
Abstract
Four commercially available serum-free and defined culture media tested on 2 human embryonic stem cell (hESC) lines were all found to support undifferentiated growth for >10 continuous passages. For hESC cultured with defined StemPro and mTeSR1 media, the cells were maintained feeder-free on culture dishes coated with extracellular matrices (ECMs) with no requirement of feeder-conditioned media (CM). For xeno-free serum replacer (XSR), HEScGRO™, and KnockOut media, mitotically inactivated human foreskin feeders (hFFs) were required for hESC growth. Under the different media conditions, cells continued to exhibit alkaline phosphatase activity and expressed undifferentiated hESC markers Oct-4, stage-specific embryonic antigens 4 (SSEA-4), and Tra-1-60. In addition, hESC maintained the expression of podocalyxin-like protein-1 (PODXL), an antigen recently reported in another study to be present in undifferentiated hESC. The cytotoxic antibody mAb 84 binds via PODXL expressed on hESC surface and kills >90% of hESC within 45 min of incubation. When these cells were spontaneously differentiated to form embryoid bodies, derivatives representing the 3 germ layers were obtained. Injection of hESC into animal models resulted in teratomas and the formation of tissue types indicative of ectodermal, endodermal, and mesodermal lineages were observed. Our data also suggested that StemPro and mTeSR1 media were more optimal for hESC proliferation compared to cells grown on CM because the growth rate of hESC increased by 30%–40%, higher split ratio was thus required for weekly passaging. This is advantageous for the large-scale cultivation of hESC required in clinical applications.
Introduction
V
Although CM produced by human feeders in KO-SR medium supplemented with fibroblast growth factor 2 (FGF-2) was found to maintain hESC in its pluripotent state, it is not completely defined [4] and evidence of animal proteins have been reported [7]. Nonhuman molecules such as sialic acid Neu5Gc was found to be incorporated and expressed by hESC cultured in the presence of animal-derived products [8,9]. As humans are likely to have antibodies developed against this molecule, exposure of hESC derivatives may cause a rejection resulting in killing of the transplanted cells. Hence, the elimination of nonconforming grade media components both in derivation and long-term culture of pluripotent hESC is necessary to expand their usefulness for differentiation into specific lineages for clinical and therapeutic applications [10,11].
For feeder-free culture of hESC, plastic dishes are commonly coated with an extracellular matrix (ECM) before use. Although unclear at present, there were suggestions that the adhesion molecules present in the complex ECM in combination with growth factors secreted by feeder cells play a synergistic role in supporting the growth of hESC [12,13]. One of the more commonly used ECM is matrigel, which is derived from a mouse sarcoma cell line. Other defined sources of ECM such as recombinant or human-sourced fibronectin, vitronectin, and laminin have been reported to be potential alternatives for matrigel [14 –16]. Other groups attempted to culture hESC in chemically defined as well as xeno-free conditions. Although some works with defined media have claimed success in maintaining hESC in their pluripotent state [15,16], others have contradicted [17]. Further efforts are extended to derive and culture new human foreskin feeders (hFFs) and hESC lines in xeno-free conditions [18 –20] and it has been proven that the maintenance of pluripotent hESC in such a platform for ξ30 passages was achievable [21]. Further efforts were made to generate clinically compliant hESC in anticipation for future applications [22].
Although the interest to develop the most optimal culture conditions continue, we investigated here several commercially available defined media namely HESGRO, KO medium with xeno-free serum replacer (XSR), StemPro, and mTeSR1. HEScGRO™ and XSR required the use of hFFs [2], whereas StemPro and mTeSR1 maintained hESC in a defined, serum- and feeder-free condition. Altogether, we have successfully cultured multiple hESC lines using the 4 media conditions for ξ10 passages and found that the cells retain hallmarks of pluripotency in both in vitro and in vivo characterization assays.
Materials and Methods
Cell culture
Preparation of feeder cells
The hFFs HS68 obtained from ATCC (CRL1635) was maintained as described by Choo et al. [2]. Briefly, the feeders were passaged and expanded with fibroblast culture medium (FDMEM) made up of 90% DMEM high glucose (Invitrogen Gibco, Carlsbad, CA), 10% fetal bovine serum (Hyclone), 2 mM
Culture of hESC
MEF was equilibrated with KO-SR medium made up of 85% KO DMEM, 15% KO-SR, 1 mM
Human ESC was passaged every 7 days. Briefly, the cells were washed once with 1× phosphate-buffered saline (PBS; Invitrogen Gibco) before being enzymatically treated for ∼3–7 min (Table 1).After neutralization with the respective culture media, the cells were dislodged and broken into small clumps or single cells before seeding onto either new ECM-coated or hFF plates at a split ratio of 1:3 to 1:8. The cells were incubated at 37°C in 5% CO2 incubator.
Conditioned media (CM) is produced from MEF and used for the feeding of human embryonic stem cell (hESC).
Human foreskin fi broblast (hFF) line utilized was HS68 (CRL1635) from ATCC.
Abbreviations: KO-SR, KnockOut serum replacer; XSR, xeno-free serum replacer; MEF, mouse embryonic fibroblast; hFF, human foreskin feeder; ECM, extracellular matrix.
Expression of pluripotent hEsC markers
Flow cytometry analysis (FACS)
The expression levels of the pluripotent markers Oct-4, stage-specific embryonic antigens 4 (SSEA-4), and Tra-1-60 in hESC populations were assessed by immunofluorescence using flow cytometry. Cells were harvested as single-cell suspensions using 0.25% trypsin-EDTA (Invitrogen Gibco), fixed, and permeabilized (Invitrogen Caltag, Carlsbad, CA) before being incubated with a mouse monoclonal antibody to Oct-4 (1:20; Santa Cruz Biotechnology, Inc., Santa Cruz, CA), SSEA-4 (1:1, DHSB, Jowa City, Jowa), and Tra-1-60 (1:50; Chemicon). Cells were then washed with 1% BSA/PBS, and incubated in the dark with goat α-mouse antibody either FITC or PE-conjugated (DAKO, Tokyo, Japan) at 1:500 dilution. After incubation, the cells were washed and resuspended in 1% BSA/PBS for analysis on a FACScan (FACScalibur, Becton Dickinson). All incubations were performed at room temperature for 15 min. For the negative control, cells were stained with the appropriate isotype control.
Staining of hESC for markers
Immunocytochemical staining of hESC was carried out by incubating the cells with fixative Reagent A (Invitrogen Caltag) for 1 h, before blocking with 3% BSA/PBS for another hour. After washing with 0.1% Triton/PBS, the cells were incubated with antibodies to Oct-4 (Santa Cruz), SSEA-4 (DHSB), and Tra-1-60 (Chemicon) for 1 h. Detection of antibodies bound to the pluripotent markers was visualized using FITC- or PE-conjugated goat α-mouse antibody (1:500; DAKO).
To assay for alkaline phosphatase activity, hESCs were stained using Vector Red Alkaline Phosphatase Substrate Kit I (Vector Laboratories, Burlingame, CA) according to the manufacturer's protocol. Briefly, hESCs were washed once in PBS followed by incubation in the dark with Vector Red substrate working solution for 45 min at room temperature. Images of the stained cells were taken with an Olympus fluorescent microscope IX70.
Cytotoxic-killing assay with monoclonal antibody (mAb) 84
Cytotoxicity of mAb 84 on hESC was carried out using propidium iodide (PI) exclusion assays and flow cytometry according to the method discussed by Choo et al. (2008) [26]. Briefly, single-cell suspensions at 2 × 105 cells per 10 μL volume in 1% BSA/PBS were incubated with 5 μg purified mAb 84 in 200 μL 1% BSA/PBS for 45 min on ice. After which, cells were washed and resuspended in 1% BSA/PBS and 1.25 mg/mL PI for analysis. As a negative control, cells were incubated with the isotype control, mAb 85.
Growth rates and doubling time
Single-cell suspensions of HES-3 were harvested daily (days 1–6 after seeding) following treatment with 0.25% trypsin-EDTA (Invitrogen, Carlsbad, CA). The total cell number was determined using a nucleocounter (Chemometec, Allerød, Denmark). Graphs of total cell number versus time were plotted to estimate the specific growth rate of cells during the exponential growth phase. From this, the doubling time (td) was calculated using the following equation, td = ln2/μ, where μ is the specific growth rate (1/h).
Karyotypic stability
Karyotyping analysis was performed by the Cytogenetics Laboratories at the Department of Obstetrics and Gynaecology, KK Women's and Children's Hospital. Cell samples were incubated with BrdU/colcemid (reagent from hospital) for 16 h in 37°C, 5% CO2 incubator.
In vitro and in vivo differentiation assay
SCID mice model and teratoma analysis
Human ESCs were harvested by collagenase (Sigma) treatment and ∼4–5 × 106 cells were injected with a sterile 22G needle into the rear leg muscle of 4-week-old female SCID mice. Mice that developed tumors ∼9–10 weeks after injection were humanely killed and the tumors were dissected and fixed in 10% formalin. Tumors were embedded in paraffin, sectioned, and examined histologically after hematoxylin and eosin staining.
Embryoid bodiesformation and RT-PCR analysis
To determine the pluripotency of HES-3 cultured with defined and serum-free media, RT-PCR analysis was carried out. Embryoid bodies (EBs) cultured in suspension for 7 days were replated as single cells onto gelatinized plates. Total RNA was isolated from replated 21-day-old EBs using NucleoSpin RNA II Kit (Macherey Nagel) and quantified using Nanofrop ND-1000 spectrophotometer (Nanodrop Technologies, Wilmington, DE). Reverse transcription reactions were performed with 1μg total RNA using oligo-dT primers and M-MLV reverse transcriptase (Promega, Madison, WI). The PCR was carried out using primers specific to differentiation markers from the 3 germ layers. Endodermal lineage was represented by Alpha-feto protein (AFP), amylase, and GATA6, ectodermal lineage by neurofilament heavy chain (NFH) and keratin-15, while mesodermal lineage was indicated by heart and neural crest derivatives 1 (HAND1) and Msh homeo box homolog 1 (MSX1). As a control, total RNA was harvested from undifferentiated HES-3 and interrogated with primers to Oct-4 as well as differentiated markers. The primer sequences are shown in Table 2. The cycling parameters used for amplification were 30 cycles of 95°C for 30 s, 55°C for 30 s and 72°C for 30 s. This was followed by a final extension at 72°C for 10 min. The amplified products were visualized on 1% agarose gels and stained with ethidium bromide.
Results
Adaptation of hESC to defined and serum-free media culture condition
Interestingly, all hESC lines tested did not require gradual adaptation when transferred from MEF CM culture directly to the serum-free or defined media platforms [29]. These cells maintained their undifferentiated phenotype in defined media for ξ10 continuous passages (Fig. 1). Unlike other platforms, cells cultured in StemPro medium were dissociated using accutase and thus did not require neutralization with media. The enzymatic treatment resulted in single cells or smaller clumps as compared to cells passaged with collagenase. Initially, a higher seeding density (eg, 3 passages at 1:2 split ratio) was required for passages to ensure higher cell viability of undifferentiated hESC during inoculation. The cells grew as a monolayer instead of typical clumps that form colonies with distinct edges on matrigel-coated surface, while still maintaining high nucleus-to-cytoplasmic ratio (Fig. 2A). Similarly, both HES-2 and HES-3 were also successfully transferred and cultured on fibronectin-coated dishes (data not shown).
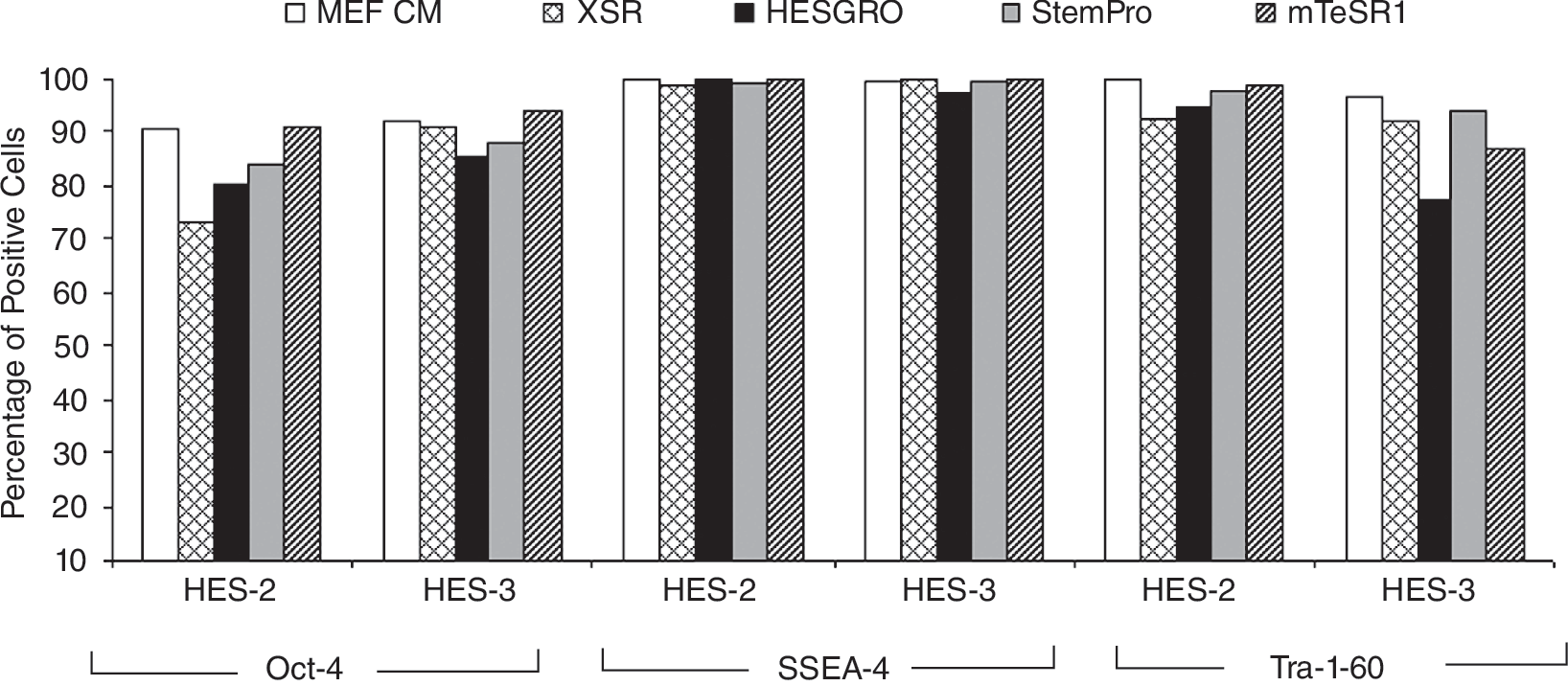
Expression of undifferentiated markers by human embryonic stem cell (hESC) cultured in various media. Undifferentiated markers Oct-4, SSEA-4, and Tra-1-60 were detected for HES-2 and HES-3 cultured on StemPro (passage 38, p38), mTeSR1 (p26), XSR (p10), and HESGRO (p10). More than 70% of hESC population remained undifferentiated when cultured under these conditions.
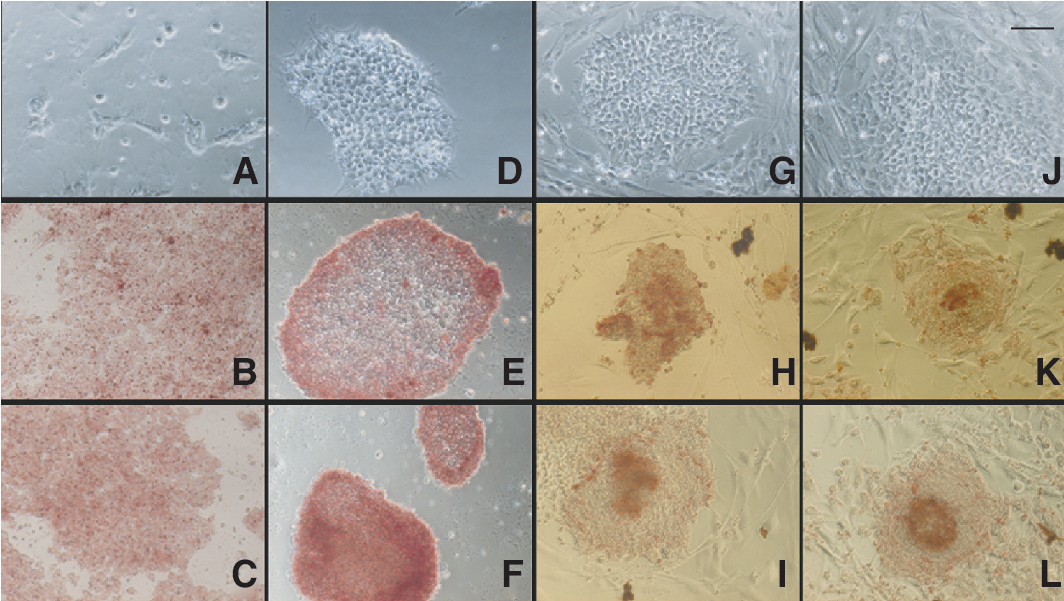
Colony morphology and alkaline phosphatase staining of hESC cultured in various defined and serum-free media. Images of HES-3 cultured in (
For cells cultured with mTeSR1 medium, colonies formed appeared to be thin but maintained tight cell-cell contact on matrigel-coated dishes (Fig. 2D). Unlike hESC cultured with MEF CM, cystic-looking regions, which indicated differentiated cell populations, were rarely observed. However, hESC cultured on mTeSR1 medium was not able to readily adapt to fibronectin-coated surfaces due to poor adhesion. This could be due to the lack of other matrix components that aid in cell adherence after passaging to allow more robust growth [25]. After several passages, a higher split ratio (1:8–1:10) is achievable for hESC cultured in StemPro and mTeSR1 media. This is advantageous as it allows greater expansion of hESC in a shorter time span.
The recommended use for XSR and HESGRO media for hESC culture required the presence of feeder cells. In our study, hFFs previously found to support the growth of undifferentiated hESC lines were used [2]. Similarly, the transfer of the hESC lines to xeno-free media platforms did not require an adaptation period. The clumps of hESC grew as compact colonies after seeding on a monolayer of hFF cells. The outward growth of hESC after seeding onto hFF can be seen distinctly several days later, with the feeder cells at the periphery of the clumps (Fig. 2G and 2J). Attempts were made to transfer cells to feeder-free culture platform (matrigel) supplemented with hFF CM. Although cells adhered and proliferated, there was a significant increase in the differentiated population of hESC and loss of pluripotent marker expression (data not shown).
Growth rates and doubling time
For StemPro and mTeSR1 media, the growth rates of hESC obtained were higher compared to cells cultured with MEF CM. Our results indicated a shorter doubling time for cells cultured in defined media (Table 3)and thus requiring less time between passaging and reaching confluency. The typical split ratio of 1:8 to 1:10 was routinely used for weekly passaging. This seemed to suggest an improvement in culture condition to render more efficient cell growth. It was suggested that a shortened G1 cell-cycle phase was linked to self-renewal of hESC and hence pluripotency [30]. To ascertain whether StemPro and mTeSR1 media provided hESC with more optimized growth conditions, HES-3 cells cultured in these 2 media for ξ10 passages were transferred back to MEF CM and maintained a further 5 passages before monitoring the growth rates. It was apparent that the growth rate of HES-3 cells became comparable to the parental culture platform when readapted to MEF CM (Table 3).
HES-3 cultured on both StemPro and mTeSR1 media were readapted to MEF conditioned media (CM) for 5 passages.
Abbreviations: KO-SR, KnockOut serum replacer; XSR, xeno-free serum replacer.
For hESC cultured on hFF and supplemented with either HESGRO or XSR media, the accelerated growth rate was not observed (Table 3).The split ratio was maintained at 1:4 unlike the faster growing feeder-free hESC cultures on defined media. The growth rate of hESC cultured on hFF with KO-SR medium is comparable to that previously reported by Choo et al. [2] where hESC was cocultured on various hFFs.
Expression of pluripotent markers
The expression of pluripotent markers Oct-4, SSEA-4, and Tra-1-60 were routinely monitored for hESC cultured in defined and serum-free media. HES-2 and HES-3 cultured on matrigel for 38 (StemPro) and 26 (mTeSR1) passages, respectively, expressed high levels of pluripotent markers. When cultured on fibronectin-coated dishes for 12 passages with StemPro media, ∼90% of the cell population for both HES-2 and HES-3 expressed high levels of these markers (Supplementary Fig. 1; Supplementary materials are available online at
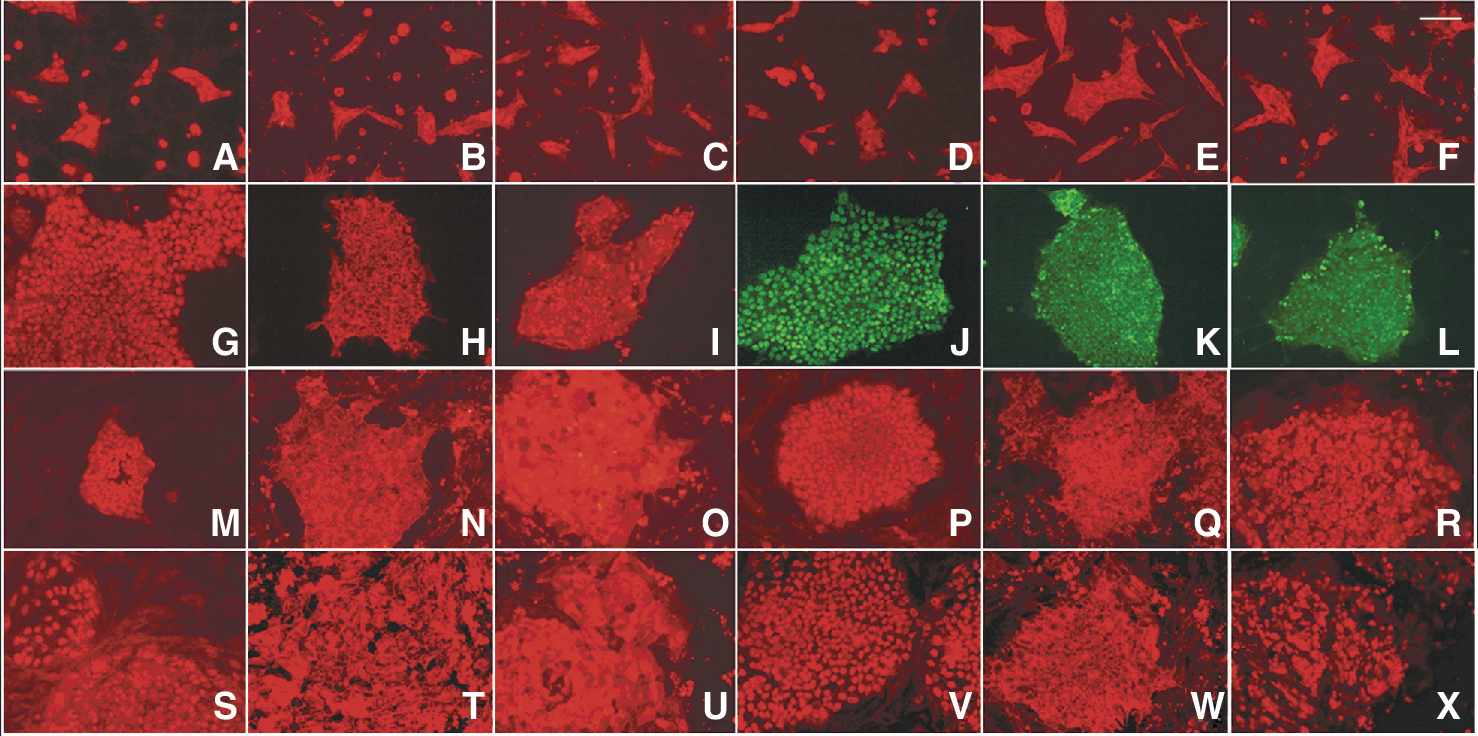
Immunostaining of hESC cultured in serum-free and defined media. Undifferentiated markers Oct-4, SSEA-4, and Tra-1-60 positively stained for HES-2 cultured on (
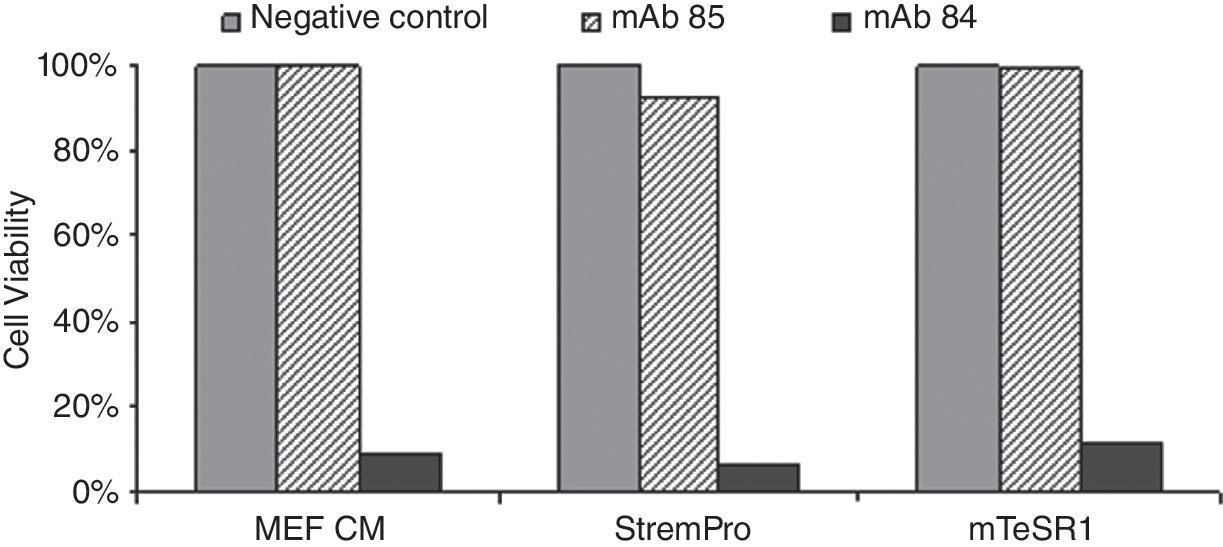
Population of undifferentiated hESC killed by mAb 84. Binding of mAb 84 to PODXL on undifferentiated hESC maintained in StemPro and mTeSR1 media. Percentage of viable hESC after treatment with mAb 84 was <12%.
Chromosomal stability
Cells were routinely monitored for abnormalities after every 10–15 passages. After long-term passaging, hESC maintained in serum-free and defined media was found to be stable maintaining a normal diploid karyotype (Fig. 5). HES-2 and HES-3 cultured on both matrigel- and fibronectin-coated dishes with StemPro media were karyotypically stable after 24 passages. Similar results were obtained for both cell lines cultured after 19, 9, and 37 passages with mTeSR1, XSR, and HESGRO media, respectively (data not shown).
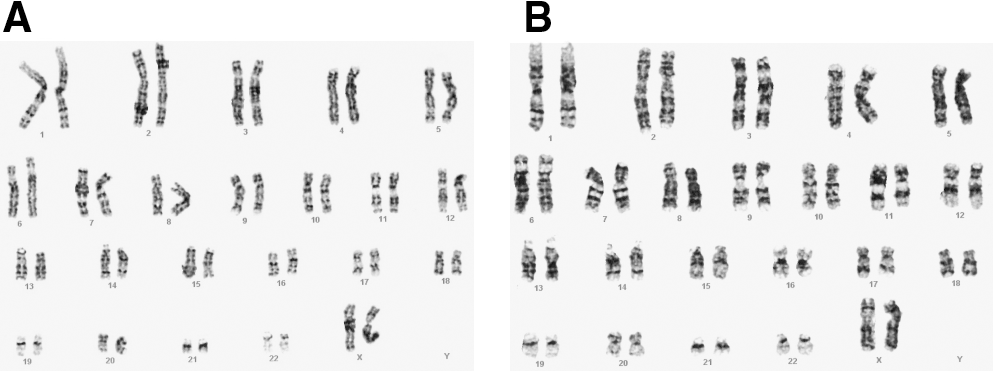
Stable karyotype of hESC cultured in various serum-free and defined media. Normal diploid karyotype (46XX) was obtained for both (
In vivo and in vitro differentiation
To evaluate the differentiation potential, HES-3 cultured in StemPro, mTeSR1, HESGRO, and XSR media were spontaneously differentiated to form embryoid bodies in vitro at passages 31, 29, 23, and 14, respectively. The various derivatives representing the 3 germ layers were found to be present in the differentiated cells and detected by RT-PCR. Alpha-feto protein (AFP), amylase, and GATA6 representing endodermal lineages, neurofilament heavy chain (NFH), and keratin-15 representing ectodermal lineages, heart and neural crest derivatives 1 (HAND1), and Msh homeo box homolog 1 (MSX1) indicative of mesodermal lineages were detected (Fig. 6). Inversely, these genes were not detected in the undifferentiated HES-3 control while Oct-4, consistent with this phenotype was detected after long-term culture in StemPro and mTeSR1 media at passage 29 (Supplementary Fig. 2). The pluripotent state of HES-3 maintained in the various media was confirmed in vivo following the observation of tumor formation 8–9 weeks after injection of cells into SCID mice. The mice were sacrificed and the teratomas were processed and analyzed. The appearances of tissues representing all 3 germ layers—ectoderm (neural epithelium), mesoderm (bone), and endoderm (gut-like epithelium)—were observed (Fig. 7).
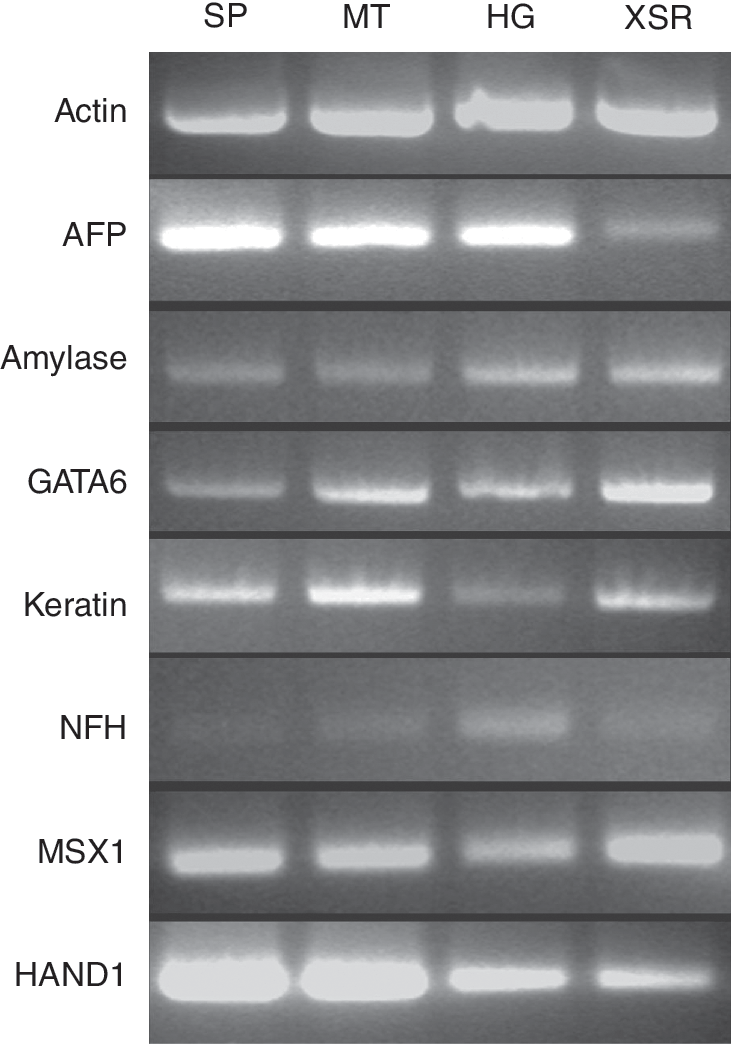
Detection of differentiated markers for 3 germ lineages. RT-PCR analysis for expression of endoderm markers: AFP, amylase, and GATA6; ectoderm markers: NFH and keratin-15; and mesoderm markers: HAND1 and MSX1 in embryoid bodies generated from HES-3 cultured for 31, 29, 14, and 23 passages in StemPro (SP), mTeSR1 (MT), XSR, and HESGRO (HG), respectively.
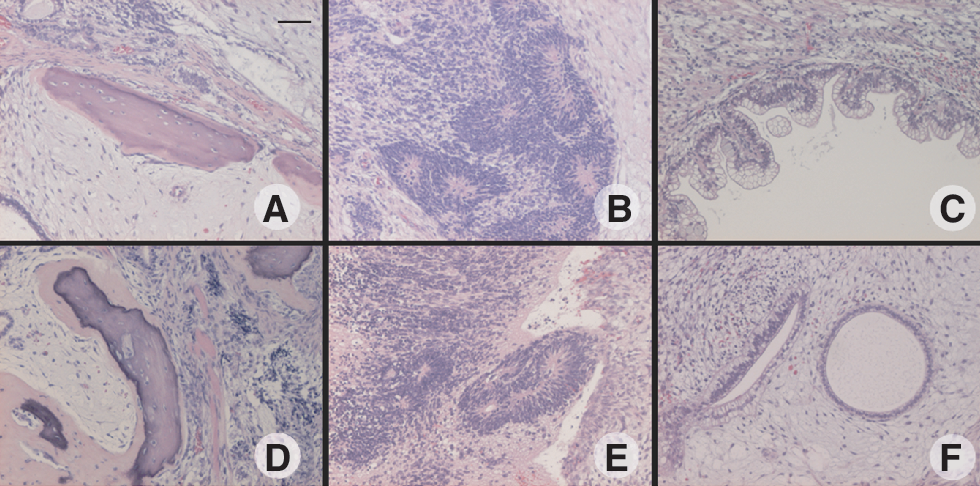
Examination of the pluripotency of hESC in vivo. HES-3 was injected intramuscularly into the rear limbs of SCID mice. Tissues representing 3 germ lineages were observed, including bone (mesoderm), neuroepithelium (ectoderm), and gut epithelium (endoderm). Teratomas resulting from HES-3 cultured in (
Discussion
To facilitate the expansion of hESC for differentiation into various phenotypic lineages for clinical uses, the development of a culture process that is defined and free of animal products is necessary. Adopting a xeno-free environment for the culture of hESC has its challenges. However, several studies have recently demonstrated that hESC can be derived and cultured successfully in a xeno-free environment [16,18,21]. Improvements to culture conditions have also been made to achieve a more chemically defined hESC culture method, from the substitution of mouse feeders to human feeders through to the setup of feeder-free systems and the use of materials that are xeno-free. The choice of serum-free and defined media is preferred as it offers ease of substituting products that do not conform to standards, as the components are known. In this study, we demonstrated successfully the culture of hESC using 4 different commercially available serum-free and defined media with the cells remaining karyotypically stable (Fig. 5) and undifferentiated (Figs. 1,3,4) for at least 10 continuous passages.
StemPro and mTeSR1 media did not require feeder cells and hence xeno-contamination can be avoided. Although BSA was added in StemPro medium, the carrier protein can be replaced with a human or recombinant source. However, the use of human serum as an alternative source of proteins may still be subjected to preparation variability as it is also complex and undefined and may result in different growth [21]. Hence, the advantage of using serum-free media allowed better consistency and reproducibility for cell culture. Although matrigel is still used in the majority of feeder-free culturing of hESC, we found that the cells can be transferred onto non-animal source ECM such as fibronectin or vitronectin without any loss in pluripotent marker expression (data not shown). This is coherent with other published literature that found either fibronectin or combinations of various human-sourced ECM can support the growth of hESC in a defined media [21,31]. Separately, we tested 2 other xeno-free media, HESGRO and XSR, which required hFF cells. Similarly, various groups have also demonstrated the derivation of feeders in xeno-free environments [19 –21]. Hence, it is possible to achieve xeno-free culture of hESC in the presence of feeders.
Typically, the transfer of hESC to a new culture condition requires an adaptation period [29,32]. Interestingly in this study, hESCs that were transferred directly to the various new media seemed to grow well with minimal adaptation, while remaining undifferentiated after ξ10 passages (Figs. 1 and 3) with no chromosomal abnormalities (Fig. 5). Furthermore, after incubation with mAb 84, a monoclonal antibody that binds to PODXL present on undifferentiated hESC [26], we found that ξ90% of cells were killed (Fig. 4). Both in vitro and in vivo differentiation assays successfully produced spontaneously differentiated hESC, as indicated by the presence of derivatives from the 3 germ lineages (Figs. 6 and 7).
Commonly, hESC is passaged as clumps using enzymes such as collagenase IV. Bajpai et al. [33] reported that hESC could be propagated efficiently as single cells, which will be advantageous for large-scale expansion of the cells as well as downstream applications. We explored passaging hESC as single cells using accutase. The cells cultured with StemPro media grew as a monolayer on ECM-coated dishes unlike the outward colony growth typical of hESC morphology (Fig. 2A–2C). Despite the differences in cell morphology, the cells remained karyotypically stable and retained the expression of undifferentiated markers for ξ10 passages (Figs. 1 and 3A–3F).
We observed that hESC cultured on StemPro and mTeSR1 media reached confluency in less than a week, hence requiring higher split ratio as compared to hESC supplemented with MEF CM. Similar observations were made by Becker et al. [30] who found that hESC exhibited a unique shortened G1 cell-cycle phase thus only requiring an estimated 16 h per cycle. It was further suggested that the competency of hESC for self-renewal was linked to pluripotency. In order to determine whether these media were more optimized with growth factors, cells were monitored for their growth rates over a week. Our data suggested that the hESC adapted to defined media had a shorter doubling time compared to parallel hESC supplemented with MEF CM (Table 3).Furthermore, when hESC cultured with StemPro and mTeSR1 media were transferred back to MEF CM and maintained for 5 passages, the doubling time reversed back to ∼28h. This observation was less apparent for hESC cultured on hFF cells that were fed with xeno-free HESGRO and XSR. We have attempted to transfer these cells to ECM-coated plates but were unsuccessful due to increase in spontaneous differentiation (data not shown). The need of hFF for hESC cultured with HESGRO and XSR suggest that additional supportive growth factors produced by the feeder cells were required to maintain undifferentiated hESC growth [12,13]. More recently, we have also evaluated a modified XSR formulation, which contains a growth factor cocktail intended to replace the requirement of feeders. Preliminary data from HES-3 feeder-free cultures on matrigel continue to maintain the expression of pluripotent markers (Oct-4, PODXL, and Tra-1-60) at passage 10 (data not shown). Effort to adapt the hESC onto human fibronectin is currently on-going.
While researchers are devising extensive methods for the establishment of xeno-free and clinical-grade hESC lines [21,22,25], we have cultured hESC on various potential defined and serum-free media that will be compatible for such uses. Although similar approaches were used to culture hESC, the results may not be reproducible as discussed by Rajala et al. for the use TeSR1 media [17,25]. This could be due to differences in the increasing number of hESC lines being derived and hence culture requirements. However, it is clear that the direction for the cultivation of hESC should be avoidance of animal products and conformance to clinical grade in order for future use in therapeutic applications [34].
Author Disclosure Statement
No competing financial interests exist.
Footnotes
Acknowledgments
This work was supported by the Biomedical Research Council of A*STAR (Agency for Science, Technology and Research), Singapore. Special thanks to Dr. Niki Wong and Fong Wey Jia for reviewing the manuscript.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.