
Abstract
Mesenchymal stem cell (MSC)-mediated tissue regeneration offers opportunities to regenerate a bio-root and its associated periodontal tissues to restore tooth loss. Previously, we proved that the apical end of developing root was acting as a promising candidate cell source for root/periodontal tissue (R/PT) regeneration. In the present study, we investigated the properties of periapical follicle stem cells (PAFSCs) isolated from the apical end of developing root of human third molars at the root-developing stage and evaluated the potential application of these cells for cementum/periodontal ligament (PDL) regeneration and bio-root engineering. Putative PAFSCs were isolated and subcultured until 20th passage. Cell characteristics of PAFSCs at early or late passage were evaluated and compared with periodontal ligament stem cells (PDLSCs) via a series of histological, cellular, and molecular analyses. PAFSCs at early passage presented crucial stem cell properties and showed a higher proliferation rate than PDLSCs in vitro. Meanwhile, PAFSCs also showed the tissue-regenerative capacity to produce a typical cementum/PDL-like complex in vivo. During long-term passage, both cell populations changed in morphology and gradually lost their stem cell properties. The alkaline phosphatase (ALP) activity and expression of mineralization-related genes markedly declined as more passages were carried out, which might lead to the loss of tissue-regenerative capacity of these 2 groups of cells in vivo. Our findings suggest that developing tissue-derived PAFSCs are a distinctive cell population from PDLSCs and might be a promising candidate for bio-root engineering.
Introduction
T
Compared with tooth crown formation, the development of R/PT involves a complicated interacting program and participation of multiple cell types [12,13]. The apical end of developing root has been proved to contain periapical follicle, dental papilla, as well as Hertwig's epithelial root sheath (HERS), which is the “growth center” of tooth root and rich in stem cells for R/PT regeneration [12,14,15]. In our previous study, we proved that the apical end of developing root in rat featured unique “embryonic” characteristics that not only provided a proper niche to induce differentiation of PDLSCs along the cementoblastic lineage [8], but also contained stem/progenitor cells capable of differentiating into multiple tooth root- and periodontium-forming cells [16]. To date, stem cells have been found in both the apical papilla (stem cells from apical papilla, SCAP) and periapical follicle (periapical follicle stem cells, PAFSCs) of the apical end of developing root, which feature unique “embryonic” characteristics [10,17]. As the progenitor cells of dental pulp, although similar in some characteristics to dental pulp stem cells (DPSCs), SCAP may represent a population of early stem/progenitor cells distinctive from that of mature tissues, behave differently in a number of aspects, and possess greater capacity for dentin/pulp complex regeneration than DPSCs [10,18]. It is generally believed that periapical follicle differentiates into the PDL, alveolar bone, and the mineralized bone-like cementum around the tooth root surfaces [19 –21]. As the progenitor cells of periodontium, PAFSCs have been also isolated from this developing organ primordium of root and proved their stem cell characteristics in vitro [17]. Although research efforts in PDLSCs biology isolated from adult tissues offer approaches to regenerate periodontal tissues, preliminary studies indicate that PAFSCs derived from developing tissues, which feature unique “embryonic” characteristics, might possess a greater regeneration capacity for R/PT engineering as an alternative promising candidate [7,17]. However, so far there is no relevant experimental study to prove the differentiation capacity of PAFSCs in vivo. It still remains elusive whether PAFSCs represent distinct stem cell population from PDLSCs and are potential candidate cells to regenerate periodontal complex. Therefore, the aim of this study was to investigate characteristics of PAFSCs isolated from human third molars at the root-developing stage and tissue-regenerative capacity in vivo compared with PDLSCs to evaluate the potential usage of PAFSCs for R/PT engineering.
Materials and Methods
Cell culture
PAFSCs were isolated and cultured as previously described [17]. According to the patients' panoramic radiographs, normal human impacted third molars (n = 10) at root-developing stage were collected from 9 individuals (12–15 years of age) after obtaining written informed consent. The experimental protocol was approved by the Committee of Ethics of Fourth Military Medical University (FMMU) and conducted in accordance with the Guidelines. Extracted teeth were placed into chilled sterile vials containing 8 mL phosphate-buffered saline (PBS) separately. Then they were transported on ice to the laboratory and processed immediately. First, they received several washes in sterile PBS. Then periapical follicles were gently removed with a scalpel from where they attached to the root dentin (Fig. 1A). The loose connective tissues were minced into 1 mm3 cubes and placed into 6-well culture dishes (Costar, Cambridge, MA). The explants were grown in α-minimum essential medium (α-MEM; Gibco BRL, Gaithersburg, MD) supplemented with 10% fetal bovine serum (FBS), 0.292 mg/mL glutamine (Invitrogen, Carlsbad, CA), 100 U/mL penicillin (Gibco BRL), 100 μg/mL streptomycin (Gibco BRL), and 100 μM
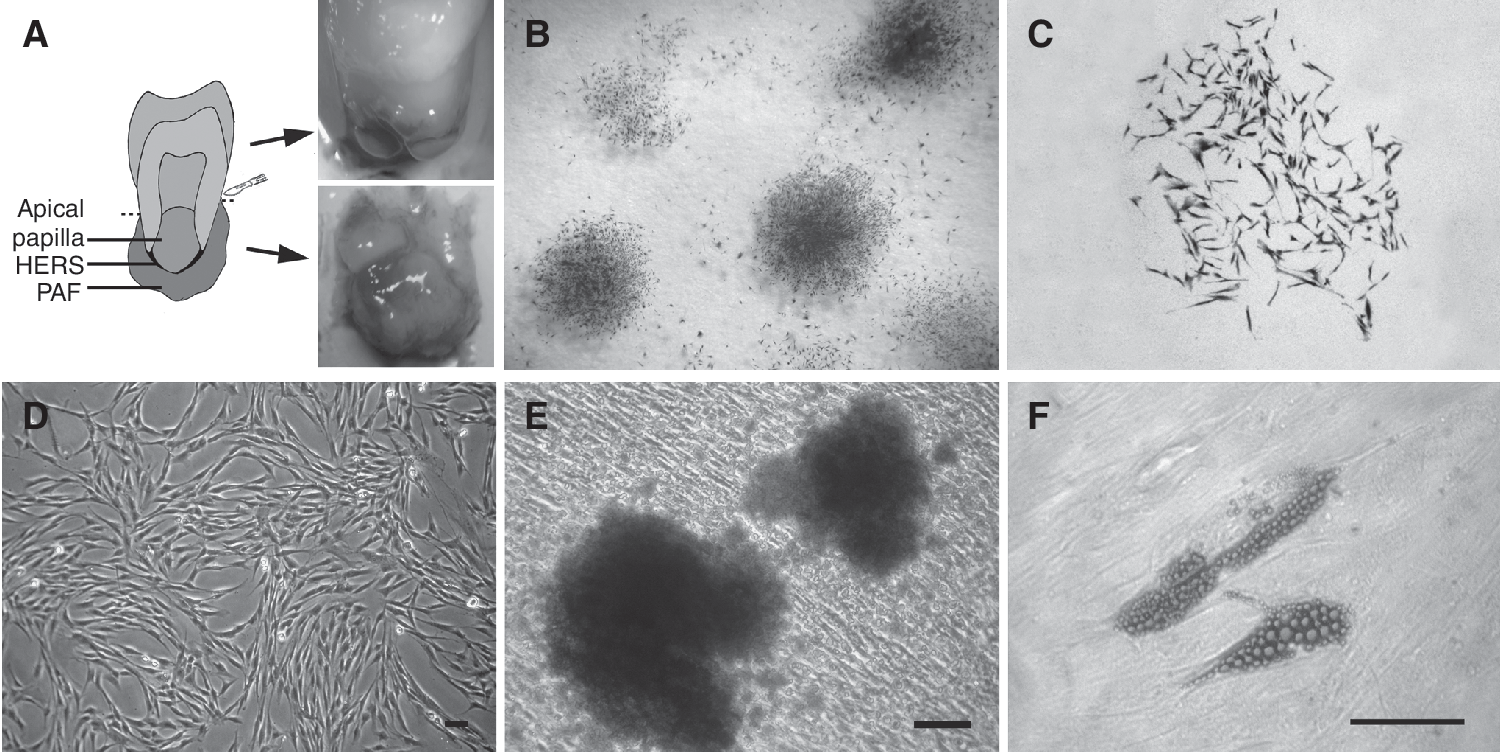
Isolation and characterization of periapical follicle stem cells (PAFSCs;
Human PDLSCs were isolated and cultured as previously described [7,8]. Normal premolar teeth intended for extraction due to orthodontic reasons (n = 14) were collected from 7 individuals (at 13–15 years of age). PDL tissues were gently separated from the middle third of root surface of multiple numbers of premolars, cut into 1-mm3 cubes, and cultured as same as what PAFSCs have been done. Multiple colony-derived PAFSCs and PDLSCs from 3rd to 20th passages were used during the experiments. In the comparison experiments, PAFSCs and PDLSCs were obtained from the same donors. For each experiment, the same passage of PAFSCs and PDLSCs was used. Morphological appearance and cell motility were observed and photographed under phase-contrast inverted microscopy (CK40-F200; Olympus Optical Co. Ltd, Tokyo, Japan).
Migration assay
The 5 × 104 PAFSCs and PDLSCs at 3rd passage were seeded in 6-well dishes, respectively. The cultures were maintained at 37°C in a humidified atmosphere of 5% CO2, and 95% air until exponential phase of growth. Using a pipette tip, a wound was produced in the monolayer at 2 different positions on each plate. The adherent monolayer was then washed 2 times in PBS to remove nonadherent cells. After 0, 24 h, the each open wound surface area was quantified by phase-contrast inverted microscopy (CK40-F200; Olympus Optical Co. Ltd, Tokyo, Japan) at 4 different positions. Each experiment was repeated at least 3 times.
Cell counting
The 1 × 104 PAFSCs and PDLSCs at 3rd passage or 20th passage were plated in 6-well dishes, respectively. Culture medium was changed every 2 days. After 2, 4, 6, 8, and 10 days of culture, plates were washed twice with PBS to remove unattached cells, and attached cells were collected by trypsinization. The number of cells was counted using a hemocytometer.
Bromodeoxyuridine (BrdU) incorporation assay
Single-cell suspensions of PAFSCs and PDLSCs at 3rd passage or 20th passage were, respectively, cultured in 24-well plates at a density of 1 × 104 cells per well. After serum starvation-induced cell cycle synchrony, 10 μM BrdU (Sigma-Aldrich, St. Louis, MO) was added 2 h prior to fixation with 4% polyoxymethylene. BrdU incorporation in proliferating cells was revealed using immunostaining against anti-bromodeoxyuridine antibody (BrdU; 1:100 dilution; Boster Biotechnology, Wuhan, China). Cells containing densely brown-stained nuclei with clear morphology were considered BrdU-positive, and were counted in 5 fields per well (center and at 3, 6, 9, and 12 o'clock). The number of BrdU-positive cells in the same frame was counted. Results were expressed as the percentage of BrdU-positive cells against total cells counted.
Flow cytometry for stem cell surface markers
The stem cell surface antigen expression for STRO-1 and CD146 (R&D Systems, Inc., Minneapolis, MN) was assessed by fluorescence-activated cell sorting (FACS) analysis according to the manufacturer's protocol. The 1 × 106 PAFSCs and PDLSCs at 3rd passage or 20th passage were washed in PBS, and then incubated with mouse anti-human STRO-1 and CD146 antibodies for 30 min at 4°C. Cells were washed twice with cold PBS containing 2% FBS and incubated with 1 μg of fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse IgG or IgM antibodies (Santa Cruz, Santa Cruz, CA) for 30 min at 4°C. Mouse isotype antibodies (BD Bioscience, San Jose, CA) served as control. Labeled cells were analyzed by using a flow cytometer (Beckman Coulter, Fullerton, CA).
Colony-forming efficiency
To assess colony-forming efficiency, 1 × 103 PAFSCs and PDLSCs at 3rd passage or 20th passage were cultured within 100-mm dish and maintained for 14 days, respectively. Then they were fixed with 70% ethanol and stained with 0.1% crystal violet by the method described by Gronthos et al. [22]. Aggregates of over 50 cells were counted as a colony under microscopic observation. Colony-forming efficiency was presented as colony number per 1 × 103 cells plated. Each experiment was repeated at least 3 times.
Osteogenic and adipogenic differentiation protocols
PAFSCs and PDLSCs at 3rd passage or 20th passage were, respectively, plated into 6-well culture dishes at a concentration of 1 × 105/well. Cultures were allowed to reach 70% confluence before differentiation was initiated. Then normal α-MEM medium was removed, and replaced with the osteogenic medium (α-MEM supplemented with 10% FBS, 50 μg/mL of ascorbic acid, 10 mM of sodium β-glycerophosphate, and 100 nM of dexamethasone) or the adipogenic medium (α-MEM supplemented with 10% FBS, 0.5 mM of methylisobutylxanthine, 0.5 μM of hydrocortisone, 60 μM indomethacin, and 10 μg/mL insulin). Cells were maintained with the fresh differentiation medium every 4 days for 4 weeks.
Alizarin red staining for calcium deposits
After 4-week induction, the samples were fixed with 4% polyoxymethylene for 30 min; Alizarin red staining was performed to determine mineralization as previously reported [22]. The mineralized nodules were imaged and analyzed with Leica Q-Win (Leica, Cambridge, UK) image analysis software.
Oil red O staining for cytoplasmic fat globules
After 4 weeks of treatment, the cells growing under adipogenic conditions were washed twice with PBS and fixed in 70% ethanol for 15 min. Oil red O staining was performed as previously reported [23]. The amount of Oil red O accumulated within each well was quantified using a standard curve produced with a serial dilution of the Oil red O stain in isopropanol.
Alkaline phosphatase (ALP) activity assay
For quantitative analysis of alkaline phosphatase (ALP) activity, single-cell suspensions of PAFSCs and PDLSCs at 3rd passage or 20th passage were seeded at a density of 1 × 103 cells/well into 96-well plates and cultured in α-MEM supplemented with 10% FBS, 0.292 mg/mL glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, and 100 μM
Quantitative real-time PCR
PAFSCs and PDLSCs at 3rd passage or 20th passage cultured in α-MEM supplemented with 10% FBS, 0.292 mg/mL glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, and 100 μM
In vivo differentiation assay
To further investigate the in vivo differentiation capacity of PAFSCs, an in vivo transplantation assay was performed. PAFSCs and PDLSCs at 3rd passage or 20th passage were implanted into twelve 6-week-old severe combined immunodeficiency (SCID) mice. Each immunodeficient mouse received a set of 4 same grafts and the total number of the grafts composed of the same cell type at each passage was 12. For a single transplant, ∼2.0 × 106 cells were mixed with 40 mg of ceramic bovine bone (CBB; Research and Development Center for Tissue Engineering, Fourth Military Medical University, Xi'an, China) powders, and then implanted into subcutaneous pockets on the dorsal surface of immunodeficient mice as described previously [25]. All procedures were reviewed and approved by the Animal Care Committee of Fourth Military Medical University. The implants were recovered at 6 weeks post-transplantation, fixed with 4% paraformaldehyde for 2 days, then decalcified for a further 14 days in 10% EDTA (pH 8.0) prior to embedding in paraffin. For histological analysis, 5 μm sections of implants were yielded and stained with either hematoxylin and eosin (H&E) or Masson trichrome staining.
Statistical analysis
All statistic analyses were performed using SPSS® version 15.0 software (SPSS Inc., Chicago, IL). All values were expressed as mean ± standard deviation. For analysis of cell counting, ALP activity, BrdU incorporation, STRO-1/CD146 expression, colony-forming efficiency, Alizarin red staining, Oil red O staining, and real-time PCR assay, the Fisher-protected least significant difference (PLSD) test was used to compare the different study groups. Cell migratory capacity between groups was analyzed for significance using Student's t-test. A P value <0.05 was considered statistically significant.
Results
Isolation and identification of PAFSCs
Periapical follicles collected from just one tooth were capable of providing larger number of tissues than PDLs did. Of the 24 teeth processed, dental tissues were placed into eight 6-well culture dishes (48 wells, 34 wells for PAF tissues, and 14 wells for PDL tissues). And 15/48 (31.3%, 11 wells for PAF tissues, and 4 wells for PDL tissues) wells became contaminated with yeast or yeast/bacteria despite the presence of penicillin and streptomycin in the culture medium. Vigorous cell growth was observed in the rest (33/48 wells, 68.7%) cultures after 14 days. The frequency of successful cell cultures in present study was similar to the study of Perry et al. with DPSCs [26].
PAFSCs formed adherent clonogenic cell clusters (CFU-F, colony-forming unit, fibroblastic) at a low density (Fig. 1B and 1C), similar to other mesenchymal stem cell (MSC) populations. The majority of PAFSCs retained their fibroblastic spindle shape (Fig. 1D). To investigate the differentiation potential of PAFSCs, multiple colony-derived PAFSCs at 3rd passage were supplemented with osteogenic medium to induce mineralization in vitro. After 4 weeks of induction, cultured PAFSCs formed extensive amounts of Alizarin red-positive mineral deposits throughout the adherent layers (Fig. 1E). Moreover, PAFSCs were capable of undergoing an adipogenic differentiation with an adipogenic-inductive cocktail (Fig. 1F).
Morphological observation
Changes of cell morphology were evaluated using an inverted light microscope (Olympus Optical Co. Ltd., Japan). The PAFSCs at 3rd passage were of a typical fibroblastic spindle shape. However, following several passages, the cell populations gradually acquired a longer spindle-shaped fibroblastic morphology. PDLSCs exhibited similar morphological changes to PAFSCs throughout the culture period.
Cell motility and proliferation activity
To assess cell motility, a scratch assay was performed onto the PAFSCs and PDLSCs at 3rd passage. Interestingly, 24 h after this injury, a clear motility difference was observed (Fig. 2A–2C). The PAFSCs population showed an improved migratory capacity (PAFSCs vs. PDLSCs: 0.87 ± 0.42 vs. 0.58 ± 0.37 mm; P = 0.035; n = 3 cultures). We subsequently assessed the proliferation activity of 2 cell populations. BrdU incorporation experiments were performed. As shown in Figure 2D and 2E, PAFSCs at 3rd passage showed a higher proliferating capacity than PDLSCs (PAFSCs vs. PDLSCs, cell counting, days 4, 6, 8, 10: P = 0.042, 0.027, 0.021, 0.011; BrdU incorporation, P = 0.035; n = 3 cultures). However, the proliferation rate of PAFSCs and PDLSCs significantly slowed down at 20th passage (3rd passage vs. 20th passage: cell counting—PAFSCs, days 4, 6, 8, 10: P = 0.005, 0.002, 0.002, 0.003; PDLSCs, days 4, 6, 8, 10: P = 0.037, 0.033, 0.012, 0.009; BrdU incorporation—PAFSCs, P = 0.001; PDLSCs, P = 0.034; n = 3 cultures), and most of them have quit the cell cycle.
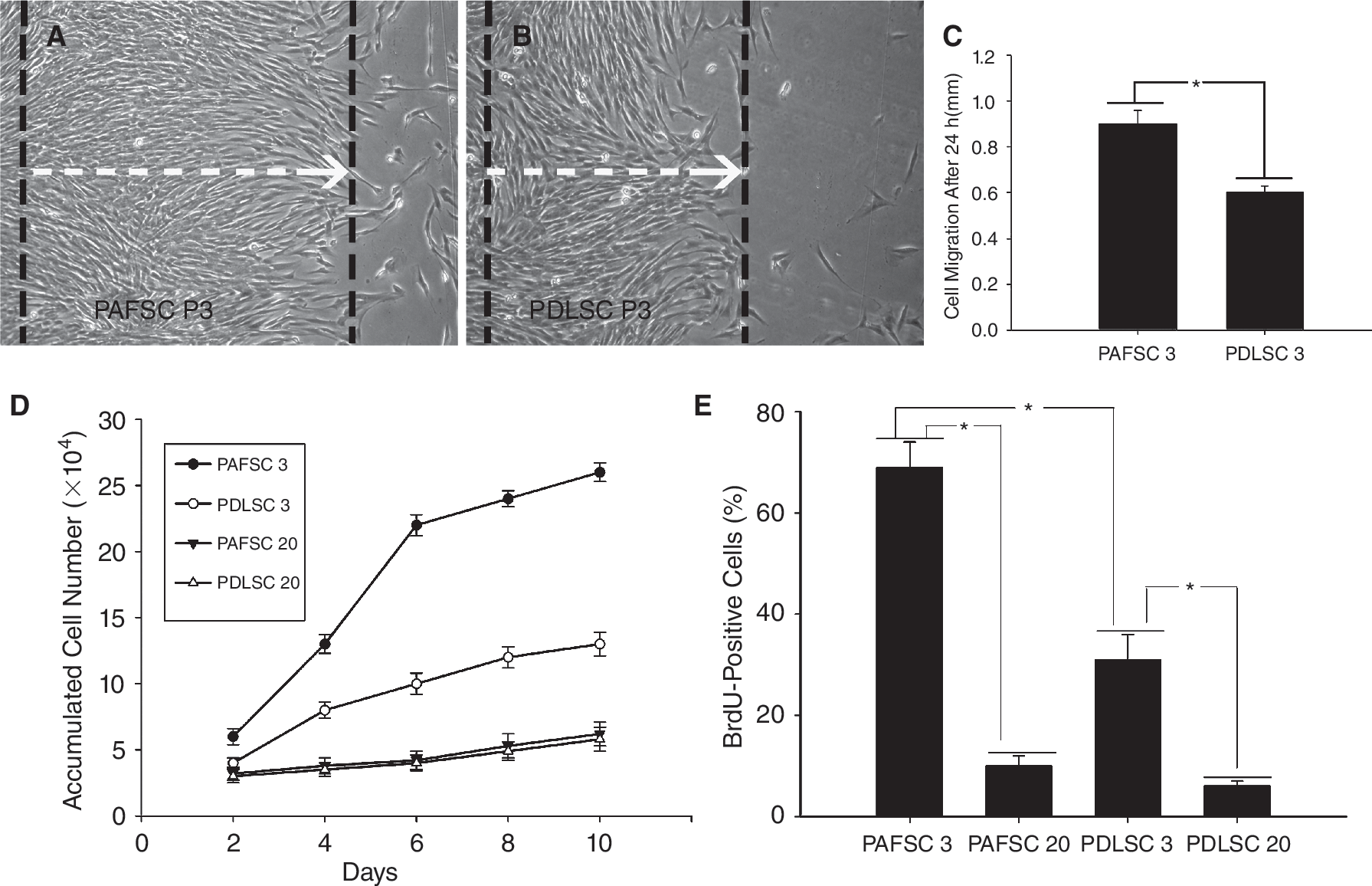
Cell motility and proliferation activity of periapical follicle stem cells (PAFSCs) in comparison with periodontal ligament stem cells (PDLSCs). (
Stem cell properties analysis
Flow cytometric analysis showed that PAFSCs expressed stem cell surface markers—STRO-1 and CD146. The percentage of STRO-1+ PAFSCs was 20.21% ± 1.3% at 3rd passage and 2.84% ± 0.69% at 20th passage; while the rate of CD146+ PAFSCs was 90.05% ± 1.8% at 3rd passage and 18.36% ± 2.4% at 20th passage. A similar change was also found in PDLSCs (STRO-1+: 3rd passage, 13.37% ± 3.21%, 20th passage, 1.89% ± 0.94%; CD146+: 3rd passage, 80.67% ± 6.14%, 20th passage, 8.54% ± 4.07%). The positive ratio of STRO-1 and CD146 from PAFSCs (at 3rd passage) was much higher than that of PDLSCs (PAFSCs vs. PDLSCs: STRO-1+, P = 0.021; CD146+, P = 0.044; n = 3 cultures). After long-term passage, the positive percentage dropped down significantly in both cell populations (3rd passage vs. 20th passage: STRO-1+—PAFSCs, P = 0.002; PDLSCs, P = 0.016; CD146+—PAFSCs, P = 0.028; PDLSCs, P = 0.033; n = 3 cultures).
Both cell populations showed the ability to form adherent clonogenic cell clusters (Fig. 3A). At 3rd passage, PAFSCs formed nearly twice the number of colonies compared with PDLSCs (PAFSCs vs. PDLSCs: 169.15 ± 15.14 vs. 86.14 ± 10.73; P = 0.025; n = 3 cultures); at 20th passage, the number of colonies of both cells was greatly decreased (PAFSCs, 15.25 ± 3.52; PDLSCs, 13.58 ± 2.06. Third passage vs. 20th passage: PAFSCs, P = 0.002; PDLSCs, P = 0.016; n = 3 cultures).
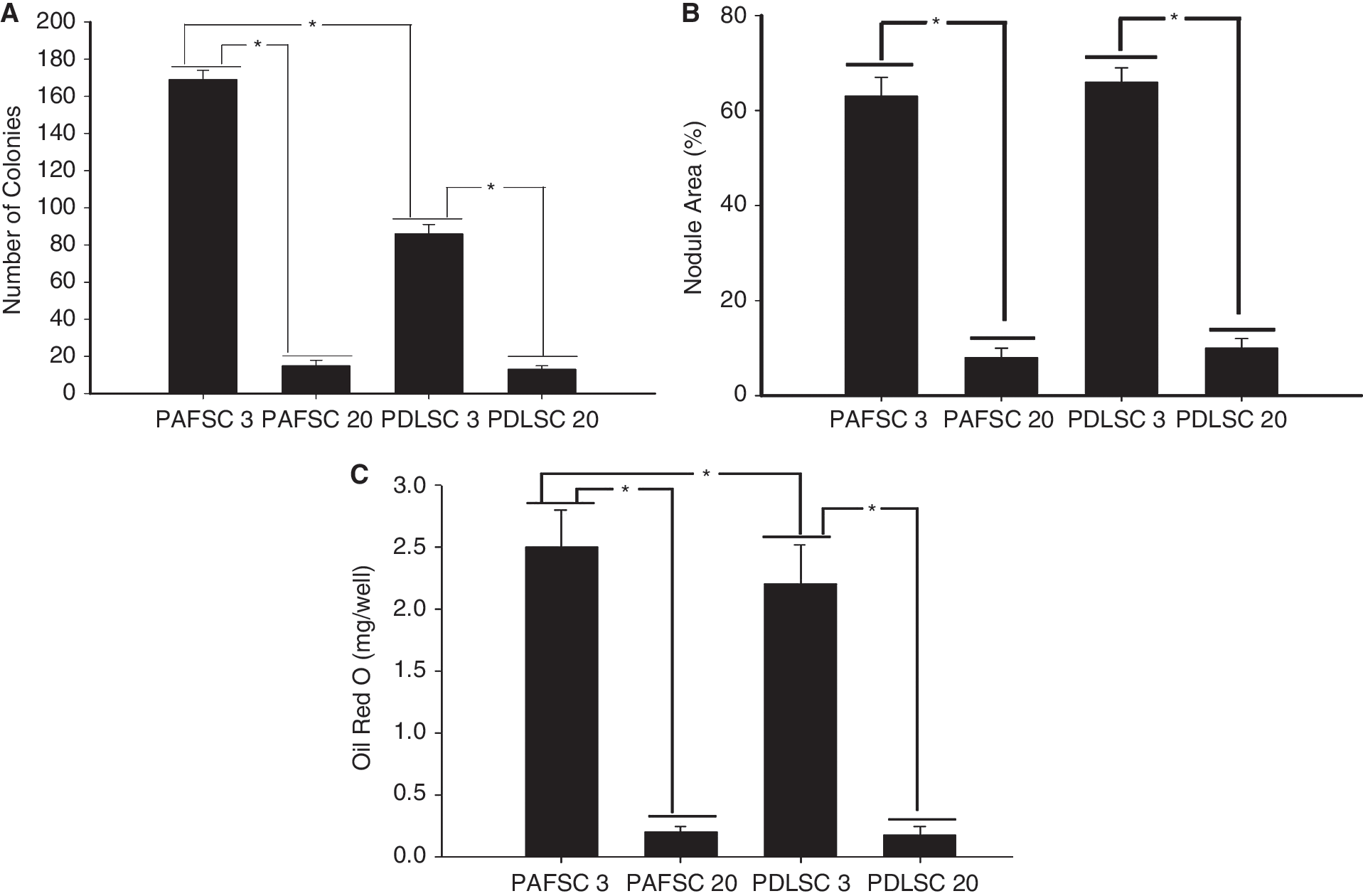
Stem cell properties of periapical follicle stem cells (PAFSCs) in comparison with periodontal ligament stem cells (PDLSCs). (
Furthermore, we assessed the effects of prolonged passage on the differentiation potential of cultured PAFSCs and PDLSCs along the osteogenic and adipogenic lineages (Fig. 3B and 3C). At 3rd passage, both groups showed calcium deposits as visualized by Alizarin red staining and quantification of calcium areas showed no significant difference (PAFSCs, 63.34% ± 4.23%; PDLSCs, 66.92% ± 3.71%; P < 0.05; n = 3 cultures); however, accumulated Oil red O levels of PAFSCs were much higher than that of PDLSCs (PAFSCs vs. PDLSCs: 2.54 ± 0.37 vs. 2.21 ± 0.32 mg/well; P = 0.039; n = 3 cultures). With extended passage, both PAFSCs and PDLSCs in the present study gradually lost their multilineage differentiation potential (20th passage nodule areas: PAFSCs, 8.31% ± 2.52%, PDLSCs, 10.06% ± 2.08%; Oil red O levels: PAFSCs, 0.28 ± 0.05 mg/well, PDLSCs, 0.18 ± 0.07 mg/well. The 3rd passage vs. 20th passage: nodule areas—PAFSCs, P = 0.011; PDLSCs, P = 0.007; Oil red O levels—PAFSCs, P = 0.009; PDLSCs, P = 0.023; n = 3 cultures).
ALP activity and the expression of mineralization-related genes
Since ascorbic acid can stimulate cell proliferation and collagen synthesis of fibroblasts in vitro [27,28], we have used α-MEM culture medium supplemented with 100 μM
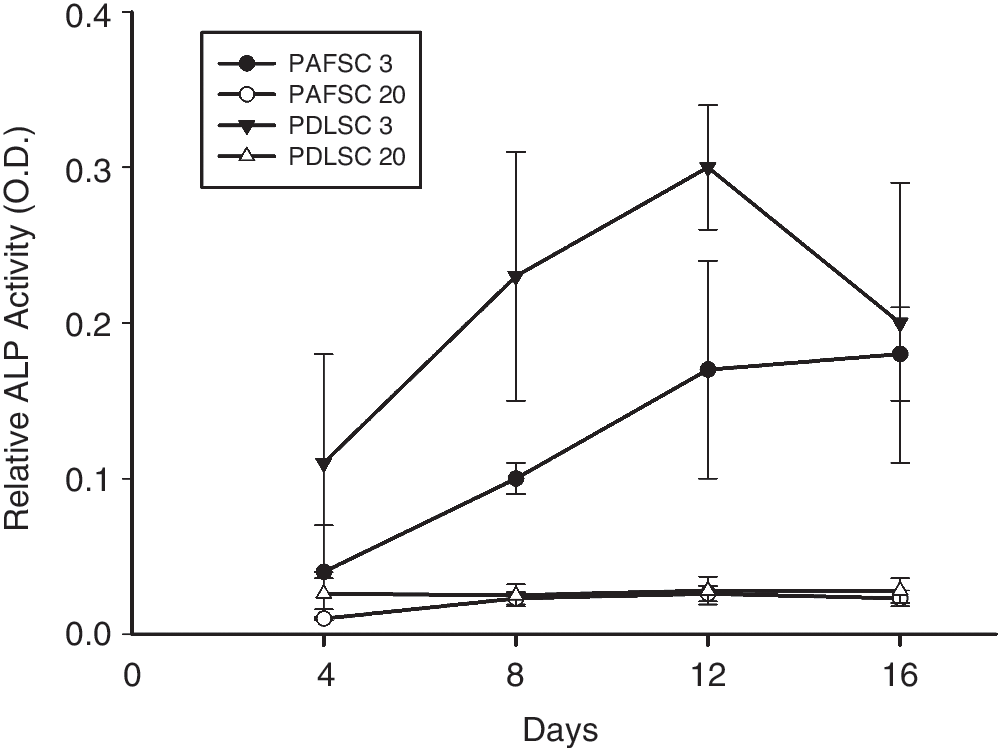
Alkaline phosphatase (ALP) activity of periapical follicle stem cells (PAFSCs) in comparison with periodontal ligament stem cells (PDLSCs). Note that ALP activity of PAFSCs at 3rd passage was lower than that of PDLSCs and decreased significantly after long-term culture.
To investigate the expression of genes responsible for PAFSCs and PDLSCs differentiation, quantitative real-time PCR analysis was performed (Fig. 5). At 3rd passage, PAFSCs showed lower levels of COL-1, Runx2, and OCN mRNA than PDLSCs (PAFSCs vs. PDLSCs, COL-1, Runx2, OCN: P = 0.028, 0.036, 0.007; n = 3 cultures). In all the samples tested, the expression of COL-1, Runx2, and OCN mRNA decreased significantly with increased passage number (3rd passage vs. 20th passage—PAFSCs, COL-1, Runx2, OCN: P = 0.009, 0.003, 0.028; PDLSCs, COL-1, Runx2, OCN: P = 0.007, 0.009, 0.014; n = 3 cultures).

Gene expression of periapical follicle stem cells (PAFSCs) in comparison with periodontal ligament stem cells (PDLSCs). (
In vivo differentiation potential
A total of 48 specimens in current study were harvested and evaluated by light microscopy (DX51, Olympus, Tokyo, Japan) after HE and Masson trichrome staining. Among 12 implants of PAFSCs at 3rd passage, 2 (16.7%) formed cementum-like mineralized tissues lining the CBB surfaces; the mineralized structure resembled cellular cementum with cementocyte-like cells embedded in the matrix (Fig. 6A and 6B). At higher magnification, Masson trichrome staining of implants of PAFSCs at 3rd passage revealed that clusters of small short PDL-like fibrous tissues interfaced with the newly formed cementum-like deposits, mimicking the physiological attachment of natural Sharpey's fibers in normal PDL (Fig. 6C). The rest transplantations of PAFSCs at 3rd passage did not exhibit any sign of mineralized tissue formation. It has been demonstrated that PDLSCs were able to form cementum/PDL-like tissues upon in vivo transplantation [7,30,31]. Likewise, implants of PDLSCs at 3rd passage in present study showed ability to form typical cementum/PDL-like structures. The 4 of 12 (33.3%) specimens produced a thin layer of cementum-like tissues around the surface of CBB, and long large collagen bundles were inserted perpendicularly into these cementum-like tissues (Fig. 6D–6F). No mineralized deposits were observed in transplants of PAFSCs and PDLSCs at 20th passage (100%), and only connective tissues were formed (Fig. 6G and 6H). And no mineralized or PDL-like tissues were observed within the CBB alone (Fig. 6I).
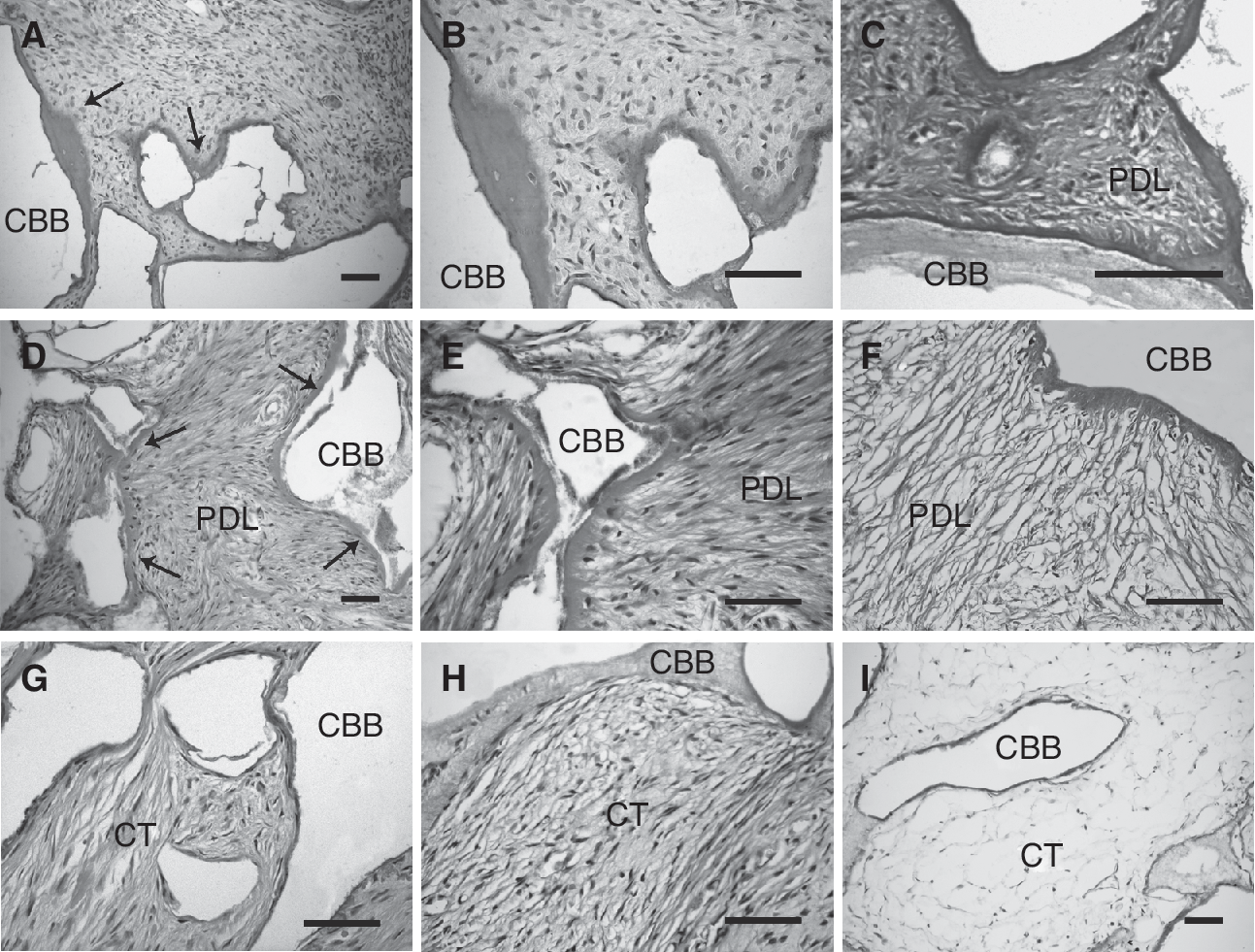
In vivo differentiation potential of periapical follicle stem cells (PAFSCs) in comparison with periodontal ligament stem cells (PDLSCs). (
Discussion
Dental follicle (DF) is a loose connective tissue sac surrounding the unerupted tooth and participating in tooth eruption and periodontium formation [32,33]. Thus, mesenchymal cells from periapical follicle of the root part in dental follicle have been classically considered as precursor cells for cementoblasts, fibroblasts, and osteoblasts, which build up the periodontal structures including cementum, PDL, and alveolar bone [19 –21]. In this study, we found that PAFSCs derived from dental follicle at the root-forming stage represent a novel population of multipotential MSCs as demonstrated by their capacity to develop into mineralized tissue-forming cells and adipocytes in vitro. This suggests that it might be a good stem cell pool for R/PT regeneration. They also express high levels of STRO-1 and CD146, putative surface markers for MSCs, and have relatively high proliferating activity, cell migration rate, and colony-forming ability. Meanwhile, they also have similar multilineage differentiation capability as PDLSCs. These data therefore confirmed that PAFSCs are a novel and unique population of postnatal stem cells from developing tissue.
The identification of human PDLSCs within the PDL undoubtedly represents a significant progress toward predicable periodontal regeneration. Sonoyama et al. [10] have successfully used both human SCAP and PDLSCs to produce dentin and PDL on a HA/TCP carrier and further to generate a root/periodontal complex able to support a porcelain crown. However, our results, as well as other reports, indicate that developing tissue-derived PAFSCs, with excellent proliferation rate, might be a more competent candidate source for R/PT regeneration [17]. It is well known that specific phenotypic characteristics of seeding cells determine the structure and function of regenerated tissues [34]. Therefore, the origin of stem cells plays an important role in tissue engineering [35]. During the past few years, various bioengineering studies have used different approaches in selecting epithelial and mesenchymal tissues to build up a “complete” tooth, and it is very likely that these tissues or cells may be too early to respond appropriately for initiation of root formation [2,36 –40]. From a development perspective, only cells at the bell and cap stages, and similar mesenchymal and epithelial primordial tissue elements, can theoretically signal and respond for R/PT formation, just as stem cells from root apical papilla (SCAP) differentiate into odontoblasts to regenerate dentin. As for postnatal PDLSCs isolated from mature periodontal tissues, since it is relatively easy to attain and are capable for cementogenic differentiation, they may be well suitable for cell-based therapies for periodontal repair. However, compared with mature tissue-derived PDLSCs, PAFSCs showed faster growth rate, more BrdU uptake, and higher migration capability at early passage, which are in good agreement with previous data [17]. Therefore, similar to SCAP, PAFSCs originated from just one tooth can provide large amount of stem cells sufficient for PAFSCs-mediated R/PT regeneration as they manifest excellent proliferation potential.
The present study also evaluates the in vivo tissue-regenerative capacity of PAFSCs. Although it is generally believed that research efforts in adult stem cell (eg, PDLSCs, DPSCs) biology offer additional approaches to the engineering of individual dental tissues (dentin/pulp-like complex and cementum/PDL-like structure), preliminary studies indicate that postnatal stem cells (eg, SCAP, PAFSCs) derived from developing tissues, which feature an unique “embryonic” characteristics and possess a greater proliferation capacity, are probably promising candidate cell source for R/PT regeneration [7,10,17,18,22]. Since the developing tissue can serve as a better stem cell source for tissue regeneration than mature tissue [10,41,42], we expect that PAFSCs from the apical end of developing root would have a better periodontal reconstruction capability to produce a typical tissue structure of periodontium including cementum, PDL, and even alveolar bone. Previous studies indicated that dental follicle cells from the root surface of bovine tooth germ were able to form cementum-like matrix in vivo [25,43], and precursor cells from dental follicle of human third molar teeth also presented osteogenic differentiation in vitro and in vivo [19]. As the progenitor cells of periodontium, it has been proved that PAFSCs at early passage presented crucial stem cell properties and showed a higher proliferation rate than PDLSCs in vitro in the present study. Meanwhile, PAFSCs also showed the tissue-regenerative capacity to produce a cementum/PDL-like complex in vivo. In current study, part of PAFSCs (at 3rd passage) transplants produced a typical cementum/PDL-like structure that were characterized by a thin layer of cementum-like mineralized tissues and associated PDL-like collagen fibers connecting with the newly formed cementum. Although the absence of a suitable marker for cementum hampers the precise identification of the mineralized tissues as cementum, several histological features attributed to cementum, such as mineralized matrix with irregular inserted collagen bundles like Sharpey's fibers, lack of hematopoietic tissues, and relatively low cellularity, strongly imply that this tissue regenerated by PAFSCs is of a periodontal nature [7,10,30,31,44]. It is interesting that low percentage of PAFSCs samples formed cementum/PDL-like structures in vivo than PDLSCs at early passage, which is a somewhat unexpected result compared with SACP and DPSCs [10]. If our observation in the present study was further proved by more extended research, then the following explanations may be possible. First, the PAFSCs and PDLSCs of different developing stages may lead to the discrepancy in differentiation capability in vivo; second, it is also evidenced by the difference of ALP activity and expression level of mineralization-related gene profiles (Runx2, OCN, and COL-1) between PAFSCs and PDLSCs at early passage. Altogether, current results suggest that creating the biological microenvironment similar to those of a developing milieu in the apical end of developing root, which could elicit the intrinsic capacity of PAFSCs to proceed to the development of a cementum/PDL-like complex, is of critical importance for future applications.
It is worth to mention in current study that PAFSCs and PDLSCs undergo the characteristic changes during passages. Observation of 2 cell populations in long-term culture reveals a remarkable change both in vitro and in vivo. These changes include: (1) morphological change; (2) gradual losing of stem cell properties; (3) decline in proliferation potential, ALP activity, and the expression of mineralization-related genes; and (4) loss of in vivo tissue-regenerative capacity. Although it has been recognized that stem cell-based tissue engineering (ie, DPSCs, PDLSCs, and BMMSCs) represents a novel and advantageous treatment for R/RT regeneration, up to now a well-defined protocol for the rapid and efficient expansion of these cells to create a homologous tissue-engineered implant for clinical purposes is still unavailable [45]. Bonab et al. [46] reported that the MSCs start to mature almost undetectably from the moment of in vitro culture and lose their multipotential nature after long period culture. Itaya et al. [11] also showed that cultured PDL-derived cells display obvious de-differentiation during passage for both osteogenesis- and tendo/ligamentogenesis-related genes. Consistent with these observations, as more passages go on, both PAFSCs and PDLSCs (at 20th passage) in the present study lost differentiation capability and produced connective tissues without any hard tissue formation in vivo, which is partly confirmed by the decline of ALP activity and expression of mineralization-related genes. In this regard, the relatively great number and outstanding proliferation potential of developing tissue-derived PAFSCs, especially in earlier passages, are doubtlessly of great significance for stem cell-mediated R/PT regeneration. Furthermore, the cementogenic microenvironment is critical and needs to be optimized to validate the roles of PAFSCs in R/PT regeneration.
From tissue engineering point of view, understanding the mechanism of tooth development in adult stem cell opens up a new way to improve the efficacy of functional tooth engineering [40]. It is well known that tooth development actually depends on the reciprocal and sequential inductions of epithelial and mesenchymal cells [12,13]. R/PT development also follows this principle. In a series of classical tissue recombination experiments, MacNeil and Thomas [47,48] showed cells from dental follicle tissue could produce an adherent cementum-like mineralized tissue on the surface of dentin fragments, where root-associated basement membrane (RBM) was preserved. Furthermore, when epithelial root sheath (HERS) was included in recombinations between root dentin specimens and dental sac, cementum-like tissue also formed in the absence of RBM; meanwhile, part of recombination in which RBM was preserved formed PDL-like tissues. This suggests HERS and basement membrane play important roles in cementogenesis and PDL formation. Therefore, in order to mimic a bio-physiological root/periodontal setup in vivo, we conjecture that the combination of PAFSCs, SCAP, and HERS might be a valuable strategy in future bio-root engineering. In this method, cell populations (including SCAP, HERS, and PAFSCs) isolated from the apical end of developing root will be loaded in regular sequence on an absorptive root-shaped scaffold. By this way, we can mimick a bio-physiological root/periodontal setup in vivo, and regenerate a functional R/PT complex able to support porcelain crown in situ (Fig. 7).

Schematic diagram illustrating the potential strategy for bio-root engineering. First, autologous stem cells from apical papilla (SCAP), Hertwig's epithelial root sheath (HERS), and periapical follicle stem cells (PAFSCs) from the apical end of developing root are isolated and expanded in vitro. Second, cell populations are loaded in a regular sequence on an absorptive root-shaped scaffold, mimicking a bio-physiological root/periodontal setup in vivo, to make a bio-root. Then, the bio-root is transplanted into the patient's jaws for continuous growth/development. Finally, post crown restoration is performed to recover the original tooth function. Abbreviations: PDL, periodontal ligament; C, cementum; D, dentin; OD, odontoblast.
In conclusion, in the present study we characterized a novel population of multipotent stem cells PAFSCs, isolated from human tooth germs at the root-forming stage, and found that these cells were highly proliferative and had the potential to generate a cementum-like matrix in vivo. However, these characteristics were lost in long-term culture, together with a change in histological, cellular, and molecular properties. These results suggest that PAFSCs might be promising candidate cells for cementum/PDL regeneration and bio-root engineering.
Footnotes
Acknowledgments
The authors thank Dr. Liang Tang, Department of Oral Histology and Pathology, School of Stomatology, Fourth Military Medical University, for his kind help. This work was supported by grants from the Nature Science Foundation of China (Project No. 30572046 and No. 30725042). The authors report no conflicts of interest related to this study.
Author Disclosure Statement
No competing financial interests exist.