
Abstract
Although the bone marrow (BM) microenvironment is the main inducer niche of early B lymphopoiesis during the adult life, other extramedullar microenvironments, such as the liver, may also have potential for supporting B-cell development. Previously, we reported that murine liver sinusoidal endothelial cells (LSECs) support in vitro and in vivo hematopoietic stem cell (HSC) proliferation and myeloid differentiation. In the present study, we investigated the capacity of LSEC to promote B lymphopoiesis from BM progenitor lineage-negative (Lin−) cells. Murine BM Lin− cells were co-cultured with LSEC, in the absence of exogenous cytokines. B cells were characterized by flow cytometry and cytokine expression by RT-PCR. We show that BM Lin− cells differentiated to early B-lymphoid progenitors (B220+) and subsequently to mature (CD19+) B cells. Functional studies showed the presence of a high number of non-adherent cells (NACs), collected from lipopolysaccharide (LPS)-treated Lin−/LSEC co-cultures, expressing IgM on their surface (sIgM). Colony formation from NAC was observed in the presence of IL-7 (CFU-IL-7). LSEC constitutively express IL-7, Flt-3L, and SCF at the mRNA level, and VCAM-1 on their surface, which may explain the capacity of these cells to promote B lymphopoiesis. These data demonstrate that LSEC promote all stages of B lymphopoiesis. To our knowledge, this is the first report that LSEC constitute an in vitro microenvironment for B lymphopoiesis. Further studies will establish whether LSEC can serve in vivo as a B-lymphopoietic niche under physiological or pathological condition, or when HSC are mobilized.
Introduction
B-
Although the BM constitutes the main hematopoietic niche in the adult, under certain clinical conditions other extramedullar microenvironments, such as at the liver, may also support the hematopoietic process [14 –16]. We have shown, in an in vivo experimental model of extramedullary hematopoiesis, the existence of a specific vascular niche for HSC constituted by the liver sinusoidal endothelial cells (LSECs) [16]. Furthermore, we have shown that the LSEC support the survival, proliferation, and differentiation of HSC, in vitro [16]. The ability of LSEC to support in vitro and in vivo hematopoiesis may be explained by their capacity to secrete numerous hematopoietic cytokines and express cellular adhesion molecules associated with the retention of HSC at the liver niche [16,17].
In the BM, recent reports have shown that osteoblasts and endothelial cells (ECs) play an important role in inducing B lymphopoiesis [18 –21]. While previous evidence had shown that the hematopoietic differentiation from HSC promoted by LSEC is mainly to myeloid progenitors [16], it is not known whether these cells may also support the differentiation of HSCs to B cells. In this work, we investigated the capacity of the LSEC to promote the production of B-cell precursors from BM progenitor lineage-negative (Lin−) cells. Our results show that LSEC support all stages of B lymphopoiesis from HSC to early B-cell progenitor and generation of mature IgM+ B cells. These data indicate that the LSEC constitute an in vitro microenvironment for B lymphopoiesis, and suggest that these cells might serve in vivo as a liver B-lymphopoietic niche.
Materials and Methods
Animals
Female C57BL/6 mice, 6–8 weeks of age, were used for this study. All animal experimentation was performed in accordance with institutional guidelines.
Reagents, monoclonal antibodies
FITC or phycoerythrin (PE)-conjugated rat monoclonal antibodies (Moabs) anti-mouse: B220, CD19, CD106 (VCAM-1), and goat polyclonal antibodies against mouse IgM were purchased from Pharmingen (San Diego, CA). Recombinant murine IL-7 was purchased from R&D Systems (Minneapolis, MN) as carrier-free preparations. Lipopolysaccharide (LPS) from Escherichia coli, 055:B5 serotype was purchased from Sigma (St. Louis, MO).
LSEC isolation and culture
The isolation, cloning, and characterization of LSEC have been previously reported [16,17]. LSEC express the EC markers flk-1, Von Willebrand factor (vWF), CD34, and VCAM-1 [16,17]. LSEC were maintained in Dulbecco’s modified Eagle’s medium (DMEM; Gibco BRL, Grand Island, NY)–10% fetal bovine serum (FBS; Sigma) and used for experiments after the cells reached 70% confluence.
Isolation of BM lineage-negative (Lin−) cells
Animals were sacrificed by cervical dislocation, and the femurs were removed aseptically. BM cells were obtained by flushing the femurs with DMEM medium. To obtain a population of primitive hematopoietic progenitors, BM Lin− cells were isolated by immunomagnetic separation following the manufacturer’s recommendations (Dynal Biotech ASA, Oslo, Norway). In brief, BM cells were incubated with a cocktail of lineage antibodies (including those against Mac-1, Gr-1, B220, Ter119, and CD3), and submitted to immunomagnetic cell separation. The cellular negative fraction (BM Lin− cells) was obtained from the supernatant and used for further assays.
Isolation of spleen cells
Animals were sacrificed by cervical dislocation, and the spleens were removed aseptically. Spleens were minced and then homogenized by disruption between sterile glass slides in DMEM. The cell suspension was passed through nylon mesh and washed once. Splenocytes were resuspended in DMEM supplemented with 10% FBS, and plated at a density of 1 × 106 cells/cm2 in culture flasks. After 24 h, non-adherent cells (NACs) were removed, and seeded on LSEC monolayers.
LSEC and BM Lin− co-cultures
BM Lin− cells were seeded (1 × 105 cells per well) onto a confluent monolayer of LSEC cultures in 24-well plates with DMEM medium containing 10% FCS. Half of the co-culture medium was collected and replaced every 5–7 days for fresh medium. The number and the viability of cells present in the non-adherent fraction (NAC) of the Lin−/LSEC co-cultures were determined by trypan blue staining. Aliquots of NAC from each well were assayed for expression of B-cell markers by flow cytometry (see below), and for morphology in cytospin preparations by Wright-Giemsa staining. BM Lin− cells were also co-cultured with LSEC under non-contact conditions. For this purpose, a 0.4 µm transwell insert (Corning, NY) was placed into the well containing a LSEC monolayer, and the BM Lin− cells were added to the insert. The number and the viability of cells present in the insert were examined by trypan blue after 8 days of culture.
Detection of B cells by flow cytometry analysis
NAC, harvested every 5–7 days from Lin−/LSEC co-cultures, were assayed for the expression of B-cell markers (B220 and CD19) by flow cytometry. In some experiments, Lin−/LSEC co-cultures were treated with LPS (10 µg/mL) for 4 days, and NAC harvested from them were examined for the expression of surface IgM (sIgM). Data collection and analysis of the fluorescent intensities were carried out using a FACSort (Becton Dickinson, San Jose, CA). Ten thousand events were acquired and analyzed using the CELLQuest software program.
Conditioned media experiments
The possible presence of soluble factors in the supernatant of LSEC able to promote the survival and proliferation of B cells was investigated. Briefly, LSEC were grown to 90% confluence, and culture medium was replaced by fresh medium, and incubated for 48 h. After incubation, the conditioned media was removed, centrifuged at 2,000g for 15 min, and passed through a 0.2-mm filter before use. NAC collected from Lin−/LSEC co-cultures were cultured in the presence of LSEC-conditioned media and assayed by flow cytometry for the expression of B-cell markers. As control, NAC harvested from BM Lin−/LSEC co-cultures were cultured in the presence of regular medium.
Assay of colony-forming cells
NAC harvested, every 5–7 days, from Lin−/LSEC co-cultures were assayed in a fibrin clot assay [16] to determine the potential of colony formation of these cells in the presence or absence of interleukin-7 (CFU-IL-7). As control, an enriched population of B-lymphoid progenitors (B220+ cells) was obtained from BM by immunomagnetic separation, and used for the CFU-IL-7 assay. After 7 days of culture, the presence of colonies was evaluated under an inverted microscope (Zeiss-Axiovert, Germany). The presence of CFU-IL-7 reflects the presence of early and functional B cells.
Detection of IL-7, Flt-3l, and SCF transcripts in LSEC by RT-PCR
Total RNA was isolated from monolayers of LSEC clones using TRI REAGENT (Molecular Research Center, Cincinnati, OH). One microgram of the total RNA was reverse-transcribed in a total volume of 20 µL in buffer containing 250 mmol/L Tris–HCl (pH 8.3), 250 mmol/L KCl, 50 mmol/L MgCl2, 50 mmol/L dithiothreitol, 2.5 mmol spermidine, 10 mmol/L deoxynucleotide mixture, 100 pmol/L random hexamer oligonucleotides, and 15 U AMV reverse transcriptase (Promega, Madison, WI). PCR amplification of the cDNA was then performed using specific oligonucleotides (Table 1) for the detection of IL-7, Flt-3L, SCF, and β-actin transcripts. PCR conditions were 1 min of denaturation at 95°C, annealing at 55°C for 1 min, and extension at 72°C for 1 min for 35 cycles. Analysis of the PCR products was performed by comparing them with the predicted PCR fragment size after ethidium bromide staining of the PCR products separated by electrophoresis in a 1.5% agarose gel.
Statistical analysis
Results are reported as mean ± standard deviation (SD) from triplicate dishes in all assays. We tested the data from the experiments for statistical significance using the Student’s t-test. A value of P < 0.05 was considered significant. All experiments were repeated at least 3 times and have documented reproducibility.
Results
LSEC promote B-cell differentiation from BM Lin− cells
Previously, we showed that LSEC promote long-term hematopoiesis in vitro [16]. In this work, we were interested in evaluating whether LSEC could support the survival, proliferation, and differentiation of B-cell precursors from lineage-depleted BM Lin− cells. Flow cytometry studies demonstrated that LSEC clone used in this work expressed the EC markers flk-1 (VEGF-R2), and CD34 on their surface, but did not express the myeloid markers CD11b and Gr-1 (Fig. 1). Other characteristics of the LSEC have been previously reported [16,17]. We seeded BM Lin− cells on a confluent LSEC monolayer in the absence of exogenous hematopoietic cytokines. After 14 and 21 days of incubation, there was a significant increase in the number of NAC collected from BM Lin−/LSEC co-cultures (Fig. 2A). At the third week of co-culture, the number of NAC had increased almost 3-fold compared with the previous week (P < 0.05; Fig. 2A). The production of cells in BM Lin−/LSEC co-cultures was maintained for >7 weeks, and the viability of harvested NAC was always >90% (data not shown). The hematopoietic proliferation in the BM Lin−/LSEC co-culture was dependent upon a direct contact with LSEC, because a significant reduction of NAC-BM cells was observed after 8 days in the non-contact transwell cultures (data not shown). Microscopic observation of BM Lin−/LSEC co-cultures showed the presence of a high number of rounded cells (Fig. 2B) and hematopoietic foci (Fig. 2C) on the LSEC monolayer.
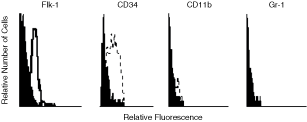
Expression of endothelial cell (EC) markers on liver sinusoidal endothelial cell (LSEC). Flow cytometry analysis of LSEC showing the expression of flk-1 and CD34, and the absence of CD11b and Gr-1 expression. Negative controls were stained with the respective isotype (black histograms).
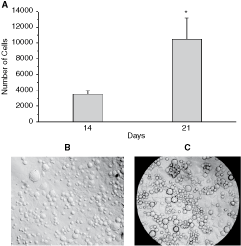
Liver sinusoidal endothelial cells (LSECs) promote the proliferation and survival of bone marrow (BM) lineage-negative (Lin−) progenitor cells. Non-adherent cells (NACs) were collected from BM Lin−/LSEC co-cultures and the cell number and viability were determined by trypan blue (
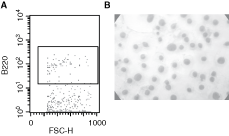
Next, we evaluated the capacity of the LSEC to promote the differentiation of BM Lin− cells to B cells. For this purpose, the NAC harvested from BM Lin−/LSEC co-cultures were examined for the expression of the specific early B-cell marker, b220, using flow cytometry. B220+ cells were found after 27 and 41 days of co-culture (Fig. 3A). These cells were detected as early as after 7 days of co-culture (data not shown). Cytospin and wright-giemsa staining of NAC collected from BM Lin−/LSEC co-cultures showed the presence of lymphoid-like cells (Fig. 3B).
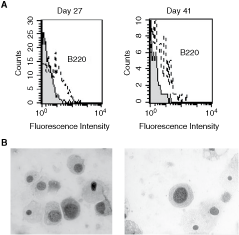
Liver sinusoidal endothelial cells (LSECs) promote B lymphopoiesis from bone marrow (BM) Lin. BM lineage-negative (Lin−) were co-cultured on LSEC monolayer. Non-adherent cells (NACs) were collected, at days 27 and 41, from BM Lin−/LSEC co-cultures, and the cells were analyzed for their expression of the B220 marker by flow cytometry (
LSEC support the generation of functional B cells from BM Lin−
We evaluated whether B cells generated from BM Lin−/LSEC co-cultures maintained their functional capacity to differentiate to early and mature stages of B-cell differentiation. For this purpose, we examined the expression of CD19 on NAC collected from BM Lin−/LSEC co-cultures. A small percentage of cells expressing CD19 were detected in the NAC fraction, as early as 7 days of BM Lin−/LSEC culture (data not shown), but a higher percentage of CD19+ cells were found in the NAC fraction collected at 21 and 41 days (Fig. 4A and 4B, respectively). Next, we examined the functional capacity of B cells generated from BM Lin−/LSEC co-cultures to express sIgM. While low levels of sIgM+ cells (2%) were found in the NAC fraction of BM Lin−/LSEC co-cultures (Fig. 4C), a higher percentage (32%) of sIgM-expressing cells were detected in the NAC collected from BM Lin−/LSEC co-cultures treated with LPS for 4 days (Fig. 4D). Bigger and round cells were observed in BM Lin−/LSEC co-cultures treated with LPS (Fig. 4F), as compared with untreated co-cultures (Fig. 4E). Together, these results indicate that B cells generated from BM Lin−/LSEC co-cultures treated with LPS had the functional capacity to express IgM. Soluble IgM was not detected by ELISA in supernatants from BM Lin−/LSEC co-cultures treated with LPS (data not shown). Further functional analysis was performed by examining the capacity of colony formation of NAC in the presence of IL-7 (CFU-IL-7). After 7 days of culture, CFU-IL-7 were observed (Fig. 4G), indicating the presence of B-cell progenitors in the NAC population generated from BM Lin− co-cultured on LSEC monolayers. Bigger colonies and a higher number of CFU-IL-7 were obtained from B-lymphoid progenitors (B220+ cells) isolated from BM than those observed from NAC collected from BM Lin− co-cultures (Fig. 4H). No CFU-IL-7 were detected in the absence of IL-7 (data not shown).
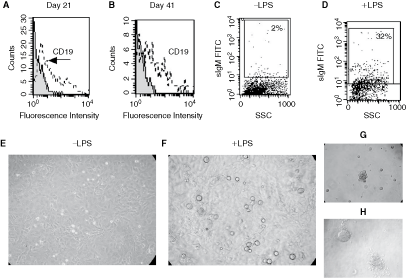
B-cell marker expression and functional analysis of non-adherent cell (NAC) collected from bone marrow (BM) lineage-negative (Lin−)/liver sinusoidal endothelial cells (LSECs) co-cultures. NAC were collected, at days 21 and 41, from BM Lin−/LSEC co-cultures and analyzed for their expression of CD19 by flow cytometry (
Effect of LSEC-conditioned media on B lymphopoiesis
Because B lymphopoiesis was generated from BM Lin−/LSEC co-cultures in the absence of exogenous cytokines, we investigated the capacity of LSEC to produce B-cell growth factors associated with the survival and proliferation of B cells. For this purpose, we evaluated the effect of LSEC-conditioned media on NAC harvested from BM Lin−/LSEC co-cultures. B220+ cells were detected in NAC fraction after 21 days of culture in the presence of LSEC-conditioned medium, suggesting the presence of soluble factors that promote the survival of B cells in this conditioned medium (Fig. 5A). Morphological studies showed the presence of lymphoid-like cells (Fig. 5B). NAC from BM Lin−/LSEC co-cultures were unable to survive after 7 days of culture in the presence of regular medium (data not shown).
Liver sinusoidal endothelial cell (LSEC)-conditioned media promote B lymphopoiesis. Non-adherent cell (NAC) collected from bone marrow (BM) lineage-negative (Lin−)/LSEC co-cultures were cultured in the presence of LSEC-conditioned media. After 21 days of culture, the cells were collected and analyzed by flow cytometry. (
Expression of B cytokines and VCAM-1 by LSEC
We investigated whether cytokines required for B-cell commitment, survival, and maturation are expressed by LSEC. We examined the expression of IL-7, SCF, and Flt-3L using RT-PCR, on total RNA extracted from LSEC, and obtained cDNA bands corresponding to the expected sizes for IL-7, Flt-3L, and SCF (Fig. 6A). Based on evidence showing that VCAM-1 is involved in B-cell development [10 –13], we examined the expression of this molecule on LSEC. Our results show a highly constitutive expression of VCAM-1 on LSEC (Fig. 6B). Together, these results indicate that LSEC express molecules involved in regulating B-cell development.
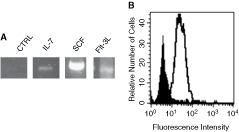
Liver sinusoidal endothelial cell (LSEC) constitutively express molecules required for B lymphopoiesis. Reverse transcription and amplification were performed as described in Materials and Methods using specific primers for IL-7, SCF, and Flt-3L. Constitutive expression for IL-7, SCF, and Flt-3L (
LSEC support the survival and proliferation of spleen B220+ cells
To determine whether LSEC have the capacity to support the survival and proliferation of spleen B220+ cells, NAC from splenocytes were seeded on LSEC monolayer in the absence of exogenous cytokines. From 35 days of co-culture, there was a significant increase in the number of NAC harvested from splenocytes/LSEC co-cultures (Fig. 7A). The production of NAC was maintained for >10 weeks, and the cell viability was >90% throughout this period (data not shown). Microscopic evaluation of splenocytes/LSEC co-cultures showed the presence of clusters of cells, indicating an active cell proliferation (Fig. 7B). Splenocytes seeded in the absence of LSEC and exogenous cytokines did not survive after first week (data not shown).

Liver sinusoidal endothelial cell (LSEC) promote the proliferation of splenocytes and the survival of B cells. Non-adherent cell (NAC) were collected from splenocytes/LSEC co-cultures and the cell number and viability were determined by trypan blue (
Finally, we examined the survival and functionality of spleen B cells. A high percentage of cells expressing the B-cell markers B220 and CD19 were found after >5 weeks in the splenocytes/LSEC co-cultures (Fig. 7C and 7D, respectively). These cells were detected in these co-cultures for >10 weeks (data not shown). Microscopic evaluation of NAC generated from splenocytes/LSEC co-cultures showed the presence of cells with lymphoid morphology (Fig. 7E). Further cytometric studies of the NAC obtained from splenocytes/LSEC co-cultures showed the presence of a very small number of sIgM+ cells (Fig. 8A). However, higher numbers of sIgM+ cells were detected in NAC generated from splenocytes/LSEC co-cultures treated with LPS for 4 days (Fig. 8B).

Lipopolysaccharide (LPS) stimulation increases the expression of IgM on non-adherent cell (NAC) harvested from splenocytes/liver sinusoidal endothelial cell (LSEC) co-cultures. Splenocytes/LSEC co-cultures were treated for 4 days with LPS. At the end of the co-culture, NAC were collected from untreated (
Discussion
In this work, we have provided further evidence about LSEC as a hematopoietic microenvironment by showing that these cells promote the survival and proliferation of primitive BM Lin− cells. These results also confirm other studies indicating that EC constitute specialized niches for HSC [22]. It was previously shown that LSEC promote hematopoietic differentiation to myeloid lineages [16]. Here, we provide evidence that LSEC also promote not only the survival, proliferation, and differentiation of primitive HSC to B-cell precursors (pre-pro-B, B220+CD19−), but also differentiation to more mature stages of B-cell development (pro-B/pre-B cells, B220+CD19+). While only few cells expressing the B-cell markers B220+CD19+ were detected as early as 7 days of BM Lin−/LSEC co-culture, higher numbers of these cells were detected after 3 weeks, and they persisted for >5 weeks of co-cultures. Together, these results indicate that LSEC promote B lymphopoiesis from HSC, and provide the first evidence that these cells constitute a B-cell microenvironment, in vitro.
There is evidence showing that sIgM+ B cells are produced during the first 6–8 days of culture from a population of pro-B cells enriched from BM [23]. Here, we show that small number of B cells generated in the BM Lin−/LSEC co-cultures expressed sIgM on their surface, but treatment of BM Lin−/LSEC co-cultures with LPS induced a significant increase in sIgM expression. The capacity of NAC collected from BM Lin−/LSEC co-cultures to survive and form colonies under stimulation with IL-7 constitutes further evidence of the functionality of the B cells generated in these co-cultures. We also show that LSEC promote not only the survival, but also the proliferation of spleen B220+ cells for >3 months. As with BM Lin−/LSEC co-cultures, increased numbers of B cells expressing sIgM were also detected in splenocytes/LSEC co-cultures treated with LPS. Together, these results indicate that LSEC induce and maintain functional precursors (B220+), which develop into mature and functional B cells (B220+CD19+IgM+). Transplantation experiments of B220+ cells generated from BM Lin−/LSEC co-cultures into irradiated congenic mice would allow to determine whether these cells are true precursors that are capable of developing into mature B cells.
The B-cell development has been reported using several stromal cell lines as in vitro microenvironments [24 –27]. Among them, the macrophage OP9 cell line constitutes one of the most efficient B-lymphopoietic microenvironment. It has been shown that the OP9 cell line promotes B lymphopoiesis from embryonic stem cells (ES) [28,29], but B-cell differentiation into functional B cells was observed only after adding IL-7 and/or Flt-3L to the ES/OP9 co-culture [28,29]. Similar results have been reported using other stromal cells, in which the appearance of B-lineage cells generally peaked at 4–6 weeks, and the full development of B cells again occurs only after addition of exogenous IL-7 [8,30 –32]. Importantly, we show here that B lymphopoiesis from BM Lin− co-cultured on LSEC monolayers occurred in the absence of exogenously added cytokines, suggesting that the LSEC produce soluble factors, which support and maintain B-cell development in vitro. This was confirmed by showing that LSEC-conditioned media promote the survival and differentiation of B220+ cells from NAC-harvested BM Lin−/LSEC co-cultures. Moreover, we show that LSEC express IL-7, which plays an important role in various stages of murine B-cell development including commitment, survival, proliferation, and maturation from HSC [7 –9]. It was previously shown that IL-7 is expressed in the liver [33]. Interestingly, this expression was detected within portal tracts, where LSEC are located [33]. Here we show that the LSEC also express Flt-3L and SCF, which together with IL-7 may exert a synergistic effect to induce B-cell differentiation from primitive BM Lin−cells [34,35].
It has been reported that B-cell development requires the relationship between cell adhesion molecules expressed on HSC (ie, VLA-4) and on BM stromal cells (ie, VCAM-1) [10 –13]. It is known that these molecules may play a fundamental role in B lymphopoiesis by promoting adhesion of early B-cell precursors to BM stromal cells [10 –12,36]. The capacity of LSEC to promote B lymphopoiesis from HSC could be related to the high constitutive expression of VCAM-1 on their surface, which may promote the adhesion of BM Lin− cells and early B-cell precursors to LSEC. This possibility is supported by the fact that antibodies against VCAM-1 on LSEC diminished the generation of cells from BM Lin−/LSEC co-cultures (data not shown). Together, these results suggest that LSEC may support B lymphopoiesis, not only by producing B-cell-stimulating cytokines, but also by promoting adhesion to HSC.
In addition to the presence of HSC [37,38], there is also evidence of cells expressing lymphoid and myeloid markers in normal adult human liver [16,39,40], suggesting that lymphoid and myeloid cells may differentiate and maturate locally. There are also various reports suggesting that B lymphopoiesis may occur in the adult liver. Likewise, extramedullary liver B lymphopoiesis has been shown in osteopetrotic mice, which are known to have abnormalities in BM hematopoiesis, mainly related to B lymphopoiesis [41]. Interestingly, there is also evidence showing T-cell maturation in the liver [42]. Based on the phenomenon that BM-resident hematopoietic stem and progenitor cells constitutively egress into the blood, and traffic to multiple peripheral organs [43], a recent work has shown the presence of hematopoietic clonogenical progenitors in the liver [44]. Thus, it is possible that various blood lineages, including B cells, differentiate from resident or circulating HSC in the liver, which could serve to maintain a local production of liver-resident B cells, or may be important for immunosurveillance [44].
An important question derived from our results is whether B cells generated from BM Lin−/LSEC co-cultures arose from HSC, which continually produce B-cell precursors. This possibility is supported by the fact that LSEC maintained HSC (Sca+/c-kit+) for >8 weeks in cultures (data not shown). Another major question is whether the capacity of LSEC to promote B lymphopoiesis in vitro may serve in vivo as a B-lymphopoietic microenvironment under physiological or pathological conditions or when HSC are mobilized. We believe this could be possible because B lymphopoiesis occurs in the fetal liver [45 –47], suggesting the presence of a liver B-lymphopoietic microenvironment during the fetal life, which could persist in the adult. On the other hand, based on in vivo evidence that the LSEC serve as a specific hematopoietic niche when HSCs are mobilized, or during extramedullary hematopoiesis [16], it is possible that these cells may represent a B-cell liver niche. Thus, the liver could be an extramedullary site for B lymphopoiesis throughout life, and the LSEC may be the specialized niche to promote the differentiation and/or maturation of B cells from resident or circulating HSC, because they not only express cytokines (ie, IL-7, Flt-3L and SCF), but also express adhesion molecules (ie, VCAM-1) involved in B-cell development. However, further studies are needed to establish whether LSEC constitute the main liver B-lymphopoietic niche.
In summary, our results demonstrate that LSEC constitute an in vitro B-cell microenvironment. The LSEC not only promote all developmental transitions required for full B-cell development from primitive HSC cells, but also maintain the functionality of these cells. On the basis of the capacity of LSEC to support hematopoiesis, in vivo and in vitro, our results suggest the existence of specific HSC niches in the liver, located in the LSEC microenvironment, which could support HSC self-renewal, myelopoiesis, and B lymphopoiesis. Further studies of the LSEC microenvironment will improve our understanding of the molecules involved in regulating B-cell development from HSC.
Footnotes
Acknowledgments
The authors wish to thank Dr. Egidio Romano and Dr. Howard Takiff for critically reading of the manuscript. The authors wish to thank Jose Alonso for technical assistance. This work was supported by FONACIT grant G2005000405.
Author Disclosure Statement
No competing financial interests exist.