
Abstract
There is growing interest in the use of cardiomyocytes purified from embryonic stem (ES) cells for tissue engineering and cardiomyoplasty. However, most transplanted cells are lost shortly after transplantation due to the lack of integration into the host tissue and subsequent apoptosis. Here we examine whether murine embryonic fibroblasts (MEFs) can support the integration of purified murine ES cell-derived cardiomyocytes in a 3-dimensional tissue culture model based on a freezed-dryed collagen matrix with tubular structure. Collagen matrix was seeded either with cardiomyocytes alone or in combination with MEFs. The collagen sponges that were transplanted with cardiomyocytes alone showed neither morphological nor functional integration of viable cells. Cardiomyocytes also did not appear to be capable of attaching quantitatively to any of 16 different 2-dimensional biomaterials. However, cardiomyocytes co-cultured with MEFs formed fiber-like structures of rod-shaped cells with organized sarcomeric structure that contracted spontaneously. Electrical coupling between cardiomyocytes was suggested by strong expression of connexin 43. In addition, MEFs as well as cardiac fibroblasts supported re-aggregation of dissociated cardiomyocytes in hanging drops in the absence of collagen matrix. We conclude that fibroblasts promote cardiomyocyte engraftment and formation of functional 3-dimensional tissue in vitro. Elucidation of the mechanism of this phenomenon may help improve the integration of cardiomyocytes in vivo.
Introduction
U
Kolossov and coworkers have observed that engraftment of ESC-CM in a murine model of heart infarction may be improved by co-transplantation of murine embryonic fibroblasts (MEFs). However, this effect was not quantified and the underlying mechanisms have not been further investigated [7].
Fibroblasts represent, beside cardiomyocytes, one of the most abundant cell types in the developed heart [20,21]. In the early stages of development the heart tube consists of few layers of early cardiomyocytes lined by endocardial cells. The fibroblasts are rare in early stages of cardiac development but make up more than half of the cell number in neonatal hearts [21]. These fibroblasts build up the cardiac matrix by secreting types of collagen that form the skeleton of the heart, mainly collagens type I and III. This collagen skeleton mechanically connects the myocytes and is of importance to guarantee the mechanical properties of the tissue [22].
To assess the role of fibroblasts in engraftment of ESC-CM we have developed an in vitro model based on a synthetic 3-dimensional extracellular matrix composed of collagen type I. In addition, we used the hanging drop method to assess the ability of MEFs to support reaggregation of dissociated ESC-CM into beating cell clusters. We demonstrate that ESC-CM alone were incapable of populating the collagen matrix but when seeded together with MEFs they integrated well and formed 3-dimensional spontaneously beating structures. This effect was independent of the collagen matrix as demonstrated by the ability of MEFs and cardiac fibroblasts to support the aggregation of ESC-CM in hanging drops. These findings suggested that fibroblasts provide mechanical support and/or surface-bound or soluble factors that promote the organization of cardiomyocytes into 3-dimensional structures.
Materials and methods
ES cell culture
For the production of cardiomyocytes the transgenic murine ES cell line αPIG (clone 44) derived from D3 ES cells was used [7]. αPIG44 ES cells express the puromycin resistance gene, puromycin N-acetyltransferase (PAC), and the internal ribosomal entry site (IRES)-flanked enhanced green fluorescent protein (eGFP) gene under the control of a cardio specific α-myosin heavy chain promoter. The ES cells were cultured on mitomycin C-inactivated MEFs in ES cell medium composed of Iscove's modified Dulbecco's medium (IMDM) supplemented with Glutamax®, 17% fetal calf serum (FCS), 100 U/mL penicillin, 100 μg/mL streptomycin, 1% nonessential amino acids, 100 μM β-mercaptoethanol and 1,000 U/mL leukemia inhibitory factor (ESGRO®, Millipore, Schwalbach, Germany). Cells were passaged every 2–3 days. If not otherwise stated, all cell culture media and reagents were supplied by Invitrogen Life Technologies (Karlsruhe, Germany).
Differentiation of ES cells toward cardiomyocytes
To initiate differentiation, ES cells were aggregated in embryoid bodies (EBs) and cultured in differentiation medium (ES cell medium without LIF) as detailed previously [23]. Briefly, ES cells were washed with phosphate-buffered saline (PBS) without Ca2+ and Mg2+ and dissociated by treatment with 0.05% Trypsin/0.2 g/L ethylenediaminetetraacetic acid (EDTA) for 5 min at 37°C. Cells (1 × 106) were resuspended in 12 mL of differentiation medium in a 10 cm bacteriological dish and agitated on a horizontal rocking table at 37°C/5%CO2. On Day 2 EBs were diluted to 1,000 per 10 cm dish in 12 mL medium and further incubated with continuous shaking until Day 8 of differentiation without change of medium. On Day 8, when the first eGFP-positive areas appeared, medium was changed. On Day 9, puromycin was added to a final concentration of 10 μg/mL. On Day 12 medium was replaced and fresh puromycin was added. From Day 14 on, purified cardiac bodies were incubated without puromycin and used for experiments within 1 week.
Dissociation of cardiac bodies into single cells
Cardiac bodies obtained after puromycin selection were washed twice with PBS and incubated with prewarmed collagenase I (1 mg/mL; Roche, Mannheim, Germany) dissolved in collagenase buffer (120 mM NaCl, 5.4 mM KCl, 5 mM MgSO4 × 7H2O, 5 mM sodium pyruvat, 20 mM taurine, 10 mM HEPES, 20 mM glucose, 35 μM CaCl2) for 4 min at 37°C. Dead cells loosely adhering to beating clusters were removed by trituration, 30 mL PBS were added and cardiac bodies were collected at the bottom of the tube by centrifugation at 50g for 3 min. The supernatant containing dead cells and cell fragments was discarded and cleared beating clusters were dissociated into single cells by treatment with collagenase I for 25 min at 37°C. ESC-CM were washed twice with IMDM supplemented with 17% FCS and resuspended in seeding medium consisting of ice-cold IMDM, 17% FCS, 100 U/mL penicillin, 100 μg/mL streptomycin, 1% nonessential amino acids, 100 μM β-mercaptoethanol, and 0.4 mg/mL rat tail collagen type I (Upstate, NY, distributed by Biomol, Hamburg, Germany). Mitomycin C-treated MEFs were trypsinized, washed as described above for ESC-CM, counted and resuspended in seeding medium. All cells were kept on ice until use. An aliquot of dissociated ESC-CM was analyzed by flow cytometry to assess the quality and purity of cells. Routinely, more than 98% of viable cells were GFP-positive cardiomyocytes [23]. In addition, 0.2 × 106 ESC-CM were plated on MEF-monolayer or on 3.5 μg/mL fibronectincoated μ-Dish (Ibidi GmbH, Munich, Germany) to allow microscopic evaluation of their morphology and viability after short- (2–4 days after plating) and long-term (30 days after plating) cultivation.
Preparation of murine embryonic fibroblasts
Murine embryonic fibroblasts were prepared from Day 14.5 embryos of outbread mouse HIMOF1 strain. Briefly, pregnant mice were sacrificed and embryos were isolated under sterile conditions. Head and organs were removed and the remaining tissue was dissociated by digestion with 0.05% trypsin/0.2 g/L EDTA. Cells were cultured on gelatine-coated dishes, expanded over 2 passages and frozen in aliquots. After thawing fibroblasts were passaged for a maximum of 3 times. To stop proliferation cells were treated with mitomycin C (10 μg/mL, Serva) for 3 h at 37°C. Mitomycin-treated MEFs were passaged on fresh culture dishes and were used within 14 days for culturing ES cells, seeding on collagen matrices or for re-aggregation of cardiomyocytes in hanging drops.
Preparation of murine embryonic cardiac fibroblasts
Hearts were first washed in PBS supplemented with 250 μg/mL Fungizone (Gibco, Grand Island, NY) and 10,000 IE/mL Penicillin/Streptomycin (Gibco) and then incubated for 25–35 min at 37°C in 10 mL of 1 mg/mL collagenase D (Roche Diagnostics, Indianapolis, IN) solution, which was prepared in 1× Hank's Balanced Salt Sodium (HBSS) and 10 mM HEPES, pH 7.5 (Gibco). Dissociated cells were filtered through a cell strainer (100 μm pore size) and directly plated in cardiac fibroblast culture medium consisting of DMEM/HamsF12 (1:1), 10% FCS and 10,000 IE/mL Penicillin/Streptomycin. Medium was changed the next day and a few days later confluent cells were further passaged. Cells in a third passage were used for the co-culture with ESC-CMs in hanging drops.
Seeding cells into collagen sponges
Collagen matrices (Matricel GmbH, Herzogenrath, Germany) used in this study was produced from pure bovine collagen type I by a directional solidification method and subsequent freeze-drying [15]. By using this technique parallely oriented tubular structures of collagen can be formed (Supplementary Fig. 1; Supplementary materials are available online at
Attachment of cardiomyocytes on different surfaces
ESC-CM were seeded on different biomaterial surfaces to identify suitable substrates for ESC-CM attachment. We used 4 degradable biopolymers, 8 degradable synthetic polymers, and 4 nondegradable synthetic polymers. The degradable biopolymers were collagen, fibrin, alginate and hyaluronic acid, the degradable synthetic polymers were poly(
ESC-CM were seeded in a density of 20,000 cells/well (96-well format for viability assay, 24-well format for live/dead staining) and cultured for 1 or 7 days in a 20% O2 and 5% CO2 humidified atmosphere. Cell viability was measured using the CellTiter Blue kit (Promega, Mannheim, Germany) according to the manufacturer's instructions. Fluorescence intensity was measured using the fluorometer FLUOstar OPTIMA (BMG Labtech, Jena, Germany). In addition, viability of attached cells was visualized after 1 and 7 days by live/dead staining using fluoresceindiacetate (FDA) and propidium iodide (PI; both Sigma, Steinheim, Germany). Briefly, 1.28 mL Ringer solution containing 20 μL of FDA (0.1 mg in 20 mL acetone) and 20 μL of PI (0.01 mg in 20 mL PBS) was added to the ESC-CM/polymer hybrids. After incubation for 20 s, cells were analyzed using fluorescence microscopy (Polyvar microscope, Leica, Bensheim, Germany).
Aggregation of ESC-CM in hanging drops
In order to determine the influence of MEFs on the capacity of ESC-CM to re-aggregate and form secondary cardiac bodies in hanging drops in the absence of collagen matrix, single cell suspensions of ESC-CM alone or ESC-CM and MEFs were prepared to contain 5 × 104 ESC-CM and 1 × 104 MEFs in 1 mL of seeding medium. Drops (20 μL) containing 1,000 ESC-CM or mixtures of 1,000 ESC-CM and 200 MEFs were pipetted onto the inner surface of lids of U-bottom 96-well plates (Falcon, Becton Dickinson) (Supplementary Fig. 2; Supplementary materials are available online at
Immunohistochemistry
To detect the GFP-positive ESC-CM seeded into collagen matrix, samples were fixed in 4% paraformaldehyde (PFA) for 2 h. To stain the nucleic acids in the cytoplasm and nuclei of the whole cell population, propidium iodide (Sigma-Aldrich, Munich, Germany) was used at a concentration of 5 μg per mL of PBS. After staining, tissue samples were incubated in 18% sucrose in PBS for 1 h at 4°C, embedded in Tissue Tek OCT (Sakura Finetek, Zoeterwoude, NL) and frozen to −80°C. Serial 300 μm cryosections were prepared and analyzed by fluorescence microscopy (Axoivert 200; Zeiss, Jena, Germany).
To perform antibody stainings, ESC-CM grown as a monolayer on fibronectin-coated plates for 2–4 or 30 days after plating were washed with PBS and fixed in 4% PFA for 15 min at RT. Collagen sponges seeded with cells were fixed with 99.8% methanol at −20°C for 15 min. This procedure results in loss of GFP fluorescence but enables recognition of structural cardiomyocyte proteins with the antibodies used. Methanol was removed and samples were incubated in PBS for 15 min. PFA-fixed ESC-CM on fibronectin-coated plates and methanol-fixed cells seeded in collagen sponges were blocked with 5% bovine serum albumin (BSA) in PBS and samples were incubated overnight at 4°C with primary antibodies in 1% BSA prepared in PBS. Primary antibodies anti-cardiac-α-actinin (clone EA-53; 1:800) and anti-connexin 43 (clone CXN-6; 1:400) were purchased from Sigma-Aldrich. Bound antibodies were visualized by anti-mouse IgG1-AlexaFluor 647 or anti-mouse-IgM-AlexaFluor 546 (Invitrogen). Nuclei were stained with Hoechst 33342 (Sigma-Aldrich). Samples were embedded with ProLongGold antifade agent (Invitrogen). Stainings were analyzed on a Zeiss Axiovert 200 fluorescence microscope equipped with Apotome. Image processing was done using Axiovision Release 4.3 (Zeiss) and Corel Draw 11.
Results
Assessment of purity and viability of lineage selected ESC-CM
After initiation of differentiation, GFP-positive and spontaneously contracting areas were first detected in EBs on Day 8 of differentiation. Five days after addition of puromycin, synchronously contracting cardiac bodies (Fig. 1A and 1B) that consisted of more than 98% of GFP-positive ESC-CM were obtained, as shown previously by flow cytometry [23]. The high purity of ESC-CM preparations was also verified by fluorescence microscopy of cells plated on fibronectin-coated plates. As shown on Figure 1C, these cultures contained exclusively GFP-positive cardiomyocytes. Subcellular organization of ESC-CM was further assessed by staining with antibodies specific for cardiac structural protein α-actinin (Fig. 1D). Typical striations reflecting sarcomeric structures were detected in all cells of mostly polygonal shape a few days after plating. However, prolonged cultivation of dense ESC-CM monolayers for 30 days on fibronectin-coated cell culture dishes resulted in maturation of ESC-CM into elongated tube-like myocytes possessing highly ordered striations perpendicular to the longitudinal cell axis (Fig. 1E). ESC-CM could also survive for protracted periods of time (20 days) when plated onto the layer of MEFs (Fig. 1F), but were not capable of adhering in large numbers in uncoated plates without feeders. These analyses demonstrate that pure ESC-CM are capable of surviving in vitro for prolonged periods of time if plated on an appropriate substrate and show signs of morphological maturation in confluent long-term cultures.

Microscopic assessment of cardiomyocytes purity and survival in short and long term cultures. Addition of puromycin to differentiating embryoid bodies at Day 8 of differentiation gives rise to pure cardiac bodies (
Engraftment of ESC-CM into collagen matrix
The engraftment into collagen matrix of ESC-CM seeded alone or in combination with MEFs was monitored daily by fluorescence microscopy. Engraftment was evaluated by visualizing GFP-positive ESC-CM and assessing their morphological appearance. One day after seeding both the ESC-CM seeded alone or with MEFs appeared round-shaped (data not shown). However, by Day 4 after seeding the ESC-CM, which had been combined with MEFs, attached and acquired a typical longitudinal rod-like shape. In contrast, ESC-CM seeded as monocultures still did not attach to the surface of the collagen matrix at that time, remaining round-shaped and retaining their GFP-fluorescence.
One week after seeding the collagen sponges were fixed with PFA and stained with propidium iodide to visualize all seeded cells. Nuclei and cytoplasm of all seeded cells stained red, whereas viable ESC-CM could be detected as GFP-positive cells. Co-cultured MEFs and ESC-CMs appeared to form dense, tissue-like structures when examined under the fluorescence microscope at the periphery of intact sponges (Fig. 2A). To determine whether cells penetrated deep into the sponge, PFA-fixed propidium iodide-stained samples were cut into 300 μm thick slices and analyzed by fluorescence microscopy. These sections clearly showed that seeded ESC-CM were attached to the surface of the sponge and penetrated the outer region of the sponge but did not reach into the core of the material (Fig. 2B). Only MEFs were found at low density in the middle of the matrix 1 week after seeding (data not shown). When analyzed after 1 (Fig. 2C) or 3 weeks (Fig. 2D) of co-culture with MEFs the ESC-CMs stayed attached to the collagen matrix and formed dense elongated structures as demonstrated by strong green fluorescence of GFP-positive cells. The ESC-CM origin of this fluorescence was demonstrated by the presence of cell nuclei in GFP-positive areas (Supplementary Fig. 3A–3C; Supplementary materials are available online at
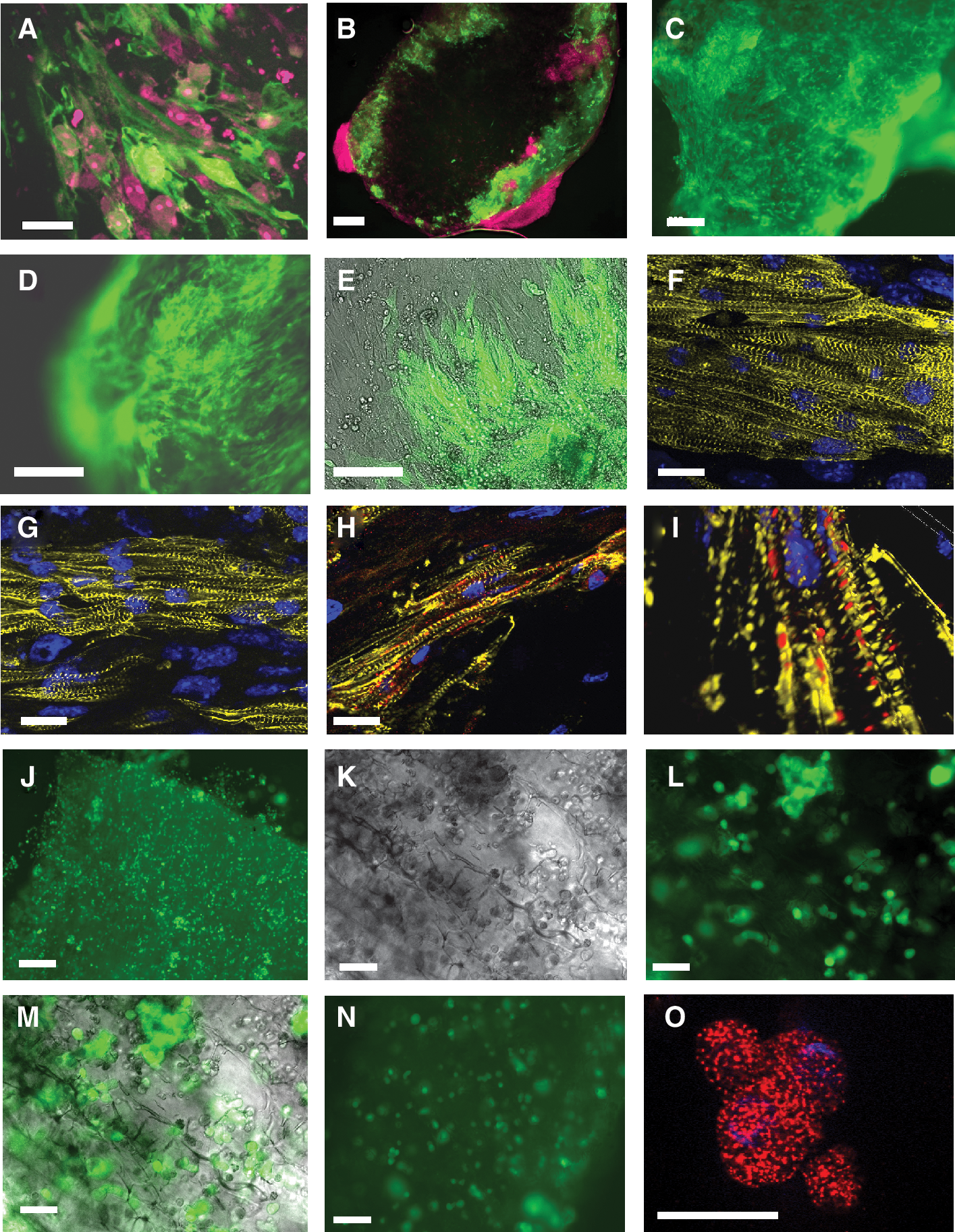
Effect of fibroblasts on cardiomyocyte engraftment into collagen matrices. (
Sarcomeric organization of ESC-CM seeded in collagen sponges
To assess the structure of ESC-CMs seeded into collagen sponges, stainings with antibodies specific for cardiac sarcomeric α-actinin were performed. Analyses were performed with intact sponges in order to preserve the cellular organization in these 3-dimensional structures. One week after seeding distinct pattern of cross striation, which is characteristic of differentiated cardiomyocytes, was found in ESC-CM seeded together with MEF (Fig. 2F and 2G). In some areas, these cells were partially arranged in groups of different sizes and formed fibre-like structures located on the surface of the pores in the collagen matrix. These fibers contained numerous parallelly oriented and tightly packed cells, which did not appear to be highly interspersed by fibroblasts. To further investigate the functional integration of ESC-CM within fibers 1 week after seeding we performed immunohistochemical stainings with connexin 43 antibodies. Gap junctions containing connexin 43 could be identified on the cell–cell interfaces, which indicates that ESC-CMs were electrically coupled within these fibre-like structures (Fig. 2H). The connexin 43-rich junctions appeared to be distributed in regular distances between the cells as demonstrated by the Zeiss Apotome technique that was used to generate a 3-dimensional view of spatially oriented cardiomyocytes (Fig. 2I).
In contrast to these observations, seeding experiments with pure ESC-CMs resulted in an entrapment of ESC-CMs in pores of the collagen sponge after 1 week (Fig. 2J). No elongation and structural maturation of ES-CM was observed. Even after 2 weeks of culture entrapped cardiomyocytes could be identified in the pores of the collagen matrix, which did not appear to show engraftment of elongated cells similar to that observed in co-cultures with MEFs (Fig. 2K–2M). Even after 3 weeks only few GFP-positive ESC-CMs were identified that still retained their round-shaped morphology (Fig. 2N). Stainings of scaffolds seeded with pure ESC-CMs for 1 week with antibodies for cardiac sarcomeric α-actinin revealed a spotted pattern of the actinin protein expression which was clearly different from a striated pattern observed in ESC-CMs co-cultured with MEFs (Fig. 2O). These observations demonstrate that MEFs support the engraftment of purified ESC-CMs in collagen matrices and that ESC-CM alone lack the ability of spreading out on this material.
Attachment of cardiomyocytes on different surfaces
To determine whether the inability of ESC-CMs to attach to a collagen matrix is a specific property of this material or the result of a highly selective requirement of ESC-CMs for a substrate to adhere, we seeded ESC-CMs on a variety of polymer substrates to identify those materials, which would support ESC-CM attachment and viability (Fig. 3 and Supplementary Fig. 4; Supplementary materials are available online at

Attachment of cardiomyocytes to different polymer surfaces. ESC-CM were seeded at a density of 20,000 cells/well in 96-well plates and cultured for 7 days. Cell number/viability was analyzed by measuring the metabolic activity of the cells that adhered to the surfaces. Cardiomyocytes seem to be very selective for attachment to substrates since none of the polymers supported cardiomyocyte attachment comparable to control cells seeded on fibronectin, gelatine, or tissue culture polystyrene (TCPS). One representative experiment from 3 independent experiments is shown. The data are shown as mean values and standard deviations of quadruplicate measurements.
Formation of secondary cardiac bodies in hanging drops
In order to determine whether the ESC-CM alone are capable of forming 3-dimensional aggregates in the absence of collagen matrix, we have cultivated different numbers of dissociated ESC-CM in hanging drops and monitored the formation of secondary cardiac bodies in the presence or absence of MEFs. This method is a widely used approach for inducing differentiation of murine ES cells. In this model, ES cells (usually seeded at 400–600 cells/20 μL drop) aggregate at the tip of hanging drops and form spherical embryoid bodies. Cell Tracker Red CMPTX-labeled MEFs alone formed very compact spherical aggregates after 2 days of culture in hanging drops (Fig. 4). After being transferred onto the surface of a 96-well plate, MEFs attached, grew out of a cluster and spread out onto the surface of a plate few a days later (Fig. 4). However, when purified ESC-CMs were cultured in hanging drops after 2 days they formed loose cell aggregates of irregular shapes containing scattered dead cells around the aggregate, as indicated by the lack of GFP-fluorescence in these cells (Fig. 4). ESC-CMs in these clusters remained GFP-positive (ie, viable) and continued beating for the whole observation period of 16 days but appeared to accumulate cell debris in the course of cultivation (Fig. 4, Supplementary Fig. 5, available online at

Fibroblasts support reaggregation of cardiomyocytes in hanging drops. Hanging drops (20 μL) were formed with single cell suspensions of murine embryonic fibroblasts (MEFs) alone (1,000 cells/drop), ESC-CMs alone (1,000 cells/drop), or admixtures of 200 MEFs and 1,000 ESC-CMs per drop. On Day 2 of culture, cell aggregates formed in hanging drops were transferred to the bottom of a corresponding 96-well plate by centrifugation and the same cell cluster was imaged on days indicated on the left side of each panel row. MEFs were labeled with Cell Tracker Red CMTPX and can be easily identified by their red fluorescence. Viable ESC-CMs express enhanced green fluorescent protein (eGFP) and are depicted as green fluorescent cells. Dead cardiomyocytes lost their eGFP expression and are seen as dark cells scattered around the clusters, mostly in “CM only” aggregates. These images are representative of more than 60 cellular aggregates formed in hanging drops in each experimental group in 3 independent experiments. One additional example of cell aggregates formed in all 3 experimental conditions at different time points is shown in Supplementary Figure 5. Videos corresponding to cell aggregates formed by ESC-CMs alone and ESC-CMs and MEFs in this figure are also available online at
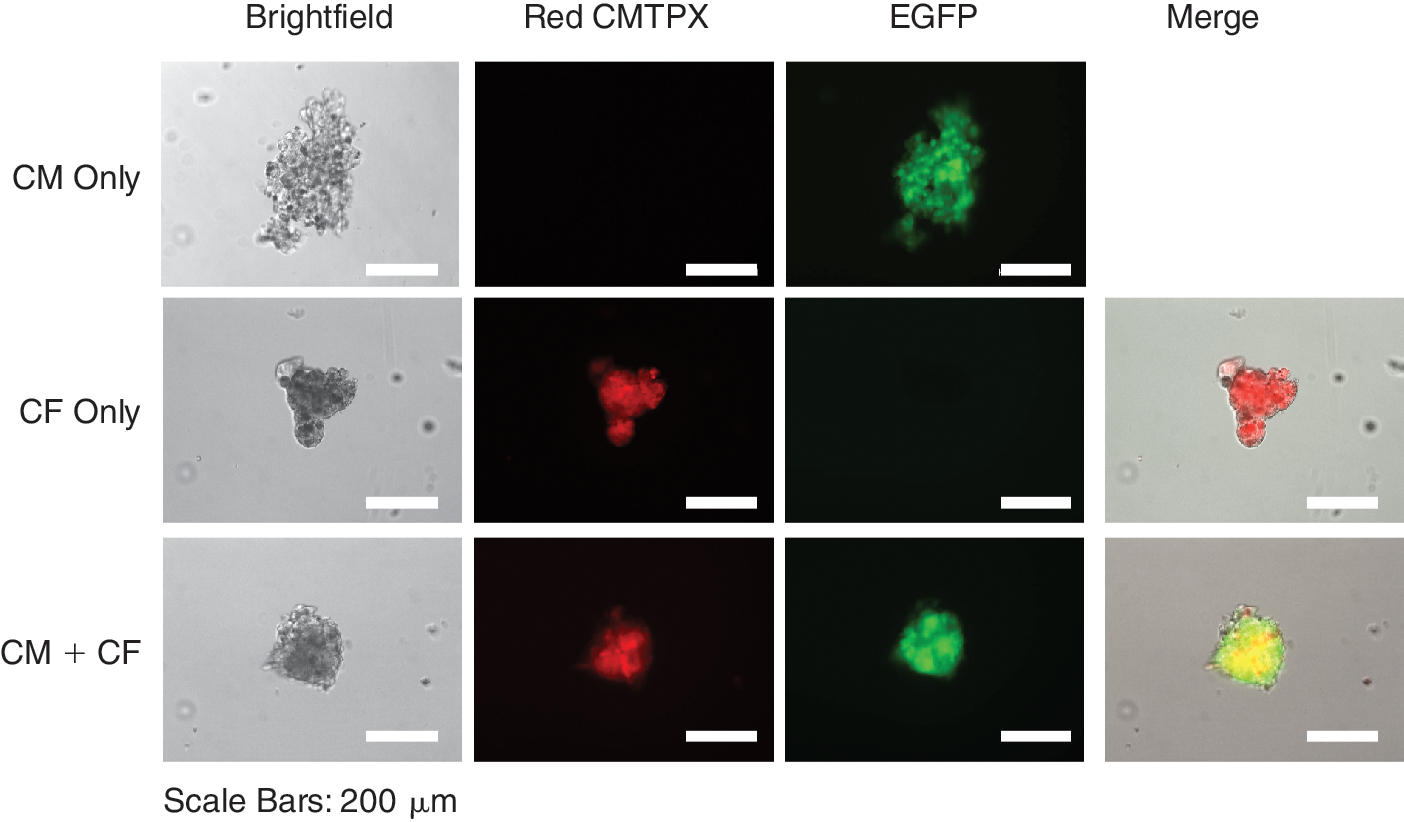
Cardiac fibroblasts support reaggregation of cardiomyocytes in hanging drops. Hanging drops (20 μL) consisting of cardiac fibroblasts alone (1,000 cells/drop), ESC-CMs alone (1,000 cells/drop), or admixtures of 200 cardiac fibroblasts and 1,000 ESC-CMs per drop were prepared and imaged on Day 2 of culture as described in the legend to Figure 4. Scale bars: 200 μm.
Discussion
Developing therapeutic approaches for patients suffering from heart failure by the use of ESC-CMs represents one of the greatest challenges for stem cell research. One of the essential prerequisites for the success of this strategy is a long-term structural and functional integration of highly purified ESC-CMs into the host tissue. Transplantation of purified murine and enriched human ESC-CMs in animal models of heart infarction resulted in functional improvement of damaged heart [7 –9,25,26]. However, cell survival is very poor and >90% of cells are lost in a period as short as few days after transplantation [8]. Kolossov and colleagues have reported that transplantation of lineage selected murine ESC-CMs into syngeneic animals results in complete loss of transplanted cells in 78% of the recipient hearts [7]. Co-transplantation of ESC-CMs with MEFs resulted in a substantial increase of ESC-CM engraftment with a complete loss of cells in only 32% of the animals. Therefore, the presence of supporting cells seems to be of crucial importance for enhancing the retention and survival of transplanted ESC-CMs. Cardiac regeneration can be promoted not only by directly transplanting cells into the myocardium but also by transplantation of in vitro engineered contractile patches composed of different cell types that are present in the heart [27 –30]. In both cases it would be necessary to understand the impact of non-cardiac cells on integration and function of cardiomyocytes.
To examine the interaction between fibroblasts and ESC-CMs, we have developed an in vitro model based on a synthetic 3-dimensional extracellular matrix composed of collagen type I. We employed highly purified ESC-CMs generated by antibiotic selection to avoid interference with other cell types that would otherwise be present in differentiating embryoid bodies. We show here that ESC-CMs alone were incapable of attaching to a variety of 2-dimensional polymers and the 3-dimensional collagen matrix due to poor cell-to-scaffold and cell-to-cell interactions. This conclusion is supported by a spotted appearance of the sarcomeric α-actinin staining in ESC-CMs seeded in collagen matrix, which is suggestive of insufficient cell–scaffold interactions. However, when ESC-CMs were seeded together with MEFs they were retained in the matrix, became elongated and formed 3-dimensional structures of parallelly oriented ESC-CMs that were spontaneously beating. They also exhibited a high degree of sarcomeric organization and were coupled via connexin 43 gap junctions. This finding may explain why ESC-CMs transplanted along with MEFs into cryoinfarcted hearts engrafted better than ESC-CMs transplanted without any supportive cells [7]. The ability of MEFs to support the formation of 3-dimensional tissue of ESC-CMs was not limited only to the collagen matrix but also was seen in hanging drops, in which MEFs as well as cardiac fibroblasts promoted re-aggregation of dissociated ESC-CMs into beating cell clusters in the absence of collagen.
The failure of ESC-CMs to seed the matrix composed of collagen I was unexpected, because collagen I is the major component of the extracellular matrix of the normal heart [9] and in vivo interacts directly with cardiac myocytes via cell surface integrins [20,31 –33]. In addition, neonatal rat ventricular myocytes were shown to adhere in vitro equally well to all types of collagens (I, II, III, and IV) as well as laminin and fibronectin [34]. The reason for the lack of attachment of ESC-CMs to a synthetic collagen I scaffold is unclear but may be due to the absence of appropriate receptors on immature ESC-CMs or lack of recognition of an altered collagen structure that may have been induced in the course of the manufacturing process. However, the inability of ESC-CMs to adhere to 16 different 2-dimensional polymers in this study and to form compact clusters in homotypic CM cultures in hanging drops suggests that ESC-CMs possess highly selective attachment requirements. These requirements appear to be provided by fibronectin-coated tissue culture plates or monolayers of MEFs, where ESC-CMs can adhere and function for prolonged periods of time. The panel of polymers used in this study was earlier utilized by us to find suitable substrates for stem cell-based tissue engineering [24]. Using 7 different types of stem/progenitor cells, we identified polymers, which were universally cytocompatible, such as fibrin or Resomer LT706, and polymers which were selectively cytocompatible, for example, PCL for hematopoietic stem cells, endothelial progenitor cells, and dental pulp stem cells. These data emphasize the selectivity of ESC-CMs for specific 2D and 3D substrates.
Fibroblasts are a highly heterogeneous family of cells displaying developmental and site-specific differences in gene expression profiles, matrix biosynthesis and secretion of bioactive factors such as angiogenic, growth and differentiation factors, and chemokines [35,36]. In the heart, fibroblasts produce extracellular matrix proteins [37], interact with each other as well as cardiac myocytes and play a role in transmission of mechanical force and regulation of the heart wall elasticity. The appropriate matrix plays a crucial role in providing the attachment sites to cardiac cells through integrins. The formation of these contacts leads to adhesion of cells and to clustering of integrins in the cell membrane. Integrins are linked to intracellular proteins of the cytoskeleton and guide organization of protein filaments and may provide survival and maturation signals to cardiomyocytes. The attachment of cardiomyocytes to appropriate matrix compounds may not only support the retention of seeded cells but also influence the organization of cytoskeletal structures, as was demonstrated in this study with ESC-CMs seeded in collagen matrix. Cardiac fibroblasts [38] as well as fibroblasts from other anatomical sites may be a source of angiogenic and/or cardioprotective factors, which may affect the function and survival of surrounding myocytes in a paracrine or direct juxtacrine fashion [39]. For example, fetal lung and skin fibroblasts produce neuregulin 1 [35], which has been reported to act as a pro-survival factor for CMs via PI3K/Act pathway[40], enhance integrin-based myocyte–matrix interactions [41] and promote differentiation of murine ESCs into cardiomyocytes in vitro [42]. Similar paracrine mechanisms may have mediated the positive effect of MEFs and cardiac fibroblasts on attachment and aggregation of ESC-CMs in our in vitro model systems. Recently it was found that embryonic cardiac fibroblasts can activate proliferation of cardiomyocytes revealing a previously unknown paracrine function of cardiac fibroblasts in cardiomyocyte cell cycle regulation [43]. Thus, embryonic fibroblasts seem to have a potential to affect cardiomyocyte behavior by acting directly on them or indirectly on other neighboring cell types. In a more complex in vitro engineered tissue consisting of ESC-CMs, endothelial cells and MEFs, MEFs were shown to promote the survival and proliferation of human endothelial cells and formation of vascular network, which in turn stimulated proliferation, differentiation and maturation of cardiomyocytes [44]. In vitro studies by the work group of Kohl demonstrated that fibroblasts support the formation of organized structure that mimic the in vivo ventricular tissue organization and are capable of forming gap junctions with both each other and with cardiomyocytes [45,46].
However, different types of fibroblasts may differently affect the cardiomyocyte physiology. For example, Ieda and coworkers reported that embryonic cardiac fibroblasts are capable of inducing proliferation of cardiomyocytes while adult cardiac fibroblasts promoted cardiomocyte hypertrophy rather than proliferation [43]. In addition, there are also reports claiming that under the influence of fibroblast-conditioned medium cardiomyocytes exhibit marked hypertrophy and diminished contractile capacity [47]. Therefore, additional studies are required to clarify the issue of how various types of fibroblasts and other cell types affect cardiomyocyte behavior. Future investigations should define the optimal combinations of cells for in vitro cardiac tissue engineering and cell transplantation in order to cure diseased heart. Our data indicate that it is crucial to provide the cardiomyocytes with an environment that supports their integration and structural maturation. This may include the addition of supportive cells or other types of substrates and substances that promote cell adhesion and prevent cell death upon transplantation. In vitro models, such as those employed in this study, may allow for controlled investigation of interaction between different cell types and provide valuable information that could be tested in a more complex in vivo system. These in vitro assays may also facilitate the rational design of safer and more efficient cardiac cell therapies.
Footnotes
Acknowledgments
Collagen matrices were a generous gift from Dr. Ingo Heschel, Matricel GmbH, Herzogenrath, Germany. We thank Katja Martina Eckl and Damir Jacob Illich for isolation of cardiac fibroblasts. We thank Nadin Lange, Rebecca Dieterich, and Stephanie Brosig for excellent technical support and Susannah Stone for proof-reading. This study was supported by grants from the Federal Ministry for Education and Research to T.Š. and H.J. (Grant Nr. 01 GN 0947), and the Else-Kröner Fresenius Stiftung (Grant Nr. 85/2009) and the Köln Fortune Program to T. Š. (Grant Nr. 85/2009). LPF was supported by a stipend from the Köln Fortune Programm (KF133/2007).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.