
Abstract
Mesenchymal stem cells (MSCs) can differentiate into multiple mesodermal cell types in vitro; however, their differentiation capacity is influenced by their tissue of origin. To what extent epigenetic information on promoters of lineage-specification genes in human progenitors influences transcriptional activation and differentiation potential remains unclear. We produced bisulfite sequencing maps of DNA methylation in adipogenic, myogenic, and endothelial promoters in relation to gene expression and differentiation capacity, and unravel a similarity in DNA methylation profiles between MSCs isolated from human adipose tissue, bone marrow (BM), and muscle. This similarity is irrespective of promoter CpG content. Methylation patterns of MSCs are distinct from those of hematopoietic progenitor cells (HPCs), pluripotent human embryonic stem cells (hESCs), and multipotent hESC-derived mesenchymal cells (MCs). Moreover, in vitro MSC differentiation does not affect lineage-specific promoter methylation states, arguing that these methylation patterns in differentiated cells are already established at the progenitor stage. Further, we find a correlation between lineage-specific promoter hypermethylation and lack of differentiation capacity toward that lineage, but no relationship between weak promoter methylation and capacity of transcriptional activation or differentiation. Thus, only part of the restriction in differentiation capacity of tissue-specific stem cells is programmed by promoter DNA methylation: hypermethylation seems to constitute a barrier to differentiation, however, no or weak methylation has no predictive value for differentiation potential.
Introduction
M
MSC-like cells in BM, adipose tissue, muscle, and other organs have been shown to share a common perivascular niche within the microvascular environment in their respective tissues [10,11]. A perivascular origin of MSCs is supported by the discovery that pericytes carry surface antigens common to MSCs in vivo, include progenitors of mesodermal cell types, individually exhibit multilineage potential, and do not form teratomas [11]. Nevertheless, whilst MSCs may originate from pericytes [11], not all pericytes are MSCs because precursors of hematopoietic and neuronal lineages also inhabit the perivascular compartment [12].
A common origin of MSCs predicts that they would be intrinsically similar regardless of their tissue of residence, but there is to date no molecular evidence for similarity or difference in the potential for expression of lineage-specific genes in progenitor cells from various tissues. Gene expression potential is regulated by epigenetic processes among which cytosine methylation in CpG dinucleotides and post-translational modifications of histones are best characterized [13]. Methyl groups on DNA create target sites for methyl-binding proteins that in general induce transcriptional repression by recruiting co-repressors [14]. DNA methylation is carried out by DNA methyltransferases that are responsible for replication-coupled maintenance methylation, or for de novo methylation. The relationship between DNA methylation and gene expression is intricate and evidence highlights promoter CpG content as a component of this complexity [15]. Whether promoter CpG methylation predicts gene activation and differentiation potential in adult precursor cells is unknown.
Here, we carried out a single-nucleotide resolution mapping of DNA methylation patterns in lineage-specific promoters in relation to gene expression and differentiation capacity, to unravel an epigenetic relationship between human adipose, BM, and muscle MSCs. We also show that MSCs are epigenetically distant from human embryonic stem cells (hESCs), hESC-derived mesenchymal cells (hESC-MCs), and hematopoietic progenitor cells (HPCs). We suggest that strong promoter DNA methylation predicts lineage restriction in adult progenitor cells, whereas no or weak methylation has no predictive value on transcription and differentiation capacity.
Materials and Methods
Cells
ASCs were purified from the stromal vascular fraction of liposuction material and cultured as described [16]. CD14CD34+ BMMSCs were isolated and cultured as described [7]. CD34+ HPCs (>97% purity assessed by flow cytometry) were isolated from human BM [17]. Muscle progenitor cells (MPCs) (CC-2580; Lonza;
Adipogenic, myogenic, and endothelial differentiation
For adipogenic differentiation, cells were cultured to confluency and stimulated for up to 3 weeks with 1-methyl-3 isobutylxanthine, dexamethasone, insulin, and indomethacin [6,20]. Cells were stained with Oil Red-O to visualize lipid droplets and staining quantified by A500 measurement of extracted Oil Red-O in triplicate cultures. For myogenic differentiation, MPCs were grown to 80% confluency and cultured for 6 days in DMEM/2% horse serum and 1% penicillin; ASCs, BMMSCs, and hESC-MCs were cultured to 70% confluency and stimulated for 5 weeks in DMEM/5% horse serum, 50 μM hydrocortisone, and 1% penicillin [21]. Nuclei were stained with Hemacolor® (Merck;
Reverse transcription PCR
Reverse transcription (RT)-PCR was performed from 0.5 μg total RNA (Qiagen RNeasy;
Bisulfite genomic sequencing
DNA was purified and bisulfite-treated using MethylEasy™ (Human Genetic Signatures; North Ryde, Sydney, Australia;

DNA methylation profile of adipogenic and myogenic promoters in mesenchymal stem cells (MSCs) and hematopoietic progenitor cells (HPCs). (
Results
MSCs from adipose tissue, bone marrow, and muscle show similar lineage-specific promoter methylation patterns albeit distinct differentiation capacity
We examined by bisulfite sequencing the CpG methylation profile of 3 adipogenic promoters (LEP, PPARG2, FABP4), 4 myogenic promoters (MYOG, MYOD1, PAX7, MYF5), and 1 endothelial promoter (CD31/PECAM1) in MSCs from adipose tissue (ASCs), bone marrow (BMMSCs), and skeletal muscle (MPCs) (Fig. 1A). We identified 4 of these promoters (LEP, MYOD1, PAX7, MYF5) as high CpG promoters (HCPs) and 4 as low CpG promoters (LCPs) (FABP4, PPARG2, MYOG, CD31) (Fig. 1A) [15].
In ASCs, adipogenic promoters were largely unmethylated regardless of CpG content (Fig. 1B and 1C), confirming earlier findings [23]. Among myogenic promoters, MYOG contained 34% methylated CpGs, while the MYOD1, PAX7, and MYF5 HCPs had under 10% methylation (Fig. 1B and 1C). All these promoters were also largely unmethylated in both BMMSCs and MPCs irrespective of CpG content (Fig. 1B). Notably, although the 4 CpGs within the FABP4 LCP were methylated in MPCs, in contrast to ASCs or BMMSCs (Fig. 1B), spreading of these CpGs over the 413 bp examined resulted in an essentially unmethylated region. Promoter hypomethylation in MSCs was not an artifact of culture because all LCPs examined were methylated in cultured primary keratinocytes, in contrast to MSCs (Fig. 1D), and cultured ASCs have been shown to globally retain methylation patterns of their freshly isolated uncultured counterparts [22 –24]. In addition, no adipogenic transcripts were detected by RT-PCR in ASCs, BMMSCs, and MPCs despite their hypomethylated state, and only MYF5, MYOD1, and MYOG were expressed in MPCs (data not shown). Therefore, for the promoters examined there was no correlation between global methylation or methylation status of a specific CpG and transcriptional activity, corroborating recent genome-wide data [15].
Next, we asked whether poor methylation in these promoters in MSCs also characterized other progenitor cell types, and examined CD34+ HPCs purified from BM. As anticipated, the MYOD1, PAX7, and MYF5 HCPs were unmethylated in HPCs; however all other promoters, including the LEP HCP, were hypermethylated compared to all MSC types (Fig. 1B and 1C). HPCs therefore segregate from MSCs by the hypermethylation of non-CpG-rich myogenic and adipogenic promoters, suggesting that hypomethylation of non-HCP mesodermal lineage promoters constitutes an epigenetic signature of MSCs.
To determine whether methylation profiles identified in MSCs reflected differentiation capacity, each cell type was induced to differentiate into adipocytes and myocytes in vitro. ASCs and BMMSCs differentiated efficiently into adipocytes based on lipid accumulation (Fig. 2A and 2B) and up-regulation of adipogenic genes (Fig. 2C). In contrast, MPCs showed very limited adipogenic differentiation (Fig. 2A and 2B). Additionally, 6 days of myogenic stimulation of MPCs elicited the formation of multinucleated cells, a hallmark of myogenesis (Fig. 2D; Day 6), and myogenin protein expression in 57% of the cells (Fig. 2E). MPCs cultured for 6 days in control proliferation medium did not form multi-nucleated cells (Fig. 2D; Day 6 control) although 27% (P = 0.002 relative to differentiated cells) expressed intranuclear myogenin (Fig. 2E). Contrary to MPCs, ASCs and BMMSCs stimulated for up to 5 weeks showed no sign of multinucleation (Fig. 2D), myogenin expression (Fig. 2E), or up-regulation of myogenic genes (data not shown), indicating a lack of myogenic differentiation under these conditions. Of note, the lack of myogenic commitment of BMMSCs was recently confirmed by the retention of a stromal phenotype upon co-culture with rat embryonic cardiomyocytes [25].
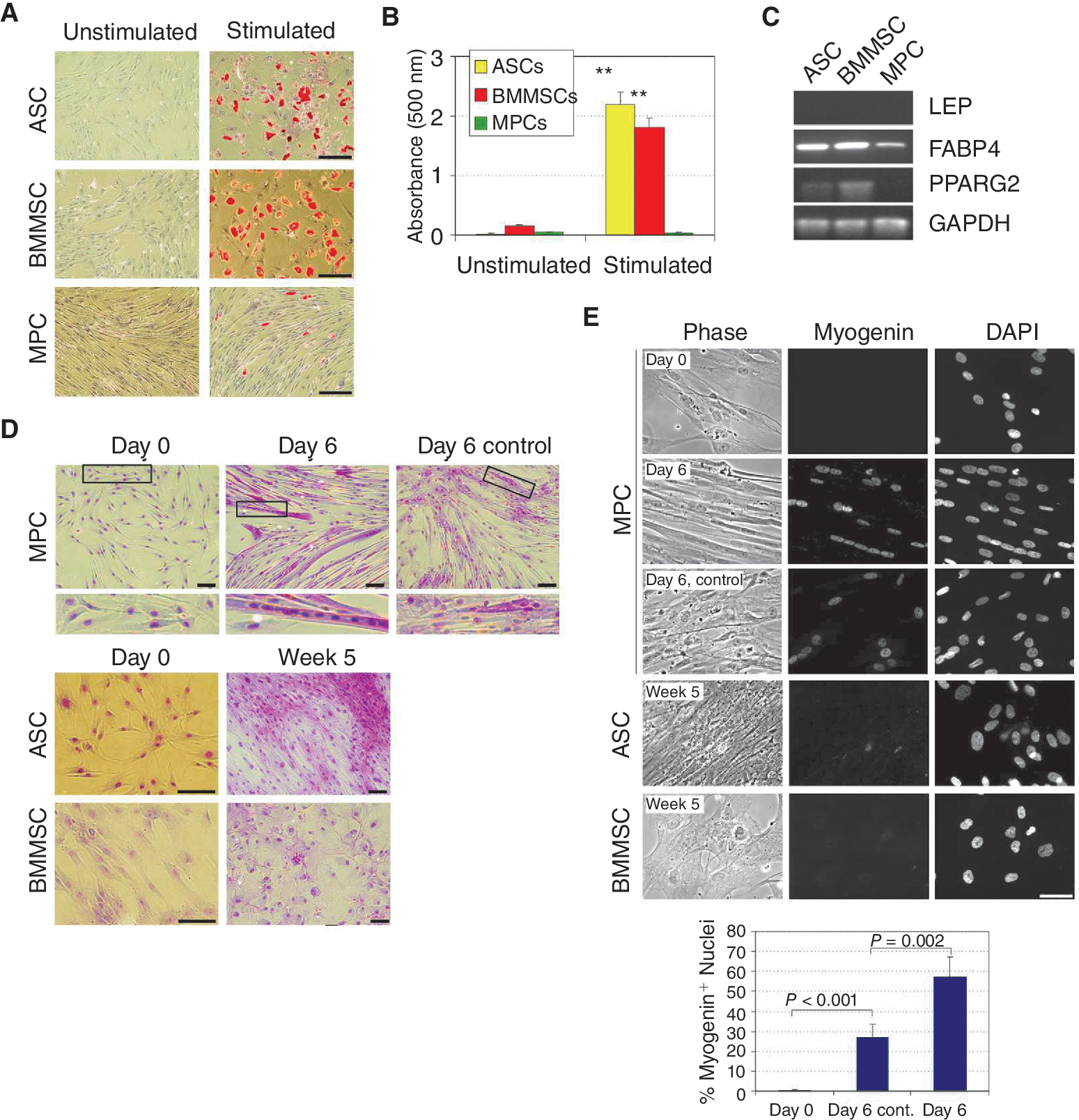
Adipogenic and myogenic differentiation of adipose stem cells (ASCs), bone marrow mesenchymal stem cells (BMMSCs), and muscle progenitor cells (MPCs). (
Based on transcriptional and phenotypic evidence, we suggest therefore that weak or no methylation of adipogenic and myogenic promoters in MSCs, regardless of CpG content, is uncoupled from the potential for transcriptional activation, and thus has no predictive value for differentiation potential. In contrast, promoter hypermethylation seems to predict lineage restriction.
Methylation states are maintained after MSC differentiation
To determine the extent of epigenetic commitment of MSCs to lineages predicted by their tissue of origin, we first compared methylation states of undifferentiated ASCs to adipocytes. Regardless of CpG content, extent and profiles of promoter methylation were similar in ASCs, adipogenic differentiated ASCs and in adipocytes [26] (Fig. 3A and 3B). Next, we found that FABP4, PPARG2, and MYOG methylation patterns were maintained after myogenic differentiation of MPCs, and similar to those identified in a muscle biopsy (Fig. 3C and 3D). The only trend detected was mosaic methylation around the TSS of LEP in differentiated MPCs and in muscle relative to undifferentiated cells, reflecting heterogeneity in these cell populations (Fig. 3C). Overall maintenance of promoter DNA methylation patterns after MSC differentiation and in differentiated cells strongly suggests that DNA methylation marks of differentiated cells are largely established at the progenitor stage.

Differentiation of adipose stem cells (ASCs) and BMMSCs maintains promoter DNA methylation profiles. (
Hypermethylation of the endothelial CD31 promoter in MSCs correlates with restriction in transcriptional activation potential
To reinforce the specificity of methylation profiles of adipogenic and myogenic promoters established so far, we examined a promoter not known to be implicated in adipo- or myogenic MSC differentiation. The endothelial CD31 proximal promoter was fully methylated in ASCs, BMMSCs, and MPCs, as well as in adipocytes and muscle (Fig. 4A and 4B). However, CD31 was unmethylated in HPCs both in the promoter and immediately downstream of the TSS, as in human umbilical vein endothelial cells, examined as positive control (HUVEC; Fig. 4A and 4B). To determine whether promoter methylation reflected a potential for CD31 gene expression, we elicited endothelial stimulation of MSCs and HPCs for 7 days in methylcellulose, and showed by RT-PCR that CD31 remained unexpressed in ASCs, BMMSCs, and MPCs, but was transcriptionally induced only in HPCs (Fig. 4C and 4D). Thus the unmethylated state of CD31 in HPCs is compatible with transcriptional activation, whereas strong methylation in ASCs, BMMSCs, and MPCs seems to limit its potential for activation.

CpG methylation status of the CD31 promoter correlates with potential for transcriptional activation upon endothelial stimulation. (
Lineage-specific promoters are hypermethylated in hESCs and hESC-derived mesenchymal cells
Mesenchymal cells can be derived from hESCs (hESC-MCs) and harbor many surface markers common to MSCs [20,27 –30]. hESC-MCs can give rise to osteogenic and chondrogenic cells in vitro but their adipogenic potential is variable [20]. To determine to what extent hESC-MCs are epigenetically related to bona fide MSCs, we examined methylation patterns in 2 hESC-MC cultures derived from 2 hESC lines.
First, we showed that in 2 undifferentiated hESC lines, in contrast to MSCs, all lineage-specific promoters examined were strongly methylated regardless of CpG content, while as expected the pluripotency gene OCT4 and NANOG promoters were unmethylated (Fig. 5A). These patterns were in contrast to those of ASCs and BMMSCs (Fig. 5B). These profiles did not result from contamination by feeder cells, which displayed distinct methylation patterns, and were nearly identical in undifferentiated embryonal carcinoma cells (Fig. 5A). This suggests that hypermethylation of these lineage-specific promoters constitutes a hallmark of cultured embryonic cells.

Human embryonic stem cells (hESCs) and hESC-MCs contain hypermethylated adipogenic, myogenic, and endothelial promoters. CpG methylation in indicated promoters in (
After derivation of MCs, LEP, FABP4, and PPARG2 remained strongly methylated, MYOG was demethylated and displayed a profile more similar to MSCs, while CD31 was mosaically demethylated upstream of and around the TSS, as well as in the first exon (Fig. 5C). OCT4 and NANOG were moderately methylated in hESC-MCs, as in ASCs and BMMSCs (Fig. 5C). Therefore, promoter-specific demethylation events upon MC derivation from hESCs result in methylation patterns that are distinct from hESCs and MSCs. Moreover, hESC-MCs failed to differentiate into adipogenic or myogenic cells and to up-regulate adipogenic or myogenic genes after stimulation (data not shown). Thus hypermethylation of adipogenic promoters in hESC-MCs correlates with the absence of adipogenic capacity; however, the unmethylated state of the MYOG LCP is not indicative of transcription or myogenic differentiation potential.
Discussion
We show here by bisulfite sequencing of lineage-specific promoters a common methylation signature between MSCs from human adipose tissue, BM, and muscle, which is independent of differentiation capacity. These methylation patterns however distinguish MSCs from HPCs and reflect a differential epigenetic programming of mesenchymal and hematopoietic progenitors. MSCs are also epigenetically distinct from pluripotent hESCs, in which in contrast to MSCs, lineage-specific promoters are strongly methylated. Such hypermethylation implies a programming of these cells for functions other than differentiation (eg, maintenance of pluripotency permitted by the unmethylated state of OCT4 and NANOG, or replication), when cultured in a proliferative state. We also suggest that whereas no or weak promoter methylation has no predictive value on differentiation potential, hypermethylation constitutes a differentiation barrier and may thereby predict lineage restriction.
The ontogeny of MSCs remains poorly understood; however, recent identification of the perivascular compartment as a niche for MSCs throughout the body points to a common origin of MSCs from various tissues [4,11]. The common promoter DNA hypomethylation signature of lineage-priming promoters in ASCs, BMMSCs, and MPCs identified here provides molecular “intrinsic” evidence substantiating the idea of a common MSC ancestor based on immunophenotypic and functional data [11]. To support this view, our preliminary analysis of pericytes reveals promoter methylation patterns similar to MSCs (A.L.S., I. Andersen, K.V., P. Kaur and P.C., unpublished data). Interestingly, cultured pericytes purified from human muscle do not express any known myogenic marker despite their myogenic potential [11]. Expression of these markers upon myogenic differentiation would be consistent with a permissive chromatin state on a hypomethylated DNA background on myogenic promoters, in line with our observations.
Mesenchymal progenitors derived from hESCs [27,28,30] may constitute a putative patient-specific therapeutically useful option, as they originate from an unlimited source of pluripotent cells and do not harbor the tumorigenicity of hESCs (see ref. [30] and references therein). hESC-MCs may differentiate into adipo-, osteo-, and chondrogenic cell types [27,28,30], but not all lines are able to do so [20]. Based on our studies, we propose that adipogenic potential of some of these cultures is precluded by the hypermethylated state of key adipogenic promoters. The present work indicates that epigenomes of hESC-MCs differ from that of bona fide MSCs, which reflects their distinct differentiation capacity [20] and may restrict any therapeutic use of hESC-MCs.
A consistent observation when analyzing methylation profiles by bisulfite sequencing is mosaicism between the genomic fragments examined. This mosaicism reflects variations in the methylation state of specific CpGs between alleles and/or cells in a population; it also suggests heterogeneity within stem cell compartments, supporting mounting evidence. Indeed, clonogenic MSCs constitute a mix of progenitors [9], and within the hematopoietic compartment, cells with a uniform immunophenotype have different abilities to produce lineage-specific descendants [31]. The differentiation potential of these cells has been proposed to be an “intrinsic property” [32] of these cells. This property is presumably established by their epigenome, as their myeloid or erythroid propensities are retained after transplantation [31]. It should be nonetheless mentioned that heterogeneity in stem cell populations may alternatively correlate with spatial localization and signals received from heterogeneous microenvironments. Whether distinct microenvironments within stem cell niches influence epigenetic states remains to be explored. The known links between pathological states such as viral infection, bacterial infection, or cancer and epigenetic makeup, in particular, DNA methylation [33] suggest so.
Despite a common promoter methylation signature among MSCs, their differentiation capacities differ, indicating that the relationship between promoter DNA methylation and gene activation or differentiation potential is not straightforward. First, we find that HCPs are unmethylated even when inactive, corroborating the view that HCPs are protected from methylation regardless of expression [15]. Second, none of the genes examined, irrespective of promoter CpG content, are expressed in undifferentiated ASCs or BMMSCs despite their hypomethylated state. Third, and importantly, methylation of the promoters examined here is unaltered after MSC differentiation and transcriptional activation [23]. This finding extends recent reports of overall stability of methylation profiles after terminal differentiation of mouse ESC-derived neuronal precursors in vitro [34,35], and argues that promoter methylation patterns of differentiated cells are already largely established at the progenitor stage.
Our observations highlight a functional significance of lineage-specific promoter DNA methylation in stem cell function and identity, yet this significance is enhanced by the epigenetic context brought about by histone modifications. No or weak promoter methylation does not imply transcriptional activity, and although hypomethylation constitutes a transcriptionally permissive chromatin state [36], it is not a predictor of gene activation potential or stem cell differentiation capacity. Transcriptional repression of unmethylated promoters is likely mediated by other mechanisms such as Polycomb-mediated trimethylation of H3 lysine (K)27 [35,37,38]. In line with this view, the hypomethylated LEP, FABP4, PPARG2, and MYOG promoters have been shown to be trimethylated on H3K4 and H3K27 in undifferentiated ASCs [39]. In contrast, we argue that strong promoter methylation constitutes a barrier to differentiation, even though methylation of LCPs is not necessarily inhibitory [15]. Consequently, on hypermethylated promoters, repressive histone marks may play a less prominent role on transcriptional repression. Examination from online data [40] of the histone methylation state of the lineage-specific promoters studied here in hESCs interestingly indicates that the hypermethylated LEP, FABP4, PPARG, MYOG, and CD31/PECAM1 promoters (Fig. 5C) are neither trimethylated on H3K4 nor H3K27. This finding is consistent with CpG hypermethylated sequences being antagonistic to H3K4 methylation [41] and argues that silencing of these promoters in hESCs is mediated by DNA methylation rather than, for example, Polycomb-mediated H3K27 methylation. In contrast, MYOD1, MYF5, and PAX7 CpG-rich promoters harbor the “bivalent” H3K4me3 and H3K27me3 marks, in agreement with Polycomb-mediated repression of these genes.
Integration of analyses of embryonic, pluripotent, and multipotent cell types points to complementary functions of DNA methylation and additional epigenetic factors in the regulation of stem cell differentiation potential. DNA methylation state may create a permissive or repressive environment underlying a fine-tuned control of gene activation by histone modifications, association with histone variants [42], and binding of transcriptional regulators.
Footnotes
Acknowledgments
We thank Gisele Bonne (Institut de Myologie, Paris) for the muscle biopsy, Aboulghassem Shahdadfar (Rikshospitalet, Oslo) for HPCs and BMMSCs, and Ingrid Andersen for help with bisulfite sequencing. This work was funded by the Swedish Research Council and the Petrus and Augusta Hedluinds Foundation to L.A.-R., the ERC program of the National Science Foundation Contract grant EEC-9731643 to S.L.S., and by the Research Council of Norway, the Norwegian Cancer Society and Health South-East Norway to P.C.
Author Disclosure Statement
S.L. Stice is founder and consultant for Aruna Biomedical, Inc.