
Abstract
Human umbilical cord blood (CB) could be an attractive source of hematopoietic repopulating cells for clinical stem cell therapy because of its accessibility and low propensity for unwanted immune reaction against the host. However, CB recipients suffer from severely delayed and often chronically deficient platelet recovery of unknown cause. Here we show that human short-term repopulating cells (STRCs), which predominantly carry early hematopoietic reconstitution after transplantation, display an intrinsically fixed differentiation program in vivo that changes during ontogeny. Compared to adult sources of hematopoietic cells, CB myeloid-restricted STRC-M showed a markedly reduced megakaryocytic and erythroid cell output in the quantitative xenotransplantation of human short-term hematopoiesis in NOD/SCID-β2m–/– mice. This output in vivo was not altered by pre-treating CB cells before transplantation with growth factors that effectively stimulate megakaryocytopoiesis in vitro. Moreover, injecting mice with granulocyte colony-stimulating factor did not affect the differentiation of human STRC. These findings demonstrate that the differentiation capacity of human STRCs is developmentally regulated by mechanisms inaccessible to currently available hematopoietic growth factors, and explain why thrombopoiesis is deficient in clinical CB transplantation.
Introduction
C
Recent studies in mice have revealed changes in the differentiation programs that hematopoietic stem cells activate during ontogeny [14 –17]. Fetal hematopoietic stem cell regulation in mice abruptly switches to an adult program 3–4 weeks after birth [17]. Longitudinal analysis of fetal hematopoietic stem cell transplantation in adult mice has demonstrated that this switch is intrinsically controlled.
Based on this data, we hypothesized that ontogeny-related changes of cellular programming might also extend into more differentiated STRC. Our data demonstrate that cell source-specific programming underlies the low propensity of CB STRC to generate megakaryocytic cells. The fixed ratio of cellular lineage differentiation is independent of the cellular environment and delays platelet reconstitution after clinical CB transplantation.
Materials and Methods
Human cells
All human material was acquired with informed consent and used according to approved institutional protocols. Apheresis blood was collected from 6 adult patients after mobilization chemotherapy followed by daily injections of human G-CSF according to protocols approved by the Institutional Review Board of the Freiburg University School of Medicine. CB was obtained from full-term delivered infants after informed parental consent.
Cell purification
Mononuclear cells (MNC) were separated by Ficoll-Hypaque (PAA Laboratories, Pasching, Germany) density gradient centrifugation. CD34+ cells were isolated from MNC by passing the cells over a magnetic antibody separation column twice following the manufacturer's instructions (Miltenyi Biotech, Bergisch Gladbach, Germany) (CB = 79% ± 4% pure; mPB = 93% ± 5% pure). These cells were then cryopreserved in DMEM (Gibco, Paisley, UK) containing 30% FCS (StemCell Technologies, Vancouver, BC, Canada) and 15% DMSO (Sigma, Steinheim, Germany).
Xenotransplantation of hematopoietic cells
NOD/LtSz-Prkdcscid-β2microglobulin–/– (NOD/SCID-β2m–/–) mice were bred and housed at a specific pathogen-free animal facility according to all applicable laws and regulations following approval by the institution's animal care and ethical committee. At 8–10 weeks of age, they were irradiated with 325 cGy of 137Cs X-rays and subsequently injected intravenously with human test cells (as indicated) together with 106 irradiated (1,500 cGy) human mPB MNC as carrier cells. Serial bone marrow aspirations of the transplanted NOD/SCID-β2m–/– mice were done to determine human engraftment. Groups of mice were injected subcutaneously with G-CSF (Filgrastim, Neupogen, Amgen, München, Germany) at 250 μg/kg/day starting 7 days after irradiation and xenotransplantation. Treatment was interrupted for 7 days after the first BM aspiration and then continued daily until 8 weeks post-transplantation.
Liquid cell suspension cultures
CD34+ CB cells were seeded at 1.4–1.7 × 105 cells/mL in serum-free CellGro SCGM medium (CellGenix, Freiburg, Germany) supplemented with 1% penicillin/streptomycin (BioWhittaker, Verviers, Belgium) and the following growth factors: 15 U/mL erythropoietin (EPO), 10 ng/mL FLT3-Ligand (FL3), 50 ng/mL thrombopoietin (TPO), 10 ng/mL SCF, 5 ng/mL IL-3, 10 ng/mL IL-6, and 10 ng/ml IL-11 for megakaryocytic-erythroid differentiation or 100 ng/mL FL3, 20 ng/mL SCF, 20 ng/mL IL-3, 20 ng/mL IL-6, 20 ng/mL GM-CSF (R&D Systems, Wiesbaden-Nordenstadt, Germany), and 20 ng/mL G-CSF (Filgrastim, Neupogen, Amgen, München, Germany) for granulomonocytic differentiation. The cultures were incubated at 37°C in a humidified atmosphere containing 5% carbon dioxide for up to 21 days with the addition of fresh medium plus growth factors after 4, 8, and 14 days of incubation to double the volume at each of these time points.
Progenitor assays
Progenitor assays were performed as previously described [18]. In order to quantify the megakaryocytic colony-forming cells (CFCs), test cells were plated in a collagen-based media (MegaCult; StemCell Technologies) following the manufacturer's instructions. After incubation for 10–12 days at 37°C in a humidified atmosphere with 5% CO2 in air, the colonies were dried and fixed with methanol/acetone and stained with a primary antibody to the human MK-specific antigen glycoprotein (GP) IIb/IIIa linked to a secondary biotinylated antibody–alkaline phosphatase avidin conjugate.
Flow cytometry
For immunophenotyping, hematopoietic cells were washed with HBSS plus 2% FBS (HF; StemCell Technologies) and incubated for 30 min at 4°C with FITC-conjugated anti-CD34, anti-glycophorin A, or anti-CD15/66b or PE-conjugated anti-CD33 or anti-CD41. After being washed in HF and 1 μg/mL propidium iodide (Molecular Probes Inc., Eugene, OR), the cells were resuspended in HF and analyzed. To phenotype human cells present in the mouse BM aspirates, cells were stained with antibodies to human CD45 and CD71, CD19 and CD20, CD15 and CD66b, CD33, CD34, CD41, and glycophorin A cells and assessed as previously described [10]. All antibodies were purchased from Becton Dickinson except for anti-glycophorin A (Immunotools, Friesoythe, Germany). Acquisition and analysis were performed by Cell Quest software (Becton Dickinson, Franklin Lakes, NJ). Positive cells were defined as those exhibiting a level of fluorescence that exceeded 99.99% of the level reached by PI−; cells with irrelevant isotype-matched control antibodies labeled with the same fluorochromes. Mice with >10 CD45/71+ human cells per 20,000 PI−; cells were scored as positive. The combined presence of CD15+, CD33+, CD41+, and/or glycophorin A+ cells 3 weeks post-transplant was used to measure STRC-M activity whereas the presence of both myeloid and lymphoid (CD19/20+) human cells was used to define STRC-ML activity after 5–8 weeks.
Statistical analyses
The frequencies of human repopulating cells were calculated using Poisson statistics and the method of maximum likelihood (L-calc software; StemCell Technologies). Comparisons were made using the Student's t-test. In brief, the frequencies of STRC-M and STRC-ML were determined by transplanting limited numbers (2 × 105 to 4 × 106) of CD34+ mPB or CB cells into NOD/SCID-β2m–/– mice in 6 independent experiments. Human cell engraftment in the BM of transplanted animals was analyzed by flow cytometry at 2, 3, and 8 weeks after transplantation. Number of human hematopoietic human cells was standardized for 105 injected cells.
Results
Ontogenetic changes in cell lineage output of human short-term repopulating cells
We transplanted limiting dilution sets of CD34+ cells from pooled human CB samples into sublethally irradiated NOD/SCID-β2m–/– mice and measured the numbers of human erythroid (glycophorin A+), megakaryocytic (CD41+), granulopoietic (CD15/66b+), and B-lymphoid (CD19/20+) progeny generated in the bone marrow (BM) of the mice 3 and 5–8 weeks later (Fig. 1A). Most of the human cells detected in mice reconstituted after 3 weeks with limiting numbers of CB cells were myeloid (erythroid, granulopoietic, or megakaryopoietic) as indicative of their origin from STRC-M (Fig. 1B). The frequency of CB STRC-M was 1 in 34,000 ± 9,200 CD34+ CB cells, 5.4-fold lower than in adult BM [10]. Further, this lower number of CB STRC-M contained a still much smaller proportion of cell clones that were either megakaryocytic or erythroid. These were 4.7-fold and 3.8-fold lower, respectively, in CB STRC-M compared to clones derived from adult BM STRC-M. CB STRC-M clones were larger in NOD/SCID-β2m–/– mouse bone marrow as compared to adult BM [10] (P < 0.05; Fig. 1D).
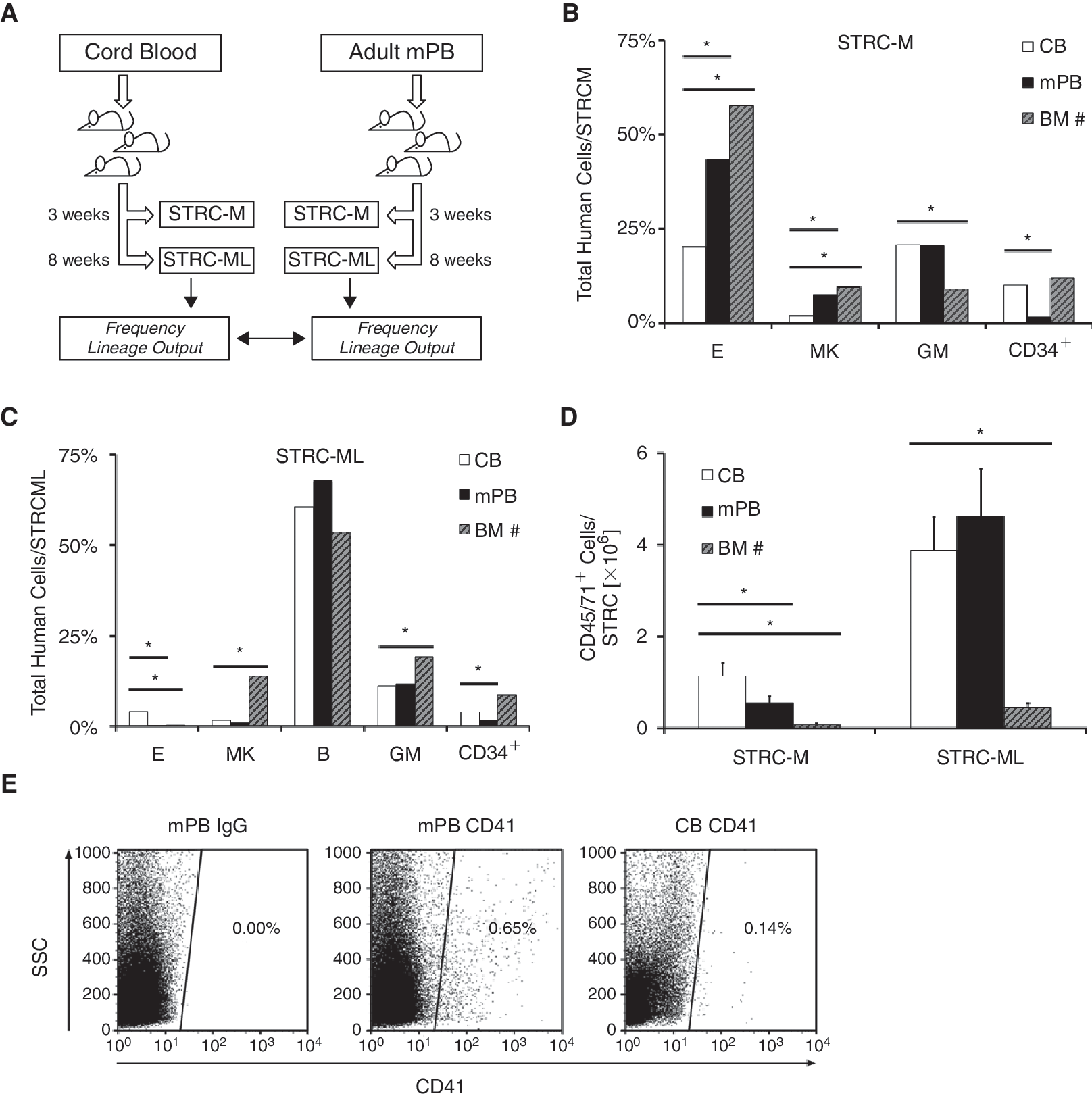
The lineage differentiation of human short repopulating hematopoietic cells (STRC) changes during ontogeny. (
After 5–8 weeks, both human lymphoid and human myeloid cell engraftments were consistently detected demonstrating their origin from CB STRC-ML (Fig. 1C). The frequency of these STRC-ML clones in CD34+ CB cells was 1 in 30,000 ± 9,800, 3.6-fold lower than in BM. The average size of CB STRC-ML clones was significantly larger than adult BM [10] STRC-ML clones (8.5-fold; Fig. 1D). STRC-ML in both CB and adult BM generated clones of similar lineage composition including the megakaryocytic cell output, except for a larger proportion of erythroid cells produced by CB STRC-ML (P < 0.05).
To determine whether these differences in the properties of STRC from CB and adult BM [10] were related to their developmental status or to their mobilized status outside the marrow cavity, we determined STRC in adult CD34+ cells isolated from granulocyte colony-stimulating factor (G-CSF)-mobilized human peripheral blood (mPB) harvests. After transplanting 82 NOD/SCID-β2m–/– mice with CD34+ mPB cells, we found that the relative representation of each of the lineages amongst the progeny obtained from adult mPB and BM was very similar, and strikingly different from the lineage preferences inherent to CB STRC (Fig. 1B–1E). Taken together, these findings indicate that not only the number but also the proliferation and differentiation programs of human STRC change during ontogeny.
Ex vivo growth factor stimulation does not change the lineage differentiation
Recent reports have suggested that ex vivo growth factor stimulation applied to partial or complete CB transplants may alleviate PB cytopenia post-transplant, especially thrombocytopenia [19,20]. We therefore determined whether the low megakaryocytic cell output of CB STRC can be influenced by prior exposure to growth factor stimulation with erythropoietin, FLT3-ligand, thrombopoietin, stem cell factor, IL-3, IL-6, and IL-11 (Fig. 2A), a combination capable of expanding megakaryopoiesis in vitro [21 –24]. Growth factor stimulation resulted in a 44 ± 12-fold expansion of megakaryocytic cells after 14 days in culture, a 880 ± 200-fold increase in the number of erythroid cells and a 15 ± 5-fold increase in the number of megakaryocytic CFCs after 8 days (P < 0.05; Fig. 2B). The number and lineage output of STRC before and after being cultured for up to 21 days were then examined in 220 NOD/SCID-β2m–/– mice. STRC-M and STRC-ML numbers dropped during the first 4 days in culture and after a transient 2-fold increase in STRC-ML after 8 days, both types of STRC stabilized at 15%–30% of the input value after 21 days in culture (Fig. 2C). Strikingly, neither the size nor the lineage content of the clones subsequently generated by the cultured cells were significantly different from those present in the freshly isolated CD34+ population (Fig. 2D and 2E). We found no evidence of megakaryocytic-erythroid-restricted clones in oligoclonally engrafted mice.
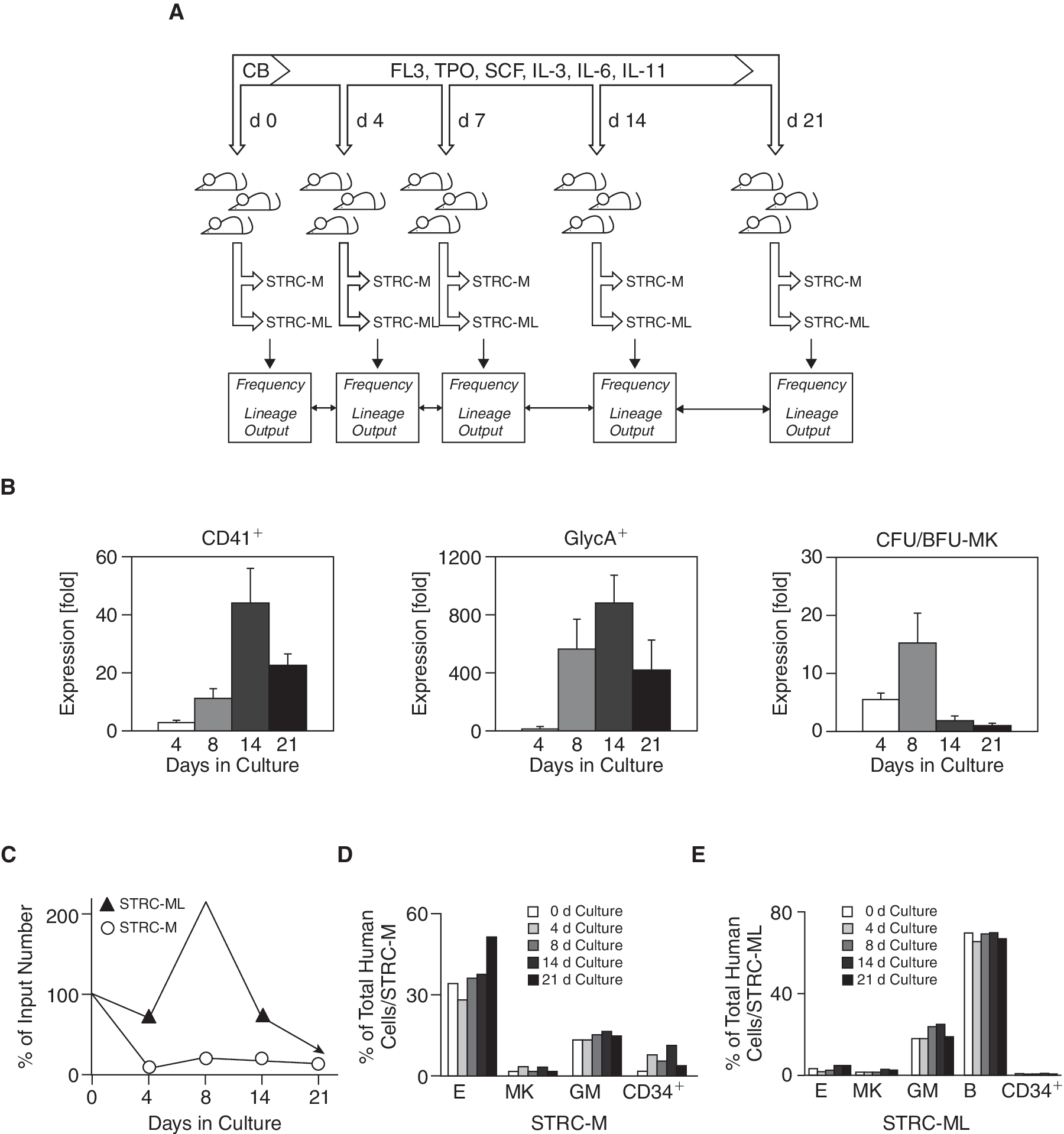
Unchanged lineage differentiation of cord blood (CB) short-term repopulating cells (STRCs) after directed megakaryocytic-erythroid ex vivo differentiation. (
To extend this investigation to other growth factors, CB cells were incubated under conditions that favor the production of monocytes and granulocytes [25 –28] (Fig. 3A). Again, progenitor cells with a defined differentiation capability were efficiently generated in the ex vivo cultures, including CD33+ and CD15/66b+ cells, morphologically recognizable granulocytes, and monocytes (data not shown) as well as granulopoietic CFC (Fig. 3B; n = 3). In vivo, however, the number of CB STRC-M declined 1.3-fold by day 4 and 14-fold by day 14 in culture, as determined by limiting dilution analysis of the repopulation of 126 NOD/SCID-β2m–/– mice (Fig. 3C). STRC-ML numbers increased 2-fold during the first 8 days in culture, and then decreased to 70% of the input value by day 14. Although these cultures generated a large number of lineage-restricted progenitor cells, the differentiation programs of both types of STRC remained unchanged (Fig. 3D and 3E). Moreover, there was no evidence for an additional repopulating cell type with other lineage restrictions [5,6]. The gross differences in STRC and CFC population changes in vitro indicate that colony-forming progenitor cells do not contribute to the activity of STRC, emphasizing the value of the NOD/SCID-β2m–/– mouse model for investigating early post-transplant hematopoiesis. The inherent differentiation program of STRC is ontogenetically regulated but not affected by ex vivo exposure to several potent growth factors known to stimulate human hematopoiesis in vitro and in vivo.
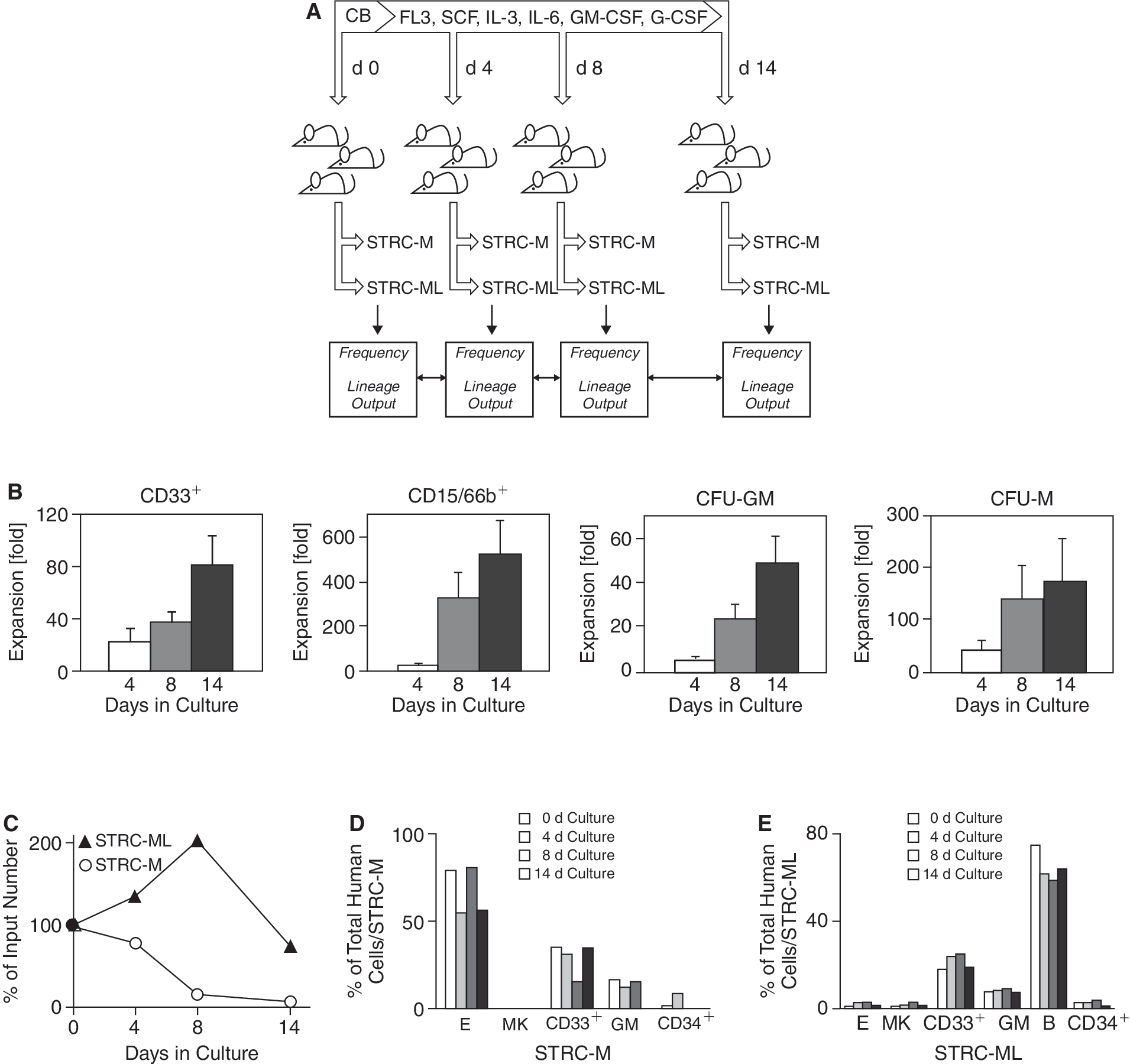
Directed granulomonocytic differentiation ex vivo does not affect the differentiation program of human short-term repopulating cells (STRCs). (
Unchanged differentiation of STRC after in vivo growth factor stimulation
In order to determine whether growth factors might alter STRC differentiation if provided to the cells in vivo within their bone marrow niche, we administered 14 daily injections of 250 μg/kg/day of G-CSF to 49 human CB transplanted NOD/SCID-β2m–/– mice before the analysis of STRC-M and 28 daily injections before the analysis of STRC-ML activity. Analysis of the numbers and types of human cells present in the BM of these mice showed that even extended G-CSF treatment in vivo did not significantly alter the cell output activity of the injected STRC (data not shown).
Discussion
We demonstrate that the differentiation program of STRC undergoes drastic changes during ontogeny. Compared to adult STRC-M, CB STRC-M have a >4-fold lower ability to produce megakaryocytic cells. In our previous study on human BM STRC, a different selection method for the purification of CD34+ cells and a slightly lower irradiation dose (325 instead of 350 cGy) were used [10]. Nevertheless, the blood cell lineage contribution of adult STRC purified from the bone marrow or from the mobilized blood of different donors was similar strongly arguing for an ontogeny-related and not a purification or patient-dependent change in the differentiation program of STRC-M. Together with the reduced frequency of STRC in CB and the about 10-fold lower number of total CD34+ cells transplanted, the capacity of a CB transplant for megakaryopoiesis is >2log lower as compared to an adult BM transplant. The higher proliferative potential of human CB STRC-M as compared to adult BM STRC-M may compensate in part for the small CD34+ transplant size but clearly fails to compensate for the largely deficient megakaryopoiesis after clinical CB transplantation. This data explains why patients transplanted with CB cells show delayed platelet recovery long after the neutrophil count has recovered [2,3].
Our data indicate that primitive human hematopoietic cells are endowed with pre-set differentiation programs (road-maps) [29] that change during ontogeny. Several observations underscore that the differentiation and proliferation program of STRC is intrinsically regulated. The differences in cell lineage output were independently maintained in a standardized adult murine microenvironment. As potential helper cell effects of co-transplanted non-clonogenic CD34−;cells cannot be excluded, we co-transplanted a standardized number of irradiated MNCs in all experiments. Redirection of differentiation by hematopoietic growth factors in vitro or in vivo had no detectable impact on the hematopoietic cell lineage output or clone size of STRC. In mice a fetal stem cell program that is characterized by high self-renewal activity and accentuated myeloid cell output switches to an adult program at about 3 weeks after birth and then remains unchanged [17]. The exact timing of developmental switches in hematopoietic stem and progenitor programs has not yet been assessed in humans because serial hematopoietic stem cell samples are not available. Our results clearly indicate that such changes occur at a post-natal time point. Changes in the rate of telomere loss in peripheral blood granulocytes during early childhood point out that switches in proliferative activity may not be restricted to STRC but may also occur at the more primitive hematopoietic stem cell level [30].
Conclusion
An intrinsically fixed differentiation program of human STRC leads to the delayed platelet reconstitution after CB transplantation. The lack of any effect of growth factor treatment on STRC differentiation in vivo regardless of the factors used or the duration of the treatment argues strongly against the likelihood that currently available extrinsic manipulations will be of significant utility to generate CB transplants with more rapid platelet reconstituting activity, just like the thrombocytopenic marrow environment in the recipient fails to reprogram these cells. Recent reports on the successful generation of inducible pluripotent stem cells (iPS cells) from adult fibroblasts have established the feasibility of a fundamental reprogramming of adult cell fate [31,32]. Our results underscore that genetic reprogramming of inherent differentiation preferences in CB progenitors will be required to overcome extended thrombocytopenia after CB transplantation.
Footnotes
Acknowledgments
We thank Marie Follow and Klaus Geiger for cell sorting. We also gratefully acknowledge helpful discussions and support from Dr. C. Peters and Dr. R. Mertelsmann. This work was supported by the Bundesministerium für Bildung und Forschung, grant 01KV9907, by the Deutsche Forschungsgemeinschaft DFG, grants KA 976/5-1, KA 976/5-2, Ka976/5-3, and Ka976/5-4 and by the Deutsche Krebshilfe grants 10-1860-GI I and 107217.
Author Disclosure Statement
The authors have no potential conflicts of interest to declare.