
Abstract
In human physiology and animal models, bone marrow mesenchymal stem cells (MSCs) exert an immunosuppressive role in both in vitro and in vivo experiments. However, cellular and molecular mechanisms involved in this process are not clear and remain largely elusive. Several studies have suggested the implication of cell–cell contacts or soluble factors including transforming growth factor-b1 (TGF-b1), interleukin-10 (IL-10), indoleamine 2,3-dioxygenase (IDO), or human leukocyte antigen-G (HLA-G). Here, we show that both Galectin-1 and Semaphorin-3A (Sema-3A), 2 soluble factors capable to inhibit T-cell proliferation through neuropilin-1 (NP-1) binding, are highly expressed by MSCs and may account for their known suppressive activities. Furthermore, MSCs suppressive functions are completely reverted by soluble recombinant NP-1, the main receptor of both Galectin-1 and Sema-3A. Similar results were obtained by using blocking antibodies against Galectin-1 or Sema-3A. Taken together, these results demonstrate the critical role of Galectin-1 and Sema-3A in MSCs functions and may open new perspectives in the understanding and treatment of various immune and neoplastic disorders.
Introduction
B
Several studies reported the immunosuppressive properties of MSCs in vitro and in vivo. Recently, MSCs have been shown to exert suppressive effect on T-cell proliferation [4,5], on NK cell activities [6,7], on B-cell functions [8,9], and on dendritic cell differentiation and maturation [10]. While this immunosuppression is well established, the mechanism of action is still unclear and controversial. Several studies have previously demonstrated the implication in this process of cell–cell contacts and soluble factors such as cytokines (TGF-b, IL-10), nitric oxide, or PGE2/IDO [11]. Nevertheless, the precise mechanism and the factors which govern these MSCs immune regulatory functions remain largely elusive and are still a matter of debate.
A recent study has shed light on the constitutive presence of Galectin-1 in MSCs [12]; however, its function in this model is elusive. Galectin-1 is a homodimeric galactose-binding lectin with a single carbohydrate-recognition domain that is able to selectively bind neuropilin-1 (NP-1) [13]. NP-1, a neuronal receptor, is constitutively expressed on T-cell surface [14,15] and is also involved in the regulation of T-cell proliferation through its main ligand Semaphorin-3A (Sema-3A) [16] that arrests T cells in G0/G1 cell cycle phase [17]. Given the immunomodulatory properties of both Galectin-1 and Sema-3A, we have tested the probable impact of NP-1 ligands in MSCs functions.
Here, we show that both Galectin-1 and Sema-3A are highly expressed and secreted by MSCs. Moreover, addition of both recombinant immunosuppressive factors (Galectin-1 or Sema-3A) inhibits T-cell proliferation. Furthermore, anti-Sema-3A or anti-Galectin-1-blocking antibodies and recombinant NP-1 neutralized MSCs immunosuppressive properties on T cells. Therefore, Galectin-1 and Sema-3A seem to be crucial immunoregulatory proteins involved in MSCs functions.
Materials and Methods
Cell culture setup and expansion
Experimental procedures, with human BM, have been approved by the Necker and Saint Louis Hospital Ethical Committees for human research, in accordance with the European Union guidelines and the declaration of Helsinki. MSCs were isolated from washed filters used during BM graft processing for allogenic BM transplantation. MSCs were obtained and cultured as previously described [18]. Cells were seeded at 104 cells/cm2 and fed every 2 or 3 days until confluence. Adipogenic and osteogenic MSCs differentiation assays were performed in specific media as described [18].
Flow cytometry and western blot analysis
MSCs were stained with PE, FITC, or APC-conjugated anti-CD90, CD34, CD45, CD166, CD44, CD29, CD13, and CD105 monoclonal antibodies or with appropriate controls and analyzed using a FACSCalibur (Becton Dickinson, France). Western blot analysis and immunoprecipitation were performed using Sema-3A and Galectin-1-specific antibodies detected by HRP-conjugated antibody and revealed with ECL kit (Pierce, France). Protein loading was controlled using GAPDH antibody (Tebu Santa-Cruz, France).
Mixed leukocyte reaction (MLR)
MSCs were irradiated (25 Gy) and seeded with freshly isolated peripheral blood mononuclear cells (PBMCs) as previously described [18] at concentrations ranging from 0.1% to 20% of responder T cells/wells (ie, 100–20,000 MSCs). For blocking experiment, MSCs were seeded at the concentration of 20% of responder T cells and were incubated with neutralizing TGF-b1, IL-10, Galectin-1, and Sema-3A antibodies (10–40 μg/mL) or with recombinant neuropilin-1 (Fc-NP-1) (2–5 μg/mL; R&D systems, France).
Statistical analysis
The results were analyzed by independent sample 2-tailed and unpaired Student's t-test, and were presented as means ± standard error deviation.
Results and Discussion
In this study, we have investigated the mechanisms involved in the immunosuppressive functions of MSCs. In accordance with previous report [19], BM MSCs used here did not express hematopoietic antigens, such as CD14, CD34, and CD45, or endothelial markers like CD31. Expanded MSCs expressed CD13, CD73, CD90, CD29 (b1-integrin), CD44 and CD166 (adhesion molecules), and CD105 (endoglin receptor) antigens (Fig. 1A). Furthermore, MSCs differentiated into adipogenic (defined by the accumulation of lipid-rich vacuoles) and osteogenic (characterized by ALP activity and deposition of a calcium-rich mineralized extracellular matrix) lineages [1] (Fig. 1B and 1C, respectively).
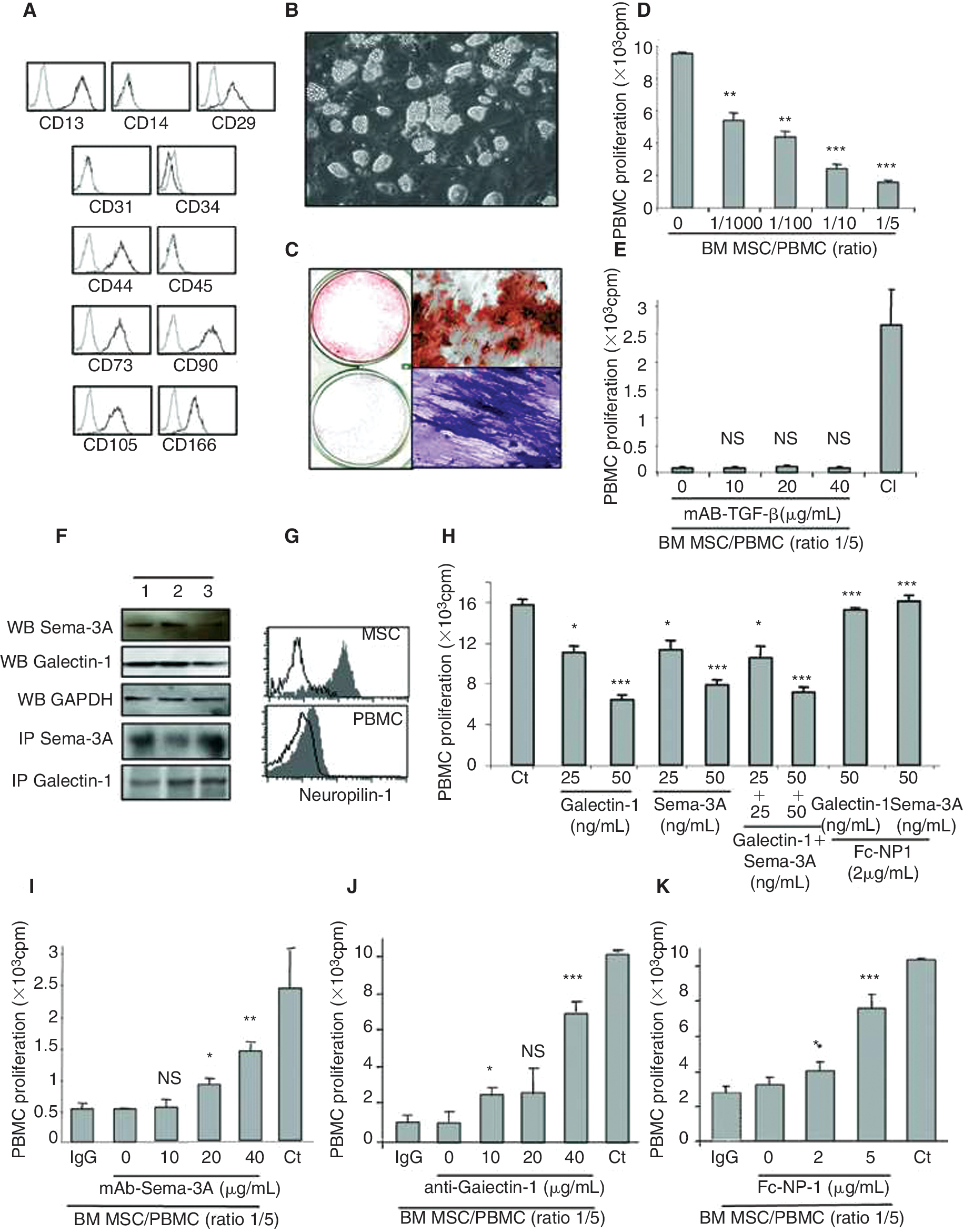
Human bone marrow (BM) mesenchymal stem cells (MSCs) immunosuppression depends on soluble neuronal receptor (NP)-1 ligands. (
As MSCs from BM, synovium, or adipose tissues are able to inhibit T-cell proliferation in a mixed-lymphocyte reaction, we tested their immunosuppressive activity on allogenic T-cell proliferation at different MSCs:T cells ratios (Fig. 1D), in accordance with the literature [5,20 –22]. The immunosuppression of T-cell proliferation by MSCs was more efficient at higher MSCs:T cells ratio (Fig. 1D). Many studies have pointed out the role of soluble factors such as TGF-b1 in the mechanism by which MSCs inhibit T-cell proliferation [5]. Consistently with previous reports [23], in our hands, no significant reversion of MSCs immunosuppressive effect was observed on T-cell proliferation in the presence of TGF-b1-blocking antibodies (Fig. 1E), while MSCs secreted TGF-b1 (1,787 ± 652 pg/mL). Recent work also pointed to the presence and the potential role of Galectin-1 in MSCs functions [12]. However, no experimental results have proved yet this concept. Since Galectin-1 as well as Sema-3A [15,16] are both immunosuppressive proteins acting through NP-1 [13] expressed on T cells, we investigated whether these soluble factors were involved in the MSCs immunosuppressive properties. We first demonstrated the expression of both molecules and their receptor neuropilin-1 in MSCs (Fig. 1F and 1G, respectively). We next assessed whether Sema-3A and Galectin-1 were directly involved in T-cell immunosuppression mediated by MSCs, since both ligands are capable to specifically inhibit T-cell proliferation (Fig. 1H). To this aim, MSCs and PBMCs were co-cultured in the presence of neutralizing Galectin-1 or Sema-3A antibodies. The blockade of Sema-3A and Galectin-1 secreted by MSCs restored the proliferation of PBMCs in a dose-dependent manner (Fig. 1I and 1J, respectively). To reinforce this result, we also used recombinant soluble NP-1 that bound and blocked both Sema-3A and Galectin-1 proteins, allowing the MSCs immunosuppressive activity neutralization (Fig. 1K). Taken together, these results demonstrated the implication of both Galectin-1 and Sema-3A in the immunosuppressive properties of MSCs.
Sema-3A is a member of a large protein family, which is phylogenetically conserved secreted protein originally implicated in axon guidance [24]. These proteins have been extensively studied in the nervous system, but accumulated evidence showed that they have important functions on nonneuronal cell system including angiogenesis and tumor biology. More recently, we and others have shown that they are also involved in the complex system of interactions between epithelial cells and thymocytes within the thymus as well as between T and DC cells in the periphery [14,15,25]. More consistently, Galectin-1 is overexpressed in regulatory T cells and its neutralization significantly reduced the inhibitory effects of human and mouse CD4+CD25+ T regulatory cells [26]. These results suggest that Galectin-1 is a key effector of the regulation mediated by immunosuppressive cells. Here, we show for the first time that MSCs also expressed another immunosuppressive protein, Sema-3A. Taken together, expression of both Sema-3A and Galectin-1 by MSCs may explain their immunosuppressive activities, in accordance with MSCs molecular signature [27]. Finally, these results also provided new insights in the physiopathology and treatment of various immune and neoplastic disorders.
Footnotes
Acknowledgments
S.L. was supported in part by a grant from the French Agence Nationale de la Recherche (no. ANR05PRIB01103). This work was supported in part by grants from the Leducq Foundation (Cardiac Progenitors Transatlantic Alliance network 04CVD04), Pôle de Compétitivité Meditech Santé Ingenis (no. 052906053), Ligue Contre le Cancer, Arreca, Canceropole Ile de France, Fondation pour la recherche médicale, Fondation de France and INCa. We wish to thank Dr. F. Audat and Dr. J.A. Ribeil (Department of Biotherapy, Hospital Necker-Enfants Malades) for providing us with blood samples. We wish to thank Dr. A. Shirvan and Dr. A. Barzilai for providing us anti-Sema-3A.
Author Disclosure Statement
No competing financial interests exist.