo [15]. To further investigate the fate and possible application of hAD-MSCs in renal tubular injury, hAD-MSCs were transplanted into ischemia-reperfusion (I/R) kidneys in an immunocompetent C57BL/6 mouse model to observe their homing and differentiation, and to elucidate the related mechanisms.
Materials and Methods
Isolation of MSCs from adult human adipose tissue
Human adipose tissue was obtained from patients undergoing tumescent liposuction, according to procedures approved by the Ethics Committee at the Chinese Academy of Medical Sciences and Peking Union Medical College. The isolation procedure of Cao was modified [11]. The adipose tissue was washed with D-Hank's BSS (∼3 mL/g) and then resuspended in 0.075% type IA collagenase (Sigma, St. Louis, MO)/D-Hank's BSS (∼2 mL/g) and incubated at 37°C for 1 h. The digested adipose tissue was passed through a 100-μm filter to remove debris and then centrifuged at 1,200 rpm for 10 min to obtain a cell pellet. The pellet was resuspended and washed twice with D-Hank's BSS.
Animals and isolation of MSCs from mouse bone marrow
eGFP C57BL/6 mice (The Jackson Laboratories, Bar Harbor, ME) and Rosa26 mice (B6; 129S-Gtrosa26; The Jackson Laboratory) of 5–6 weeks were used as donors. Recipient C57BL/6 mice of 7–8 weeks were purchased from the Animal Center of the Chinese Academy of Military Medical Sciences (Beijing, China). All animals used were of the same genetic background. They were bred and maintained under specific pathogen-free conditions. All animal handle and experiment procedures were approved by the Animal Care and Use Committee of the Chinese Academy of Medical Sciences.
Bone marrow cells were obtained from donor mice by flushing bone marrow from the femur and tibia. Mouse bone marrow-derived MSCs (mBM-MSCs) were prepared as previously described [12]. The cells were 1:1 diluted with D-Hank's BSS and centrifuged in 20 mL of 1.077 g/mL Ficoll-Paque at 1,500 rpm for 20 min. The interface was collected and centrifuged at 1,200 rpm for 10 min to obtain a cell pellet. The pellet was resuspended and washed twice with 20 mL of D-Hank's BSS.
Expansion of MSCs and labeling
After isolation, 30 mL of resuspended cells were plated in expansion medium at a density of 5 × 106 nucleated cells/100-mm tissue culture dish and incubated at 37°C in a humidified environment containing 5% CO2. Expansion medium contained 58% DF-12 medium (Dulbecco's modified Eagle's medium/Ham's F-12 medium; Gibco, Life Technologies, Paisley, United Kingdom), 40% MCDB (medium complete with trace elements-201; Sigma), 2% FCS (Gibco), 1× insulin-transferrin-selenium (ITS; Gibco), 1 × 10−9 mol/L dexamethasone (Sigma), 1 × 10−4 mol/L ascorbic acid 2-phosphate (Sigma), 10 ng/mL EGF, 10 ng/mL PDGF-BB (Sigma), 100 U/mL penicillin, and 100 μg/mL streptomycin (Gibco). The third passage of human adipose-derived MSCs (hAD-MSCs) and mouse BM-MSCs (mBM-MSCs) were used. The immunologic phenotypes of hAD-MSCs were positive for CD29, CD44, CD105, Flk-1 and negative for CD31, CD34, CD45, and HLA-DR, as described before [16].
For Feridex labeling, hAD-MSCs were incubated overnight with micron-sized particles of iron oxide (MPIO, 2 μL/cm2, encapsulated magnetic microspheres; Bangs, Fishers, IN) on the day before transplantation [17].
Ischemia/reperfusion experiments and cell transplantation
Mice were anesthetized with 100 mg/kg ketamine and 10 mg/kg xylazine injected intraperitoneally, and a midline incision was made. For unilateral ischemia/reflow (I/R), the right renal pedicle was clamped for the indicated time using a vascular clamp. The abdomen was covered with gauze moistened in PBS, and the mice were maintained at 37°C using a warming pad. After 35 min, the clamp was removed, followed by reperfusion and closing of the abdominal wound.
Then the I/R recipients were injected with 5 × 105 third passage MSCs in 0.3 mL physiologic saline (PhyS) via tail vein immediately (in 24 h) in the treatment groups. Control animals were treated identically but infused with 0.3 mL PhyS without cells.
Histopathology
At 3 days, 21 days, and 6 months post I/R, both treated and untreated mice were euthanized and the right kidneys were thoroughly perfused with saline to remove blood from the vascular beds. The kidneys were then resected from the abdominal cavity and extraneous tissue cleared off. The specimens of the right kidney were fixed in 4°% paraformaldehyde for 24 h, dehydrated, and embedded into paraffin wax, then sectioned to 4 μm slides and processed for hematoxylin-eosin staining and Masson trichrome staining.
To quantify the renal interstitial fibrosis, the extent of fibrosis (blue-staining area) was measured on photographs of Masson trichrome-stained sections using Image-Pro Discovery 5.1 software. We defined the fibrosis rate as the radio between the blue area and the whole area. For statistical analysis, we randomly took 25 photos from 5 mice in each group.
Immunochemistry staining
The 4-μm tissue sections were deparaffinized, washed in PBS, incubated in 3% hydrogen peroxide for 10 min, and then washed in PBS. The nonspecific binding was blocked by incubating the sections in serum for 60 min. The slides were then incubated with the first antibody for 1 h, and incubated with biotin-conjugated secondary antibodies for 15 min at room temperature, colorated with DAB, counterstained with hematoxylin, and then observed under microscopy.
The first antibodies we used were goat monoclonal antibody (mAb) against GFP (Abcam, Cambridge; 1:1,000), mouse anti-β-galactosidase mAb (Promega, San Luis Obispo, CA; 1:1,000), rabbit anti-PCNA mAb (Promega; 1:200), and mouse anti-human Nuclei mAb (Chemicon, Billerica, MA; 1:25, need a 20-min microwave pretreatment in citrate buffer).
Prussian blue staining
The 4-μm tissue sections were incubated for 30 min with 2% potassium ferrocyanide in 2% hydrochloric acid, washed, and counterstained with nuclear fast red. Images were taken using a DP-70 microscope (Olympus, Tokyo, Japan).
In situ hybridization
In brief, 4-μm kidney sections were deparaffinized and rehydrated, incubated in 3% hydrogen peroxide for 10 min, and then washed in 0.1 mol TBS. Antigen retrieval was performed at 37°C for 10 min and washed in TBS. After denaturation at 70°C, slides were incubated with the denatured probe at 37°C overnight followed by washing in 2× SSC, performed with POD transforming agent at 37°C for 45 min, and colorated with DAB.
Double immunofluorescence staining
For double immunofluorescence staining, the deparaffinized sections were microwave-heated in citrate buffer for 20 min, washed in PBS, blocked with (rat and/or goat) serum, and incubated with rabbit polyclonal anti-pan-CK (Santa Cruz Biotechnology, Santa Cruz, CA; 1:200) or PCNA and mouse anti-human nuclei mAb at room temperature for 1 h. The slides were then washed in PBS and incubated with TRITC-coupled goat anti-mouse IgG and FITC-coupled goat anti-rabbit IgG secondary antibodies (Santa Cruz; 1:200) for 30 min at room temperature. Finally, slides were counter-stained with 5 μg/mL Hoechst 33342 (Sigma).
In vitro epithelial differentiation
Renal tubular epithelial cells were isolated from normal C57BL/6 mice as described by Lin et al. [18]. Approximately 5 × 105/mL cells were resuspended in DF-12 medium, exposed to 10.0 Gy at a dose rate of 1.0 Gy per min, and then cultured for 12 h. Medium without cells were collected by percolating and then supplemented with DF-12 medium (1:4), 20 ng/mL hepatocyte growth factor (HGF), 10 ng/mL fibroblast growth factor 4 (FGF-4), and 2% FBS. This mixed medium was used as epithelial differentiation medium.
The third passage hAD-MSCs (2 × 104/cm2) were induced for 3 weeks for epithelial differentiation in epithelial differentiation medium. Undifferentiated MSCs served as negative control and human renal tubular epithelial cell line HK-2 served as positive control.
Reverse transcriptase-polymerase chain reaction (RT-PCR) analysis
Cultured cells or tissue samples were collected and total RNA was prepared as described [10]. PCR analysis for epithelial-specific markers CK18, human β-actin, and housekeeping gene β-actin were performed using primers that span the genes (94°C 60 s, 60°C 60 s, 72°C 60 s, 35 cycles). PCR products were quantified densitometrically using GelWorks software after scanning the ethidium bromide-stained 2% agarose gel with GDS8000 (Ultra-VioletProducts Inc., Upland, CA). The primers used were as follows: β-actin F, 5′-GCT CCT CCT GAG CGC AAG TA-3′; R, 5′-GAT GGA GGG GCC GGA CT-3′; CK18 F, 5′-CGC ATC GTC TTG CAG ATC GAC-3′; R, 5′-GCT GAG ACC AGT ACT TGT CCA G-3′; human β-actin F, 5′-CTG GAA CGG TGA AGG TGA CA-3′; R, 5′-AAG GGA CTT CCT GTA ACA ATG CA-3′.
Cell immunofluorescence staining
For in vitro experiments, the above cultured cells were digested into single cells and resuspended in DF-12 medium with 10% FCS to a concentration of about 105 cells/mL. The mixture were dropped onto sterile glass plates and incubated at 37°C in a 5% CO2 humidified atmosphere for 2 h to make cells adherent. Then the plates were washed by PBS and fixed with 4% paraformaldehyde at 20°C. The slides were incubated sequentially with rabbit mAb against pan-CK (Sigma; 1:200) for 1 h, followed by FITC-labeled anti-rabbit IgG antibody. The sections were counterstained with 5 μg/mL Hoechst 33342 (Sigma).
Statistical analysis
Statistical analysis was performed with the statistical SPSS 10.0 software. The paired sample t-test was used to test the probability of significant differences between samples. Statistical significance was defined as P < 0.05.
Results
Rescue effects of hAD-MSCs on injured kidneys
A renal ischemia/reperfusion (I/R) model was employed in this study. The mice were divided into transplantation group and the control group, which received 5 × 105 hAD-MSCs and saline in 24 h post-injury, respectively. The right kidneys were obtained from the 2 groups for histopathology 3 days, 21 days, and 6 months post-I/R injury, respectively. At 3 days after I/R, HE staining of kidneys in control group showed marked edema in proximal convoluted tubules with infiltration of inflammatory cells (Fig. 1D). No visible edema was observed in the transplantation group (Fig. 1A). At 21 days, infiltration of abundant inflammatory cells in the renal interstitium, necrosis of the tubular epithelium, loss of epithelial structure, and fibrosis were observed in the control group (Fig. 1E). In contrast, the tubular structure was intact in the transplantation group, with only a few inflammatory cells in renal interstitium (Fig. 1B). Six months after I/R mice in the transplantation group regained normal renal structure (Fig. 1C), while renal tubule around partial glomerule reduced, glomerular aggregation and fibrosis developed in the control group (Fig. 1F).
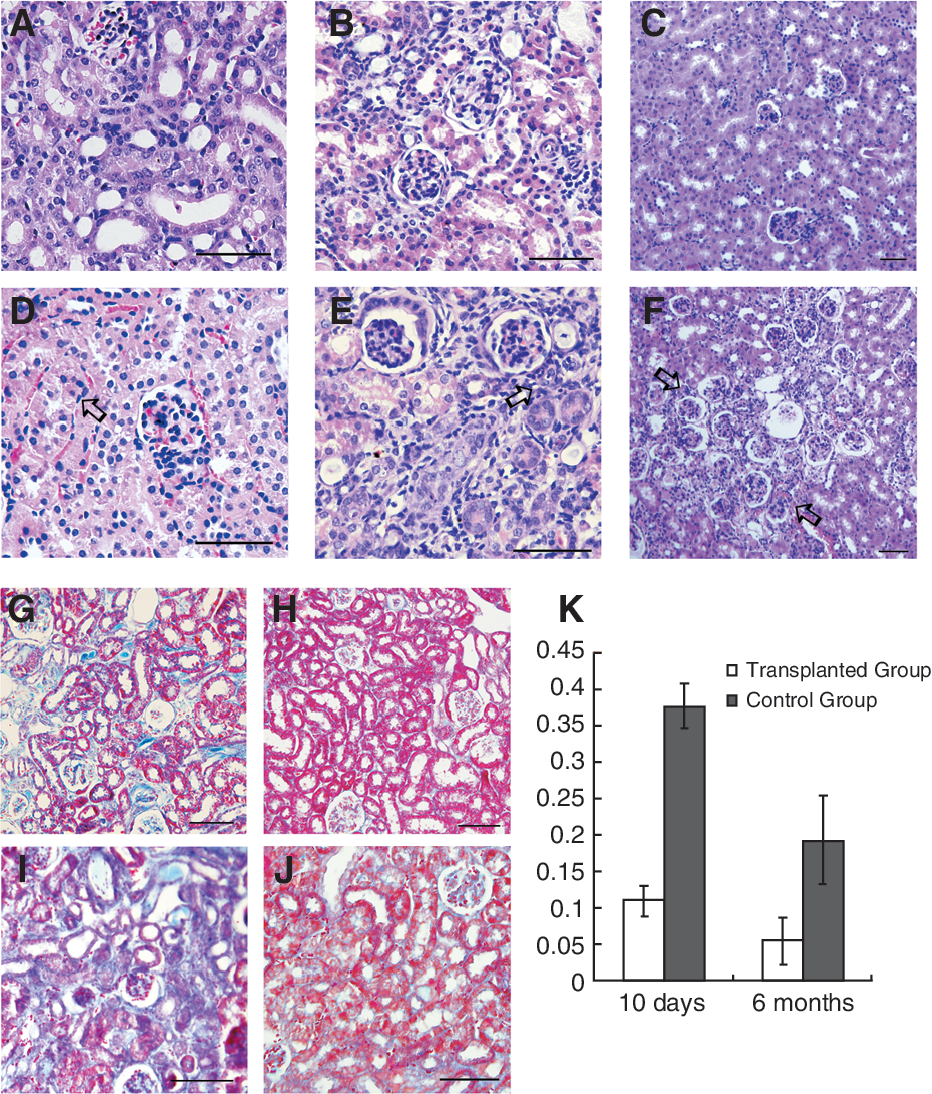
The therapeutical effects of human adipose-derived MSCs (hAD-MSCs) on injured kidneys. Figure 1A–1F showed H.E. staining of transplantation group and the control group at 3 days, 21 days, and 6 months post-renal I/R injury, respectively. At 3 days after I/R, no edema was observed in the transplantation group (
Masson staining was performed for quantitative measurement of collagen deposition in the kidney. Collagen deposition rate was defined as the ratio of the area of collagen deposition (blue) to the total tissue area (Fig. 1K). For mice sacrificed 10 days after I/R, marked collagen deposition was noted in the control kidneys (Fig. 1I) while the amount of collagen in transplantation group was significantly reduced (Fig. 1G) (deposition rate 0.38 ± 0.03 vs. 0.11 ± 0.02, P < 0.05). At 6 months, collagen deposition rate in transplantation group (Fig. 1H) was also significantly lower than the control (Fig. 1J) (deposition rate 0.05 ± 0.03 vs. 0.19 ± 0.06, P < 0.05). Thus transplantation of MSCs is effective in alleviating the kidney injuries and inhibiting fibrosis in the I/R mice.
MSCs engraftment in I/R-injured kidney
Based on the previous reports and our studies, there is no doubt that MSCs have rescue effects on injured kidney [19,20]. We next wanted to know if MSCs directly differentiated into renal tubular epithelial cells after infusion. hAD-MSCs labeled with Feridex were transplanted into the I/R mice. The kidney tissues were first tested by RT-PCR 3 days, 10 days, and 6 months after transplantation to detect the exogenous expression. Human-specific β-actin was detected only at 3 and 10 days post-transplantation, not at 6 months (Fig. 2A). These results suggested that MSCs could short-term engraft into ischemic/reperfusive kidneys.
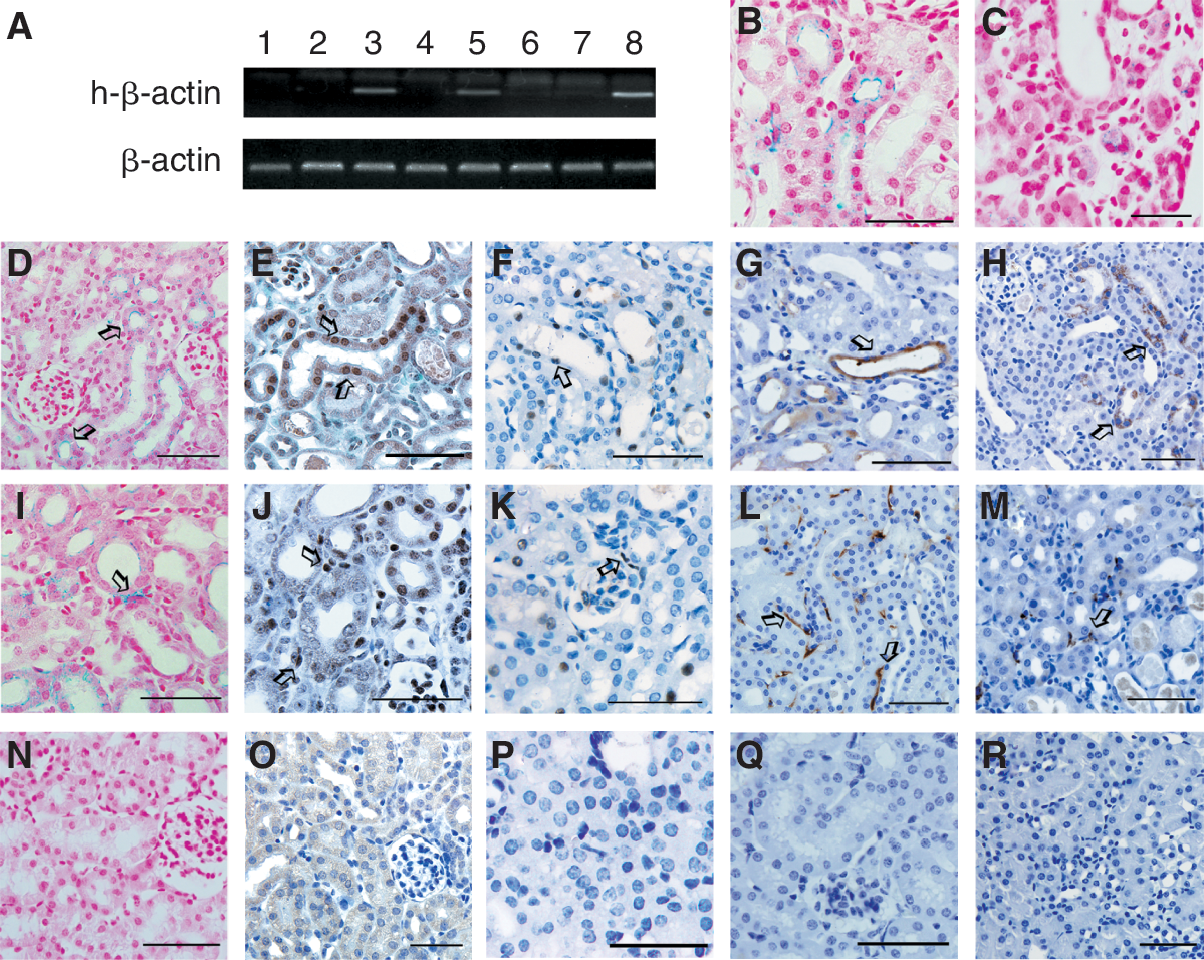
Engraftment of culture-expanded mesenchymal stem cells (MSCs) in I/R-injured kidneys. RT-PCR results of kidney samples of C57BL/10 mice receiving hAD-MSCs injection at 3 days (lane 3), 10 days (lane 5), and 6 months (lane 7) after injection of hAD-MSCs indicates that transplanted cells could not be detected after 6 months (
Then we analyzed the distribution of donor-derived cells at 3 and 10 days after transplantation by Prussian blue staining. On day 3, as shown in Figure 2B, donor cells (with blue granules) could be clearly seen in renal cortical tubules and interstitium. Though with a significant lower number, the distribution of donor cells at 10 days (Fig. 2C) was similar with 3 days (Fig. 2B). According to these results, we chose third-day post-transplantation as the analysis time point for engraftment and differentiation.
Besides Prussian blue staining for Feridex (Fig. 2D and 2I), at 3 days after I/R, immunohistological staining with anti-human nuclei antibodies also detected human cells similarly in renal cortical tubules (Fig. 2E) and interstitium (Fig. 2J) in murine kidneys. Then, we used Y chromosome tracing to further determine whether injected hAD-MSCs could ever be detected in the renal parenchyma of I/R-treated mice. For these experiments, we used male hAD-MSCs injected intravenously into I/R-treated female mice. In situ hybridization for the Y chromosome was performed on multiple sections. Y chromosome-positive tubular cells (Fig. 2F) and interstitial cells (Fig. 2K) were detected.
In addition, the engraftment of mouse bone marrow-derived MSCs (mBM-MSCs) was analyzed in the same ischemia-reperfusion kidney model. As expected, the similar results were observed as those in hAD-MSCs transplantation group. Rosa mice and eGFP transgenic mice were used as donors of mouse bone marrow. On day 3, β-gal- (Fig. 2G and 2L) and green fluorescent protein (Fig. 2H and 2M)-positive cells were detected in renal cortical tubules (Fig. 2G and 2H) and interstitium (Fig. 2L and 2M).
The above 5 detection methods are the most frequently used for cell fate determination after transplantation. Our results showed that hAD-MSCs, as mBM-MSCs, did engraft in the interstitial and cortical tubular areas of the injured kidneys.
Engrafted hAD-MSCs differentiate into renal tubular epithelial-like cells
To determine whether the transplanted cells seen in the tubules did differentiate into tubular epithelial cells, double immunofluorescence with pan-CK and anti-human nuclear antibodies were performed on the kidney tissue slices 3 days after hAD-MSC transplantation. Cells positively stained by both human nuclear antibody (red fluorescence) and pan-CK antibody (green fluorescence) were observed in transplantation group (Fig. 3A–3C), indicating that the injected MSCs had differentiated into renal tubular epithelial-like cells. The control group was negative for human nuclear antibody and positive for pan-CK antibody (Fig. 3D–3F). The kidney tissues were also tested by RT-PCR to confirm this conclusion. Indeed, both human-specific β-actin and human-specific epithelial marker CK18 were detected on third day after transplantation (Fig. 3G). The control group was negative for both markers. These results indicated that transplanted hAD-MSCs homed to the ischemic-reperfusive kidneys gave rise to tubular epithelial-like cells and contributed to the repair of the kidney lesions at least at the early phase after injury.
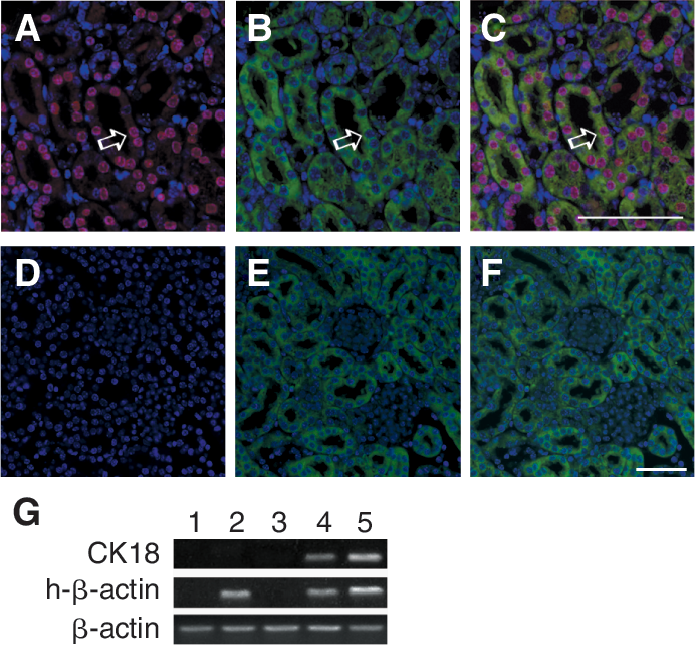
Differentiation of culture-expanded hAD-MSCs in I/R kidneys. Differentiation of hAD-MSCs into epithelial cells was demonstrated by double IF staining of anti-human nuclear antibodies (red) and pan-CK (green) in kidney specimens from I/R-injured mice at 3 days after hAD-MSCs transplantation (
hAD-MSCs were induced into epithelial cells in vitro
To further confirm that MSCs are able to give rise to tubular epithelial cells, we next conducted cell induction experiment in vitro. Single hAD-MSC was amplified and cultured in the renal tubular epithelial induction medium for 2 weeks. Marked morphologic changes were observed under light microscope, that is fusiform cells were gradually turned into epithelial polygonal cells (Fig. 4A and 4B). Immunofluorescence revealed that about 60% of the cells expressed pan-CK after induction (Fig. 4C), which was negative before induction (Fig. 4D), and RT-PCR also demonstrated that these cells expressed CK18 (Fig. 4E). This in vitro experiment confirmed the differentiation potential of hAD-MSCs toward epithelium in vivo.

Culture-expanded hAD-MSCs differentiated into epithelial cells in vitro. hAD-MSCs with fusiform-shaped (
Donor hAD-MSCs promote host cell proliferation
Proliferating cell nuclear antigen (PCNA) antibody was used to investigate cell proliferation in I/R kidneys of the transplantation group and the control group. Three days after I/R, immunochemistry assay showed significantly more proliferating cells in kidneys of the transplantation group (Fig. 5A) than the control group (Fig. 5B); 10 days after I/R, the number of proliferating cells decreased in the transplantation group (Fig. 5C), but increased in the control group (Fig. 5D). Those results indicated that MSCs can accelerate kidney repair process by enhancing proliferation, consistent with previous studies [15]. To determine whether these proliferating cells were donor MSCs that differentiated into renal tubular epithelium, double immunofluorescent staining with PCNA and anti-human nucleus antibodies was performed. Only a few double positive cells were observed, that is donor cells per se did not undergo marked proliferation (Fig. 5E–5G). Whereas, we observed that area having more donor cells also had more proliferating cells suggested that hAD-MSCs promoted endogenous regeneration that might act through paracrine processes.
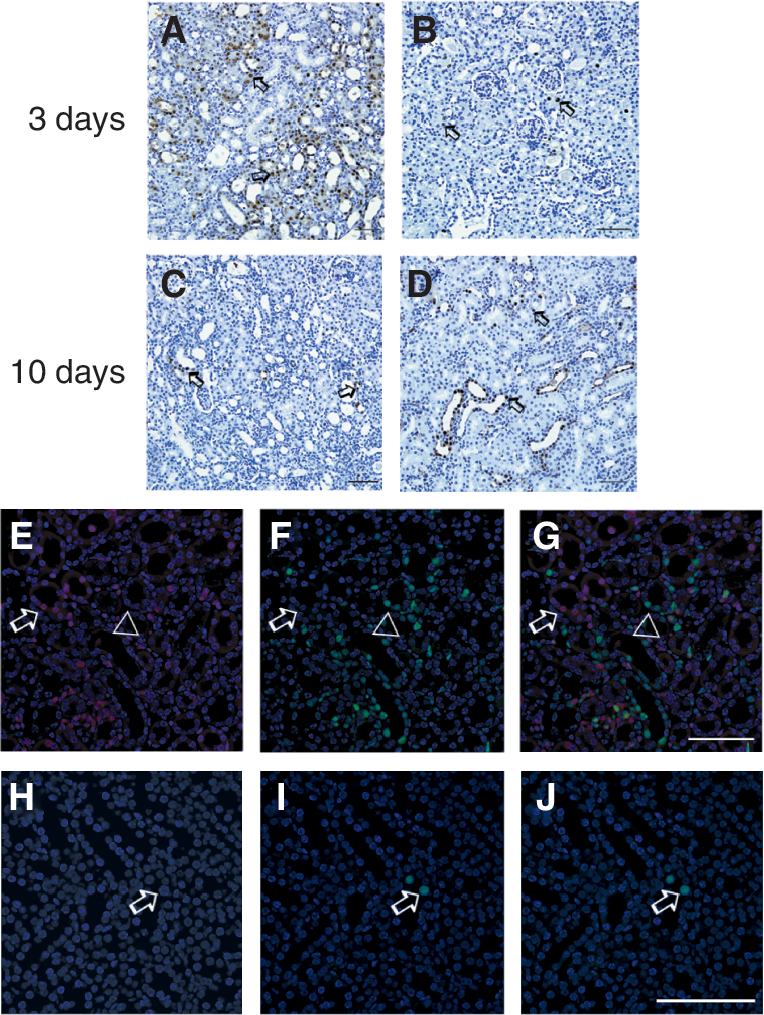
Donor mesenchymal stem cells (MSCs) may promote host cell proliferation. Immunohistochemistry staining of PCNA antibody was used to investigate cell proliferation in I/R kidneys of the transplantation group and the control group (arrow). Three days after I/R, immunochemistry assay showed significantly more proliferating cells in kidneys of the transplantation group (
Based on the above results, besides directly differentiating toward renal tubular epithelial cells, MSCs promoted restoration of post-I/R kidneys via activating endogenous repair process.
Discussion
In the past 10 years, enormous studies showed that MSCs have a greater differentiation potential than previously expected. In our laboratory, we also isolated a subpopulation of MSCs (positive for Flk-1 and negative for CD34) from the adult tissue, including bone marrow and adipose [10,16,21]. These cells have a definite phenotype that may facilitate studies. These cells possess extensive differentiating potentials, and may contribute to the repair of different organs and tissues. During recent years, adipose-derived MSCs have gained wide interest due to its similar differentiating potential to that of BM-MSCs and easy availability [22]. In this study, hAD-MSCs were transplanted into the mice with impaired kidneys and results showed that hAD-MSCs were able to differentiate toward renal tubular epithelium at early stage of injuries, which is important for the kidney to recover from I/R injury.
The kidney I/R model has been developed and widely used for studies on renal tubular injuries and renal interstitial fibrosis. The severity and duration of ischemia has direct effect on the prognosis of the tubular injuries [3]. After short-term renal ischemia, pathological manifestations include tubular edema and infiltration of a few inflammatory cells, which may recover in 1 or 2 weeks after the etiology is eliminated, without significant renal function impairment. If the duration of ischemia is prolonged, the damage to the renal tubules will be worsened and manifest pathologically as changes to the cell polarity, collapse of the tubular epithelial basement membrane, loss of integrity, and consequent damage to the tubular cells or even cellular necrosis [23,24]. Sometimes the damage may be reversed after removal of the etiology. But irreversible damage may also happen and present as chronic fibrosis and permanent renal function impairment [25]. In preliminary experiments, through trial and error, we established that 35 min of ischemia may cause a moderate to severe renal interstitial fibrosis in mouse.
Adipose-derived MSCs were infused to the mouse after I/R via tail veins. Three days after transplantation, HE staining showed renal tubules and glomerules remained histologically and structurally intact with less inflammatory infiltration in the transplantation group as compared to the control; at 6 months, structurally normal kidneys were observed in the transplantation group while fibrosis was marked in the control group. Masson staining of the late stage kidneys also revealed significantly less collagen deposition in the transplantation group than control. These results, in accordance with reports published by several other laboratories, confirmed potential therapeutic effects of MSC transplantation on injured kidneys and renal fibrosis.
In spite of the therapeutical effects of MSCs on renal injuries, the mechanisms underlying the repair process still remain controversial [26,27]. The question focuses on whether MSCs could directly differentiate into renal tubular epithelial cells. To make the results more convincing, besides human adipose-derived MSCs that were detected by in situ hybridization, anti-human nucleus antibody, and Feridex labeling, mouse bone marrow-derived MSCs as the most commonly studied cells were also chosen in the engraftment study. Rosa mice and eGFP mice were used as donors of the BM-MSCs for easy detection of donor cells. Compared to direct observation of GFP positive cells derived from eGFP mice by laser scanning confocal microscopy [18], immunofluorescent studies are known to be more reliable. Assays were undertaken 3 and 10 days after transplantation, and similar results were obtained from each group, that is there were donor cells in the cortical tubules and interstitium. Similar results obtained from cells from different species and with different labeling methods provide the advantage of avoiding false positive results associated with single tracing method. Further, immunofluorescent double staining demonstrated the expression of pan-CK, an epithelial marker, by donor cells in the tubular area. The results indicated that exogenous hAD-MSCs do have the ability to differentiate toward renal tubular epithelial cells and contribute to kidney repair. And this conclusion was confirmed by RT-PCR assays of human-specific house-keeping genes and human-specific epithelial gene CK18. In vitro induction experiments also showed that MSCs were able to differentiate into epithelial cells.
We think one reason for the discrepancy on whether the transplanted donor MSCs are able to give rise to renal tubular epithelium may be due to the diversity in MSCs used by diverse researchers. MSC is a cell population with multipotent differentiation capacities. MSCs isolated and cultured at various institutes showed distinct surface markers and variable differentiating potentials, which may explain the findings' disparation [3,26,28]. MSCs isolated from bone marrow and adipose tissues at our laboratory have multilineage differentiating potentials and definite phenotype, that is, negative for CD11a, CD31, CD34, CD45, and HLA-DR and positive for CD29, CD44, CD105, and Flk-1 [7 –16].
Supportive effects of MSCs have been well-documented [29]. Since proliferation is the major pattern of tissue repair, PCNA antibody was used to detect cellular proliferation after kidney I/R injuries. Three days after I/R, cellular proliferation level was significantly higher in the transplantation group than in the control, suggestive of the effect of MSCs on tissue repair at early stage. Next, immunofluorescent double staining with PCNA and anti-human nuclear antibodies showed that massive proliferating cells were not donor cells but host cells around donor cells. Thus MSCs may activate the innate repair cascade [30]. This result was consonance with the previous reports that MSCs were able to aid in renal tissue repair through differentiation-independent mechanisms, which in our opinion, besides the restoration of tubular epithelial cells during repairing, probably by paracrine processes. MSCs have been shown to secrete growth factors such as HGF, VEGF, IGF-1, and EGF [31 –33], which might promote the endogenous proliferating and repairing. There were studies addressed that HGF and IGF-1 reduced tubular injury when given to mice subjected to either toxic or ischemic acute kidney injury [34,35]. Meanwhile, VEGF can mediate endothelial as well as epithelial cell proliferation and survival after injury [36,37], and a vasculotropic paracrine actions of MSCs displayed important role to the recovery from acute kidney injury [38]. Meanwhile, other variety of factors secreted by MSCs, including interleukin members and leukemia-inhibitory factor [39]; TGF-(β1; and prostaglandins such as PGE2 can inhibit lymphocyte activation and thereby suppress the inflammatory responses that might otherwise protect against the tubular injury and apoptosis[32,33,40].
Our results confirmed that MSCs also promote kidney tissue repair via direct differentiation. At the early stage of tissue injury, bulky cell death results in severe cellular vacancy [41], and tissue repair is initiated by cell recruitment in vivo in response to local cues. Transplanted MSCs at this stage, under the microenvironmental regulations, differentiate into renal tubular epithelial cells or epithelial-like cells to replace the vacant space left over by the dead cells, like the “first-aids.” Structural integrity may not be promptly restored by self-repair due to large amount of cell death. The emergent aid from these MSCs contributes to maintenance of structural integrity and proceeding to subsequent tissue repair process. Furthermore, MSCs as supportive cells may promote repair via secreting cytokines [43,44]. It may be concluded that differentiation and replacement of MSCs at an extremely early stage are both important for the subsequent self-repair and -renewal of functional cells. Direct differentiation of MSCs, as an important mechanism of injured kidney repair, warrants further investigation.
Footnotes
Acknowledgments
Supported by grants from the “863 Projects” of Ministry of Science and Technology of P.R. China (No. 2006AA02A109, 2006AA02A115); National Natural Science Foundation of China (No. 30830052P); Beijing Ministry of Science and Technology (No. D07050701350701); and Cheung Kong Scholars program.
Author Disclosure Statement
No competing financial interests exist.