
Abstract
Pancreatic islet β-cell replenishment can be driven by epithelial cells from exocrine pancreas via epithelial–mesenchymal transition (EMT) and the reverse process MET, while specified pancreatic mesenchymal cells control islet cell development and maintenance. The role of human islet-derived precursor cells (hIPCs) in regeneration and support of endocrine islets is under investigation. Here, we analyzed hIPCs as to their immunophenotype, multilineage differentiation capacity, and gene profiling, in comparison to human bone marrow-derived mesenchymal stem cells (hBM-MSCs). hIPCs and hBM-MSCs display a common mesenchymal character and express lineage-specific marker genes upon induction toward pancreatic endocrine and mesenchymal pathways of differentiation. hIPCs can go further along endocrine pathways while lacking some core mesenchymal differentiation attributes. Significance analysis of microarray (SAM) from 5 hBM-MSC and 3 hIPC donors mirrored such differences. Candidate gene cluster analysis disclosed differential expression of key lineage regulators, indicated a HoxA gene-associated positional memory in hIPCs and hBM-MSCs, and showed as well a clear transition state from mesenchyme to epithelium or vice versa in hIPCs. Our findings raise new research platforms to further clarify the potential of hIPCs to undergo complete MET thus contributing to islet cell replenishment, maintenance, and function.
Introduction
T
Epithelial–mesenchymal transition (EMT) is a fundamental developmental process when ectoderm develops into mesoderm. The latter gives rise to organ development like kidney and testes as well as to ultimate mesenchymal tissues like bone, cartilage, and muscle [7,8]. In adult organisms, EMT occurs as a means of tissue repair and regeneration. It is proposed that islet regeneration requires transient EMT/MET of epithelial precursors, which migrate toward islet structures to replenish the endocrine pancreas [1]. This procedure of extensive differentiation plasticity is also important in other regeneration procedures, in cancer metastasis and fibrotic diseases [8,9].
Human islet-derived precursor cells (hIPCs) can be obtained from explanted human islets. Initially recognized as islet progenitor cells, this population has been under extensive investigation as to its mesenchymal or epithelial provenience and its multilineage differentiation capacity [10 –15]. Recent results obtained from lineage tracing studies in both rodents and humans are controversial regarding the primary origin of these cells [11,14]. Very similar to reports on human and rodent bone marrow-derived mesenchymal stem cells (BM-MSC) [16,17], hIPCs express mesenchymal marker genes and have been reported to be inducible into insulin-secreting cells in vitro and to improve glucose homeostasis in vivo.
Diabetes mellitus is due to deficient insulin signaling, caused either by insulin resistance or by autoimmune ⊠-cell destruction. The individual and economic burden of this disease is remarkable and the incidence is steadily increasing, strongly due to the rising incidence of obesity [18]. Cell-based therapy for type 1 diabetes has become a main issue of investigation. Islet transplantation is an effective but limited strategy with respect to availability of donor organs. Extensive investigation on cellular targets for β-cell regeneration or replacement has demonstrated that hIPCs besides other stem/progenitor cells isolated from different adult tissues (eg, neural cells, hepatocytes, umbilical cord, and adipose tissue-derived cells) can be directed toward insulin cell types in vitro and in animal models. However, none of these procedures has yet achieved the cure of experimental diabetes [19].
In the present study, by comparing the multilineage differentiation capacity and genome-wide signatures of hBM-MSCs with hIPCs, we intended to disclose major key factors in this cell population that may be related to its role in endocrine β-cell regeneration, differentiation, and maintenance.
Materials and Methods
Cell culture
All media, fetal calf serum (FCS), and supplements were obtained from PAA (Linz, Austria).
Human pancreatic islets were isolated at the Giessen Islet Isolation and Transplantation Centre (Germany) from human pancreases (n = 3) obtained from brain-dead multiorgan donors after obtaining legal consent and approval from the Local Ethics Committee, according to a modified semi-automated digestion–filtration method [20,21]. hIPCs were obtained from freshly isolated human islets of 3 different donors as previously described [22] and expanded in RPMI 1640 medium supplemented with 10% fetal calf serum (FCS), 1 U/mL penicillin (P), 100 μg/mL streptomycin (S), 50 μM β-mercaptoethanol (Sigma, Schnelldorf, Germany), 1 mM sodium pyruvate, 10 mM HEPES, 2 mM
hBM-MSCs were harvested from bone marrow of femoral heads from 5 otherwise healthy patients (n = 5) undergoing total hip arthroplasty. Informed consent was obtained from each patient and experiments were performed upon approval of the Local Ethics Committee. Whole bone marrow cells were initially seeded at a density of 6.6 × 105 per cm2 to obtain hBM-MSC by plastic adherence. After 2 to 3 days of cultivation, nonadherent cells were removed and adherent cells were washed twice with phosphate-buffered saline (PBS). Cells were cultivated and expanded in DMEM/Ham's F-12 (1:1) medium supplemented with 10% fetal calf serum (FCS), 1 U/mL P, 100 μg/mL S, and 50 μg/mL
Flow cytometry
hIPCs (at passages 3, 4, 6, 12) and hBM-MSCs (at passages 1–2) were detached with PBS–0.5% EDTA and resuspended in PBS–1% bovine serum albumin (BSA) in a round bottom polystyrene tube and labeled with fluorescein isothiocyanate (FITC)- or phycoerythrin (PE)-conjugated monoclonal antibodies according to the manufacturer's instructions. The antibodies used were: anti-human CD45-PE, CD14-PE, CD34-PE, CD-HLA-DR-FITC, CD13-PE, CD29-FITC, CD44-FITC, CD54-FITC (ImmunoTools, Friesoythe, Germany), CD73-PE, CD90-PE (BD Biosciences, Heidelberg, Germany), and CD105-FITC (Biotrend, Köln, Germany). Cells labeled with IgG1-FITC/IgG2a PE-conjugated antibody (BD Biosciences) served as negative control. Prior to FACS analysis, cells were washed twice and resuspended in 100 μL of PBS–1% BSA solution. Flow cytometry analysis (10,000 events) was performed using a BD FACScan and CellQuest-software (both BD Biosciences, Heidelberg, Germany).
Mesenchymal differentiation of hIPCs and hBM-MSCs
For adipogenic and osteogenic differentiation, hIPCs and hBM-MSCs were seeded at a density of 3 × 104 cells per cm2 and incubated in differentiation medium for 14 days (adipogenic) and 28 days (osteogenic) as previously described [23]. Cells maintained in expansion medium served as negative controls.
For chondrogenic differentiation, a cell culture system of high-density pellets was applied. To obtain chondrospheres, 2.5 × 105 cells were resuspended in chondrogenic differentiation medium, which contained 10 ng/mL TGFb1 (R&D Systems, Wiesbaden, Germany), centrifuged at 250g, and cultured in tubes for up to 21 days as previously described [23]. Pellets maintained in chondrogenic medium without addition of TGFb1 served as negative controls.
Pancreatic endocrine differentiation of hIPCs and hBM-MSCs
For endocrine differentiation, hIPCs and hBM-MSCs were seeded in ultra low-attachment 6-well plates (Corning, Schiphol-Rijk, Netherlands) and cultured for 4 days under specific endocrine conditions (cocktail medium), consisting of serum-free DMEM/F12 containing 17.5 mM glucose, 10 mM nicotinamide, 2 nM activin-A, 10 nM exendin-4, 100 pM hepatocyte growth factor (Sigma-Aldrich GmbH, Schnelldorf, Germany), 1% bovine serum albumin (BSA), 2 mM
RNA isolation and RT-PCR
Total RNA was obtained from differentiated and undifferentiated hIPCs and hBM-MSCs from 3 and 5 donors, respectively, using the NucleoSpin RNA II kit (Macherey-Nagel, Düren, Germany,
Real-time PCR
cDNA samples were investigated for mRNA levels by qPCR using Light Cycler 2.0 (Roche) and Q-PCR SYBR Green Capillary Mastermix (Abgene/Thermo Fisher Scientific). Concentrations of mRNA levels in samples were calculated from external standard curves generated for each specific primer pair (dilution series: 1:1; 1:10; 1:100; 1:1,000, and 1:10,000) by Light Cycler Software 3.5 (Roche). Every run with samples included an internal standard (1:1,000) from the same cDNA sample as used for generation of standard curves to normalize reaction efficiencies. Standard curves were generated from triplicates, while samples and internal standards were analyzed in duplicates. Benchtested qPCR primer pairs for human/HPRT/(housekeeping gene) (Quantitect Hs_HPRT1), human/MAF/(Quantitect Hs_MAF), human/ISL1/(Quantitect Hs_ISL1), human/SOX17/(Quantitect Hs_SOX17), human/HOXA2/(Quantitect Hs_HOXA2), human/HOXA10/(Quantitect Hs_HOXA10), human/TCF21/(Quantitect Hs_TCF21) were obtained from commercial supplier (Qiagen). qPCR conditions (according to general Light Cycler instructions): 95°C for 10 min; 40 cycles: 95°C for 15 s; 55°C (manufacturer's recommendation for all Quantitect primer) for 10 s; 72°C for 20 s; followed by melting curve analysis for specificity of qPCR products.
Cytochemical analysis
Adipogenic differentiation was examined by staining of intracytoplasmic lipid vesicles with Oil Red O. Cells were fixed in 4% paraformaldehyde (PFA) and incubated in 0.3% (wt/v) Oil Red O in 60% isopropanol (Merck, Darmstadt, Germany). Cell nuclei were counterstained with hematoxylin.
Chondrogenic differentiation was visualized by staining of the sulfated proteoglycans with Alcian Blue. Chondrospheres were fixed in 4% PFA, dehydrated with graded ethanol series, and embedded in paraffin and sectioned at a thickness of 4 μm. Sections were incubated in 1% (w/v) Alcian Blue pH 1.0 and counterstained with Nuclear Fast Red.
Osteogenic differentiation was evaluated by staining for cytoplasmic alkaline phosphatase (ALP) (ALP Leucocyte Kit 86-C; Sigma-Aldrich GmbH). For detection of calcium mineralization in the extracellular matrix, monolayers were fixed with methanol and stained with Alizarin Red S (1% w/v) (Chroma-Schmidt GmbH, Stuttgart, Germany).
Immunofluorescence analysis
Rabbit anti-human C-peptide (Linco Research, St. Charles, MO), guinea-pig anti-human insulin, and mouse anti-human vimentin (clone v9) (both Dako, Hamburg, Germany) were used as primary antibodies, anti-rabbit IgG-Cy3 at 1:500, 1:500, and 1:2,000 dilutions, respectively, in blocking buffer. Anti-rabbit IgG-Cy3-conjugated and anti-guinea-pig IgG-Cy2 at 1:1,000 dilution (both Dianova, Hamburg, Germany) and anti-mouse IgG Alexa 555 (Invitrogen, Karlsruhe, Germany) at 1:2,000 were used as secondary antibodies.
hBM-MSCs and hIPCs, human pancreatic islet cryosections (8 μm), and free-floating cell clusters generated in cocktail medium were fixed in 4% PFA prior to staining, permeabilized with 0.1% Triton X-100, blocked in 5% serum, and incubated at room temperature with primary and appropriate secondary antibodies for 60 and 30 min, respectively. Cell nuclei were counterstained with DAPI. Slides were analyzed by fluorescence microscopy (Carl Zeiss, Jena, Germany) using Axiovision 4.6.1.0 software (Carl Zeiss Vision, Aalen, Germany) and laser scanning confocal microscopy (Zeiss LSM 510) with Zeiss LSM 510 software (Esslingen, Germany).
Genome-wide gene expression profiling of hIPCs and hBM-MSCs populations
Hybridization experiments were performed using Affymetrix Gene Chips HG-U133 Plus 2.0 (54,000 probe sets for 47,400 transcripts and 38,500 genes, High Wycombe, United Kingdom) and the corresponding kits according to the Affymetrix GeneChip Expression Analysis Technical Manual version 2 (
To assess differentially expressed genes between hIPC and hBM-MSC groups, predefined conditions were established according to Affimetrix Gene Chip methodology. The number of “Present” calls for a given gene had to be greater than 50% in at least one of the groups and only those genes that displayed a signal log2 ratio of lower than −;1 or greater than 1, that is, with a fold change (FC) lower than 0.5 and greater than 2 were taken into account. All other probe sets were stated as “not differentially expressed.” In order to get reliable data, the q value, that is false discovery rate had to be <10%. For heat-map creation, the program Spotfire DecisionSite for Functional Genomics 9.1.1 (Spotfire AB; TIBCO Software Inc., Göteborg, Sweden) was used.
To evaluate gene ontology clusters, further analysis was attained using GOstat (
Results
Overlapping immunophenotypes of hIPCs and hBM-MSCs
To evaluate the immunophenotype of adult hBM-MSCs and hIPC, both cell populations were compared by means of surface antigen characterization using flow cytometry analysis. Both hBM-MSCs and hIPCs were negative for the hematopoietic markers CD14, CD34, and CD45, while both expressed the mesenchymal markers CD29, CD44, CD54, CD73, CD90, and CD105. According to recently established minimal criteria, these results reveal a MSC immunophenotype in both, hBM-MSCs and hIPCs [28] (Fig. 1A).

Characterization of human islet-derived precursor cells (hIPCs) and human bone marrow-derived mesenchymal stem cells (hBM-MSCs). (
Similar mesenchymal gene marker expression in hIPCs and hBM-MSCs
The expression of mesenchymal marker genes [8] was analyzed in both cell populations. Transcripts for fibronectin (FNT), Thy1 (THY1), snail2 (SNAI2), prolyl 4-hydroxylase, alpha I subunit (P4HA1), matrix metalloproteinase 2 (MMP2), and vimentin (VIM) were detected in both hIPCs and hBM-MSCs. Although to a lesser extent, mRNA for the majority of investigated mesenchymal genes was also found in extracts of intact human pancreatic islets (hislets; Fig. 1B). At the protein level, the classical mesenchymal marker vimentin was present in the investigated cell populations as well as in cells within the human islets (Fig. 1D).
Expression of genes related to stemness and development
Development and stemness-associated gene transcripts were detected in both hIPCs and hBM-MSCs. There was similar expression in both populations of, for example, high-mobility group box 1 (HMGB1), hairy enhancer of split (HES) and Pou domain (OCT) genes, polycomb group repressor gene 1 (BMI1), as well as ATP-binding cassette group 2 (ABCG2) transcripts. In contrast, Meis homeobox and some homeobox b (HOXb) family genes showed higher expression levels in hIPCs (Fig. 1C; Supplemental Table 2; available online at
Mesenchymal differentiation potential of hIPCs is reduced as compared to hBM-MSCs
To compare mesenchymal differentiation capacities, adipogenic, chondrogenic, and osteogenic differentiation were induced in hIPCs and hBM-MSCs at the same time. After adipogenic induction, transcripts for the adipogenic markers, lipoprotein lipase (LPL), and peroxisome proliferator-activated receptor (PPARγ2) were detected in both differentiated populations (Fig. 2A). However, oil droplet formation, a later marker of adipogenesis, was found to a lesser extent in hIPCs than in hBM-MSCs (Fig. 2B). After 3 weeks under chondrogenic conditions, transcripts for specific chondrogenic markers such as collagen type II (COLII) and Aggrecan (AGN) were detected in hBM-MSCs whereas in hIPCs only the hypertrophic cartilage marker collagen type X (COLX) was expressed (Fig. 2A). Alcian Blue staining indicated the presence of sulfated proteoglycans in both hIPC- and hBM-MSC-derived cell pellets (Fig. 2B). After 4 weeks in osteogenic conditions, both hIPCs and hBM-MSCs strongly stained for alkaline phosphatase (ALP). Similar expression patterns for the osteogenic markers osteocalcin (OC), ALP, and bone sialoprotein II (BSP) were detected, the latter, however, showed a much higher expression in hBM-MSCs (Fig. 2A). The degree of in vitro mineralization of the extracellular matrix, which is an indicator of osteoblast maturation, was clearly less pronounced in hIPCs compared to hBM-MSCs as shown by Alizarin Red staining (Fig. 2B).
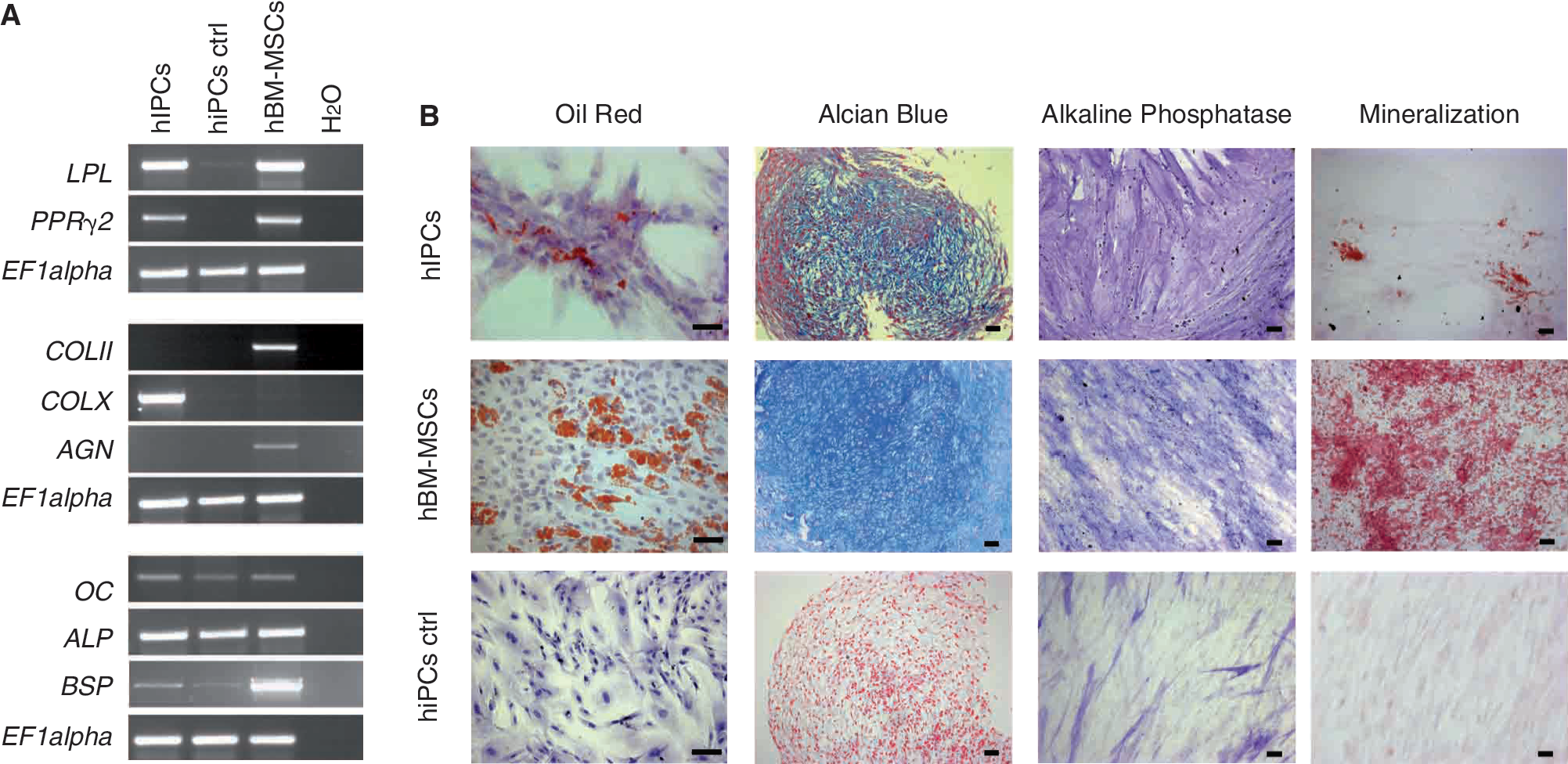
Mesenchymal differentiation potential of human islet-derived precursor cells (hIPCs) and human bone marrow-derived mesenchymal stem cells (hBM-MSCs). (
Endocrine differentiation is more effective in hIPCs than in hBM-MSCs
Both cell types were kept in specific culture conditions to evaluate their endocrine differentiation potential. Within 4 days, both cell types developed spherical cell clusters (Fig. 3A). Transcripts for the key transcription factor pancreatic-duodenal homeobox 1 (PDX1/Ipf1), as well as the early endocrine promoters of β and δ cells PAX6 and PAX4, were detected in both hIPCs and hBM-MSCs after induction of endocrine differentiation (Fig. 3B). Neurogenin 3 (NGN3), a pancreatic endocrine progenitor cell marker, which is transiently expressed during differentiation into islet-like cells, was not detected in any of the treated populations (data not shown). On the other hand, expression of specific genes involved in β-cell function such as the V-maf musculoaponeurotic fibrosarcoma oncogene homolog A (MAFA), insulin (INS), and glucose transporter type 2 (GLUT2) was observed in differentiated hIPCs but not in hBM-MSCs. Detection of mRNA for glucagon (GCG), somatostatin (SST), and P polypeptide (PP) in hIPCs suggests that some of these cells underwent differentiation steps toward various islet cell phenotypes. Synthesis of insulin protein was detected by immunostaining of hIPCs with anti-insulin and anti-C-peptide antibodies (Fig. 3C), suggesting a high level of maturation in differentiated hIPCs, which could not be observed in hBM-MSCs (data not shown).

Endocrine differentiation potential of human islet-derived precursor cells (hIPCs) and human bone marrow-derived mesenchymal stem cells (hBM-MSCs). (
Detection of epithelial and mesenchymal cells in human pancreatic islets
To further evaluate the epithelial and mesenchymal cell populations residing in the islets of Langerhans, sections of isolated human pancreatic islets were analyzed for the synthesis of insulin and vimentin by immunofluorescence. Vimentin-positive cells (red) were found among the insulinstained cells (green), which shows the existence of a mesenchymal cell population, within the pancreatic islet (Fig. 4, left panel). The detection of vimentin and insulin co-labeled cells (yellowish) in adult human islets ex vivo strongly suggests that along with mature endocrine cells and mesenchymal cells, some others are found in an epithelial–mesenchymal or mesenchymal–epithelial state of transition (Fig. 4, right panel).
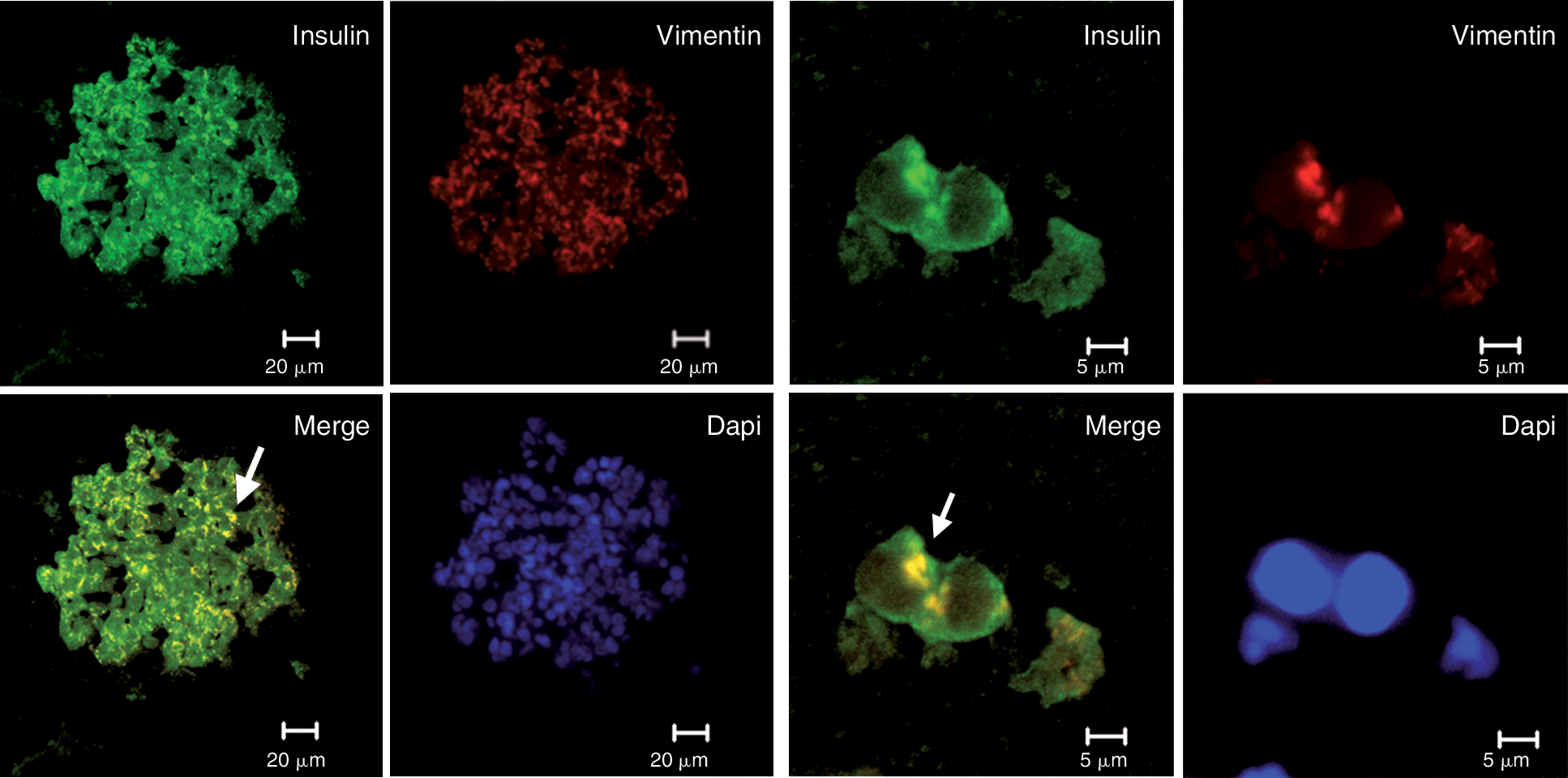
Detection of epithelial and mesenchymal cell markers in pancreatic islets. Confocal imaging of a human pancreatic islet, co-stained with insulin (green) and vimentin (red). Cell nuclei were stained with DAPI. The left panel shows insulin/vimentin-positive cells (yellow, arrow) on a section of whole pancreatic islet (bar represents 20 μm). The right panel shows single cells from the same islets labeled for insulin and vimentin (arrow) (bar represents 5 μm) (n = 3).
SAM-based genome-wide expression profiling in hIPCS and hBM-MSC
The comparative analysis of gene hybridization signals from microarrays derived from hBM-MSCs and hIPCs was carried out using significance analysis of microarrays (SAM). The 22,484 probe sets were equally expressed in hIPCs and hBM-MSCs, 1,606 were significantly higher expressed in hIPCs (FC > 2), 1,995 were significantly lower expressed in hIPCs (FC < 0.5) (Fig. 5A).

Genome-wide expression profiling in human islet-derived precursor cells (hIPCs) and human bone marrow-derived mesenchymal stem cells (hBM-MSCs). (
Functional assignment of gene ontology clusters in hIPCs and hBM-MSCs
To assign functional categories to the identified gene clusters, microarray gene expression profiles of hIPCs and hBM-MSCs were examined using gene ontology (GO) analysis. GOstat analysis in hIPCs revealed a significant enrichment of functional gene clusters related to “gland development,” “signal transduction,” and “cell differentiation.” In contrast, in hBM-MSC, ontology groups like most remarkably “skeletal development,” but also “myogenesis” and “sensory organ development,” were significantly overrepresented (Fig. 5B).
Candidate gene clusters in hIPCs and hBM-MSCs
To identify relevant genes associated with mesenchymal or epithelial determination, cell transition state, and key organ morphogenesis, a candidate gene approach was used, based on the significantly differentially expressed genes in hIPCs versus hBM-MSCs that have been obtained by SAM. The expression pattern of key genes in both populations was compared on the basis of SAM data and verified by RT-PCR. Some of these genes were quantified by qPCR (Table 1 and Fig. 6A–6C).
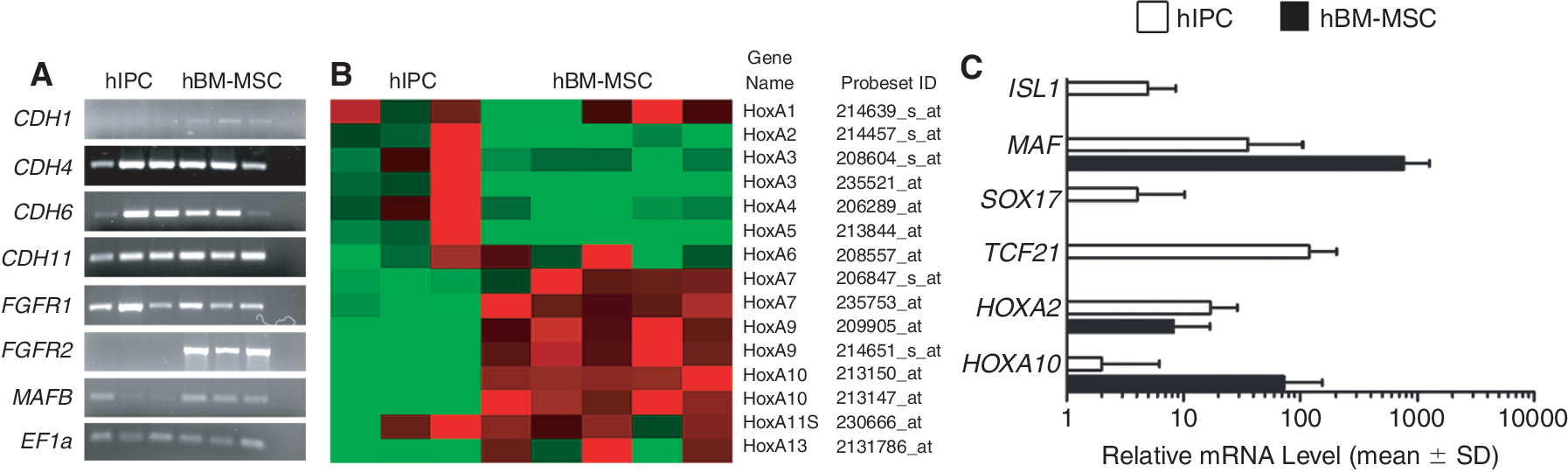
Candidate gene clusters in human islet-derived precursor cells (hIPCs) and human bone marrow-derived mesenchymal stem cells (hBM-MSCs). (
Individual preparations of hIPCs (n = 3) and hBM-MSCs (n = 5) were analyzed. Gene names, detection in hBM-MSC and hIPCs (absent: A; present: P), analyzed probe set IDs, fold changes (FC) of the expression values between hIPCs and hBM-MSCs, and the gene titles are shown. Fold changes marked with ** have highly significant q values below 0.1. When no difference between expression levels could be detected (ns: not significantly different), only the number of analyzed probe sets is given. hIPCs, human islet-derived precursor cells; hBM-MSCs, human bone marrow-derived mesenchymal stem cells.
E-Cadherin (CDH1), the classical epithelial marker was negative in hIPC albeit slightly expressed in hBM-MSC, while vimentin was strongly present in both populations (see also Fig. 1). Retinal cadherin (CDH4) was equally present in both populations according to RT-PCR analysis, in this case discongruent to SAM data. Also fetal kidney cadherin (CDH6) and osteoblast cadherin (CDH11) were equally expressed in hIPCs and hBM-MSCs. All other cadherins were either not differentially expressed or absent in either population (Table 1).
The expression pattern of E-cadherin repressor genes was very similar in hIPCs and in hBM-MSCs (Table 1)as shown by the expression of SNAI1 and SNAI2, TCF3, as well as LOXL2. In contrast, while TWIST1 (FC 0.31) and SIP1, the E-cadherin promoter silencer (FC 0.34) were lower in hIPCs, higher expression was found for TWIST2 (mean FC 2.5), the high-mobility group protein HMGA2 (mean FC 2.8), and hepatocyte growth factor (HGF) (mean FC 4.0).
Finally, we looked for mesenchymal marker genes relevant in mesoderm specification and MSC differentiation versus those that are relevant for endocrine differentiation (Table 1 and Fig. 6A and 6C). hIPCs displayed comparably low levels of fibroblast growth factor receptor 2 (FGFR2) (mean FC 0.06), which is active in MSC differentiation while mesenchyme-specific MAFK expression (FC 2.94) was relatively high. V-maf musculoaponeurotic fibrosarcoma oncogene homolog (MAF/c-MAF), which is active in chondrocyte differentiation, was low (FC 0.04) in hIPCs, as was its homolog MAFB (mean FC 0.17), which is largely expressed in developing endocrine pancreas and induces the glucagon promoter in adult α cells. This indicates that hIPCs on one hand display many mesenchymal features but their differentiation toward mature mesenchymal cells is hampered due to important signaling defects. TCF21 (Pod-1), a basic helix-loop-helix transcription factor, is highly expressed in hIPCs only (Fig. 6C). It represses terminal lineage maturation in myoblasts and is critically important for, for example, kidney organogenesis, suggesting that hIPCs may be prohibited from mesenchymal maturation [29]. The expression pattern of members of the highly conserved homeobox gene family (HOX) is different in hIPCs compared to hBM-MSCs (Fig. 6B and 6C). HoxA2, HoxA3, HoxA4, and HoxA5 are significantly higher expressed in hIPCs, while HoxA7, HoxA9, HoxA10, and HoxA13 are higher expressed in hBM-MSCs. SRY sex-determining box 17 (SOX17), a marker for definitive endoderm, was detected in hIPCs but not in hBM-MSCs (Fig. 6C, Supplemental Fig. 1; available online at
Components of the TGFb signaling pathway were in part differentially expressed in hIPCs (Table 1). While the expression of TGFBR2 and 3 were similar, TGFBR1 expression was lower (mean FC 0.37) in hIPCs. TGFBR ligands were also differentially expressed. TGFA (FC 0.13) and TGFB2 (FC 0.26) expression was reduced in hIPCs, whereas TGFB1 expression was unchanged. SMAD1–5 expression levels were also similar, while the inhibitory SMAD6 and 7 were reduced (FC 0.23 and 0.48, respectively). RUNX2 and RUNX3, genes associated with osteogenesis and differentiation into mesenchymal-derived tissues, were significantly lower expressed in hIPCs.
Discussion
Mesenchymal stem cells (MSCs) represent multipotent adult precursor populations. Such cells reside in all adult tissues and their multilineage capacity is still not unravelled, since the various niches have not been identified and the signature and potency of MSC of different locations are probably extremely variable [30]. hIPCs have been described to display a mesenchymal signature but they have never been characterized in direct comparison with the “gold standard” of mesenchymal precursors, for example the bone marrow-derived skeletal precursors (hBM-MSC). Here, we show distinct functional signatures of these 2 MSC populations and identify specific key lineage regulatory genes in hBM-MSCs and hIPCs. Our data also indicate a clear transitional state of hIPCs.
Like in recent studies in rodents [11,12] and in humans [10], our experiments strongly suggest that hIPCs derive from mesenchymal cells within the pancreatic islet. This was shown by overlapping immunophenotypes of both cell types, similar expression levels of several mesenchymal markers in hIPCs and hBM-MSCs, and a large vimentin-positive cell population in intact human islets. The latter may give rise to the adherent proliferative islet-derived cell population, hIPCs. In both populations, we detected genes that according to the literature have been related to functional stem cell characteristics like BMI1 [31], HES1 [32], HMGB1 [33], and ABCG2 [34], which may indicate a role of both MSC populations as precursor cells. Data from others suggested the potential of IPCs to adopt mesenchymal lineage phenotypes [10] and the differentiation capacity of BM-MSCs into pancreatic endocrine pathways has also been demonstrated [16]. In contrast, our direct comparison of hIPCs and their differentiation potential with the “gold standard” of MSCs reveals complementary deficits in cell maturation of hIPCs and hBM-MSCs into mesenchymal and endocrine pathways, respectively. SAM analysis of differently expressed gene clusters and functional assignment of gene ontology groups in hIPCs and hBM-MSCs provides evidence for a specified tissue signature and function of these 2 MSC populations.
Genome-wide gene expression profiling shows that the overall pattern of cadherin expression, including the mesenchyme-specific marker cadherin 11, was very similar in both cell populations [8]. However, the HOX expression pattern in hIPCs is remarkably different from hBM-MSCs, strongly indicating that a specific signature (eg, femoral head vs. endocrine pancreas) of MSC populations is maintained when expanded ex vivo, similar to site-specific dermal fibroblasts [35]. HOX genes are regulators of patterning and migration, like anterior–posterior orientation, and positional information for cranial to caudal segments during gastrulation [36] with respect to skeleton and endoderm. HoxA3–6 expression represents the region of foregut and pancreas development. HoxA7–13 genes are associated with hindgut, forelimb, hind limb, and caudal vertebrae development [36,37]. These differences may have consequences as to their respective differentiation capacity since HOX gene interacts with other specific transcription factors of the homeobox type, for example PITX1, which specifies hind limb structures and in analogy PDX and CDX, which specify endocrine pancreas [2,37]. Furthermore, SOX17, a marker for definitive endoderm, has only been detected in hIPCs. The variable expression level of SOX17 in different hIPC preparations (Fig. 6C) suggests different stages of endodermal commitment in these cells. It has been shown that SOX17 is transiently expressed during differentiation of endocrine precursors into islet-like cells, which makes it difficult to detect a consistent expression during the differentiation process [38,39].
This concept of sustained signatures of MSCs can provide important information for site-specific regeneration, based on the positional memory of MSCs at the site of injury, for example vertebrate versus amphibian limb regeneration [40].
The expression profile of mesenchymal and endocrine differentiation key genes in hIPCs and hBM-MSCs clearly reflects the differentiation defects of each population. FGFR2 and RUNX genes, essential for osteogenic differentiation [41,42], are repressed in hIPCs. Lineage specifying members of the MAF gene family [43] are concordant in that mesenchyme-specific MAFK is still high, chondrogenic MAF (c-MAF) is already low, and MAFB, characteristic for the endocrine pancreas, is absent in hIPC.
The transition state suggested by co-labeled insulin–vimentin islet cells is in accordance with the pattern of candidate gene expression that we found in hIPCs, which resembles that of a transition process like EMT or MET [8]. TCF21, which is highly expressed in hIPCs, is a key factor in MET processes, since it is expressed in the mesoderm along the gastrointestinal tube being essential for kidney development and urothelium [29]. EMT and the reverse process MET are important during development, tissue regeneration, cancer biology [8,9], and also for islet cell regeneration [1]. As the transitions are reciprocal processes, genes involved in EMT versus MET are very similar [9]. MET gene expression in hIPCs is mirrored in the well-described urothelium formation in mice (day E10.5–14), where multipotent precursors on their way to epithelial precursors express N-CAM, K-cadherin, and R-cadherin. They lose PAX2 (absent in hIPCs), acquire TCF21, and secrete collagen type 4 as do hIPCs (Table 1). Moreover, TGFb signaling is important in EMT/MET and pancreas development. Inhibition of TGFb signaling in epithelial and mesenchymal compartments in developing mouse pancreas increases the number of endocrine cells, indicating that TGFb restrains and controls the production of β cells from precursors [6]. Some components of TGFb signaling are repressed in hIPCs (eg, TGFBR1, TGFA, and TGFB2). This might indicate a permissive role of hIPCs for islet cell replenishment and raises questions as to the usage of various MSC populations for cell-based manipulation of islets or any other tissue regeneration.
Therefore, our data present strong evidence for the maintenance of a positional mesenchymal signature and for an intermediate phenotype of hIPCs. Their plasticity to either direction is more evident toward the endocrine lineage. These data may even reconcile the conflicting lineage tracing studies that have traced hIPCs to be either of mesenchymal origin [11] or a result of EMT from endocrine cells in culture [14], since the phenotype described here can be acquired through both EMT and MET. Recent results support our findings suggesting that these precursor cells can cycle between epithelial clusters and mesenchymal phenotypes [44].
In conclusion, in this study, we demonstrate that hIPCs and hBM-MSCs display overlapping phenotypes, revealing a common mesenchymal and stem cell character. However, these 2 MSC populations are strongly committed to the tissue where they reside as evidenced by their endocrine and mesenchymal differentiation capacities and their HOXA-related positional memory. These findings may be of great help to dissect the potential of hIPCs to contribute as tools or targets in pancreatic islet regeneration and maintenance.
Footnotes
Acknowledgments
C. Limbert was supported by a Research Fellowship from International Society for Pediatric and Adolescent Diabetes (ISPAD) and the Foundation of Science and Technology (FCT). We are grateful to the Orthopedic Surgeons (J. Eulert, U. Nöth, and L. Seefried, Würzburg, Germany) as well as to the Islet Isolation and Transplantation Centre in Giessen, Germany (M. Brendel, M. Brandhorst, H. Jahr, and R. G. Bretzel) who supplied us with explanted human femoral heads and pancreatic tissue, respectively. We acknowledge the technical assistance (V.T. Monz, S. Zeck, and M. Regensburger) and helpful discussions of our colleagues (A. Heymer and B. Mentrup).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.