
Abstract
Human induced pluripotent stem cells (iPSCs) hold promise as a source of adult-derived, patient-specific pluripotent cells for use in cell-based regenerative therapies. However, current methods of cell culture are tedious and expensive, and the mechanisms underlying cell proliferation are not understood. In this study, we investigated expression and function of iPSC integrin extracellular matrix receptors to better understand the molecular mechanisms of cell adhesion, survival, and proliferation. We show that iPSC lines generated using Oct-3/4, Sox-2, Nanog, and Lin-28 express a repertoire of integrins similar to that of hESCs, with prominent expression of subunits α5, α6, αv, β1, and β5. Integrin function was investigated in iPSCs cultured without feeder layers on Matrigel or vitronectin, in comparison to human embryonic stem cells. β1 integrins were required for adhesion and proliferation on Matrigel, as shown by immunological blockade experiments. On vitronectin, the integrin αvβ5 was required for initial attachment, but inhibition of both αvβ5 and β1 was required to significantly decrease iPSC proliferation. Furthermore, iPSCs cultured on vitronectin for 9 passages retained normal karyotype, pluripotency marker expression, and capacity to differentiate in vitro. These studies suggest that vitronectin, or derivatives thereof, might substitute for Matrigel in a more defined system for iPSC culture.
Introduction
H
Cells of all types interact with the ECM via integrins, multifunctional transmembrane receptors made up of α- and β-subunits, which are required for adhesion and proliferation in many cell types [9 –11]. Undifferentiated hESCs express a subset of integrins [3,5,12 –15], some of which may be up- or down-regulated during hESC differentiation [16,17], indicating the potential importance of integrins in maintaining an undifferentiated hESC state. Different hESC lines have been reported to express high levels of integrin subunit proteins α5 [3,6,15], α6 [3,7,15,17], αv [7,15], β1 [3,6,7,15], and β5 [7], as well as corresponding mRNAs [12,13]. The integrin α6β1 interacts with laminins, which are abundant in Matrigel [11]. α5β1 and αvβ1 interact with fibronectin [11] and αvβ5 and αvβ1 interact with vitronectin, both ECM proteins that contain arginine-glycine-aspartate (RGD) motifs [11,18,19]. hESCs can be successfully cultured for several passages on laminin [3,14] or vitronectin [7] alone, as well as on a combination of collagen IV, fibronectin, laminin, and vitronectin [4], although the most promising of these defined substrates may be vitronectin [7]. New, synthetic matrices containing adhesive peptides are also being explored [20 –22]. A recent study demonstrated key roles for integrins in adhesion and proliferation of hESCs on matrix substrates [7].
iPSCs have been successfully cultured on Matrigel [1,2], but iPSC culture on a defined matrix, and the quantitative characterization of functional iPSC integrins, in comparison with hESCs, have not been reported. In this study, we investigated integrin expression and function in iPSC lines generated by Yu et al. [1] via lentiviral expression of Oct-3/4, Sox-2, Nanog, and Lin-28. Expression of key integrins and their roles in adhesion, survival, and growth were analyzed using feeder-free Matrigel or vitronectin substrates. Furthermore, a fully defined, feeder-free culture system using vitronectin with mTESR1 media [4] allowed for long-term culture of iPSCs.
Materials and Methods
Cell lines and culture
hESC lines H9 and H14 [23] (WiCell Research Institute, Madison, WI,
Immunofluorescence
iPSCs were seeded onto chambered slides (Thermo Fisher Scientific) coated with Matrigel. Slides were washed with phosphate-buffered saline (PBS) pH 7.4, fixed with 4% para-formaldehyde in 0.1 M sodium cacodylate buffer, pH 7.4, for 15 min at 4°C, and stored in PBS at 4°C. Slides were washed with PBS, blocked with 1% bovine serum albumin (BSA; Sigma-Aldrich) with 0.1% IGEPAL (Sigma-Aldrich) and 1% goat or donkey serum (Sigma-Aldrich) in PBS for 1 h at 4°C. Slides were then incubated with primary antibodies overnight at 4°C: monoclonal mouse anti-α5 integrin (SAM-1, 20 μg/mL; Abcam, Cambridge, MA,
Western blot analysis
To prepare lysates, iPSCs were cultured on Matrigel with CM in 6-well tissue culture plastic plates (for at least 3 passages but <10 passages) and washed with PBS, each well treated with 25 μL lysis buffer containing 50 mM Tris-Cl pH 7.6, 150 mM NaCl, 5 mM EDTA, 1% IGEPAL, 0.2% SDS, and EDTA-free protease inhibitor cocktail set V (Calbiochem, La Jolla, CA,
Adhesion assays
Undifferentiated hESCs or iPSCs were cultured on Matrigel (except for Supplementary Fig. 5 (Supplementary materials are available online at
Proliferation assays
Undifferentiated hESCs or iPSCs maintained on feeder fibroblasts (to duplicate routine culture conditions) were manually dissected, trypsinized using 0.05% Trypsin-EDTA (Invitrogen) for 3 to 5 min, then treated with maintenance media containing 10% FBS, to generate single-cell suspensions. Because the cells in this assay were allowed to adhere to the plates over a 24-h period before integrin function-altering antibodies were added, the effects of EDTA on integrin function were thought to be insignificant, as this assay is standard [7]. After washes to remove residual FBS, trypsinized cell solutions were seeded in 4 flat-bottom 96-well tissue culture-treated plates at ∼32,000 cells per well (1.0 × 105 cells/cm2) in CM, in triplicate per condition. CM was used to duplicate standard culture conditions, as described above. Wells were previously coated with either Matrigel, vitronectin, or 1% BSA (as above). Twenty-four hours after seeding the cells, one plate was stained using a Live/Dead Viability/Cytotoxicity Kit (Invitrogen) and Hoechst 33342 (2 μg/mL; Invitrogen) and immediately imaged using an Olympus IX70 fluorescent microscope (Olympus). Remaining plates had integrin antibody inhibitors or activators added to the wells of cells in fresh CM as required. One plate was stained and imaged as described above 48, 72, and 96 h after seeding the cells, and antibodies added fresh to the remaining plate(s) until all 4 plates were imaged. Integrin function activating or inhibiting antibodies used were the same as in the adhesion assays. Images were obtained as described above. Live cells per field area were scored using ImageJ software (National Institutes of Health). Results were normalized to positive control cells on Matrigel- or vitronectin-coated wells without antibodies.
Vitronectin purification
Vitronectin was purified according to established protocol [24] except for the following modifications: After serum from pooled normal human plasma with sodium citrate (Innovative Research, Southfield, MI,
Karyotype analysis
Karyotyping of H9 (p74) and iPS(IMR90)-3 (p66) was performed by Cell Line Genetics (Madison, WI,
Results
iPSC integrin expression
To better inform development of defined substrates for iPSC culture, we characterized expression of integrin subunits. First, microarray data generated by Yu et al. [1] were analyzed to compare iPSC and hESC integrin mRNA expression (Supplementary Fig. 2). Integrins α3, α5, α6, α7, αV, β1, and β5 were detected reliably above background in 5 hESC lines and 9 iPSC lines generated using Oct-3/4, Sox-2, Nanog, and Lin-28 [1]. mRNAs encoding subunits that make up major integrin receptors for laminin (α6β1), fibronectin (α5β1), and vitronectin (αVβ5) were abundantly expressed on both iPSCs and hESCs, as has been reported previously for additional hESC lines [3,6,7,12 –15]. Because of their importance in other cell types, and because differences in physical routine handling or tissue culture history could cause changes in integrin expression of our cells compared to Yu et al. [1], we further examined the expression of these integrin subunit proteins in the iPSC lines we cultured. iPSCs and hESCs were grown on Matrigel with CM to prevent feeder contamination and duplicate standard feeder-free culture conditions. Western blot analysis showed that 3 iPSC lines, iPS(IMR90)-3, iPS(IMR90)-4, and iPS(foreskin)-2, expressed the integrin subunits α5, α6, αv, β1, and β5 (Fig. 1A). Expression in these lines was similar but not identical, with most approximate levels within 2-fold of hESC line H9. However, iPS(foreskin)-2 cells expressed higher levels of α5 and β5 subunits. We also investigated the expression of these integrin subunits in the iPSC lines using immunocytochemistry. We did not further investigate hESC integrin expression as it has been extensively reported previously [3,6,7,12 –15]. Immunocytochemistry confirmed the integrin expression in undifferentiated, Oct-3/4-positive iPSCs grown on Matrigel (Fig. 1C). The round undifferentiated cells have the characteristically small ratio of cytoplasmic to nuclear volume, sometimes resulting in faint integrin staining. iPSC colonies were often surrounded by differentiated fibroblast-like cells. These differentiated (Oct-3/4-negative) cells also expressed integrin subunits, sometimes staining more heavily (eg, β1 in iPS[IMR90]-3 and αv in iPS[IMR90]-4). However, because the amount of undifferentiated hESCs and iPSCs relative to surrounding differentiated cells (which were sometimes completely absent) was so great, this most likely did not significantly impact our western blot integrin expression analysis; hESCs on Matrigel have been previously used to characterize integrin expression in undifferentiated cells as well [3,13,15].
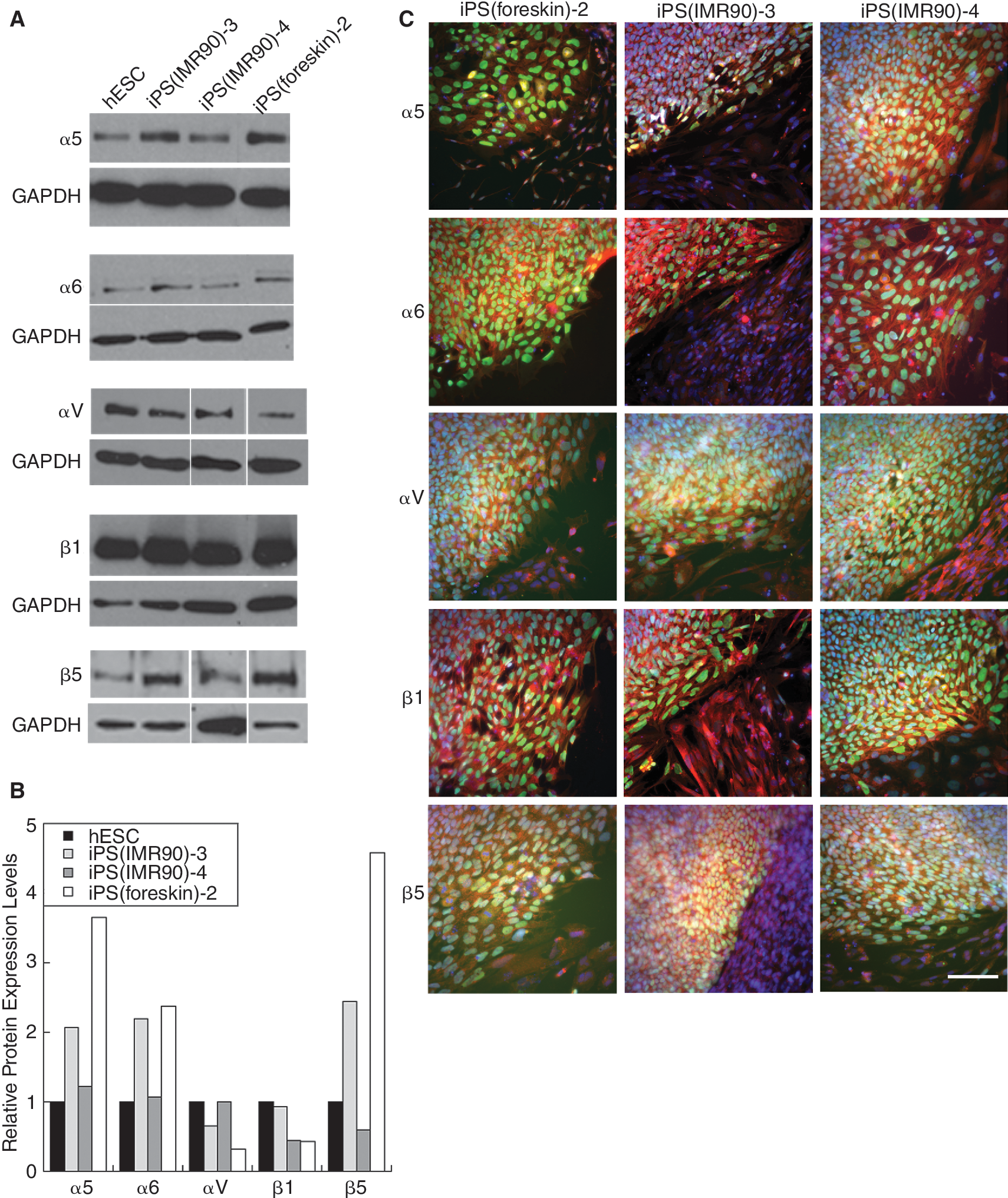
Induced pluripotent stem cell (iPSC) integrin expression. Expression of integrin subunits α5, α6, αv, β1, and β5 in 3 iPSC lines and the human embryonic stem cell (hESC) H9 line through western blot analysis (
β1 integrins are required for iPSC and hESC adhesion to Matrigel
We next investigated the function of these integrins in adhesion to Matrigel and vitronectin, surfaces that have been shown to support hESC culture. To determine the role of β1 integrins in adhesion to Matrigel, H9, iPS(IMR90)-3, or iPS(foreskin)-2 cells were incubated in Matrigel-coated wells in the presence of a β1 integrin inhibiting antibody, a β1 activating antibody, an isotype antibody control, or no antibodies. When the β1 inhibiting antibody was added, all lines tested showed a significant decrease in adhesion relative to the control with no antibodies added (Fig. 2A), though no lines showed any significant increase in the presence of the β1 activating antibody. Adhesion assays on Matrigel were also performed with α5 activating and inhibiting antibodies and an α6 inhibiting antibody (Supplementary Fig. 3). No significant effect was observed on the adhesion of H9 or iPS(IMR90)-3 cells in the presence of the α5 and α6 antibodies, separately or together, even with greater concentrations of inhibiting antibody (20–50 μg/mL, data not shown). These results indicate that β1 integrins are required for hESC and iPSC adhesion to Matrigel, but the exact combination of β1 heterodimers involved has not yet been identified (see discussion).
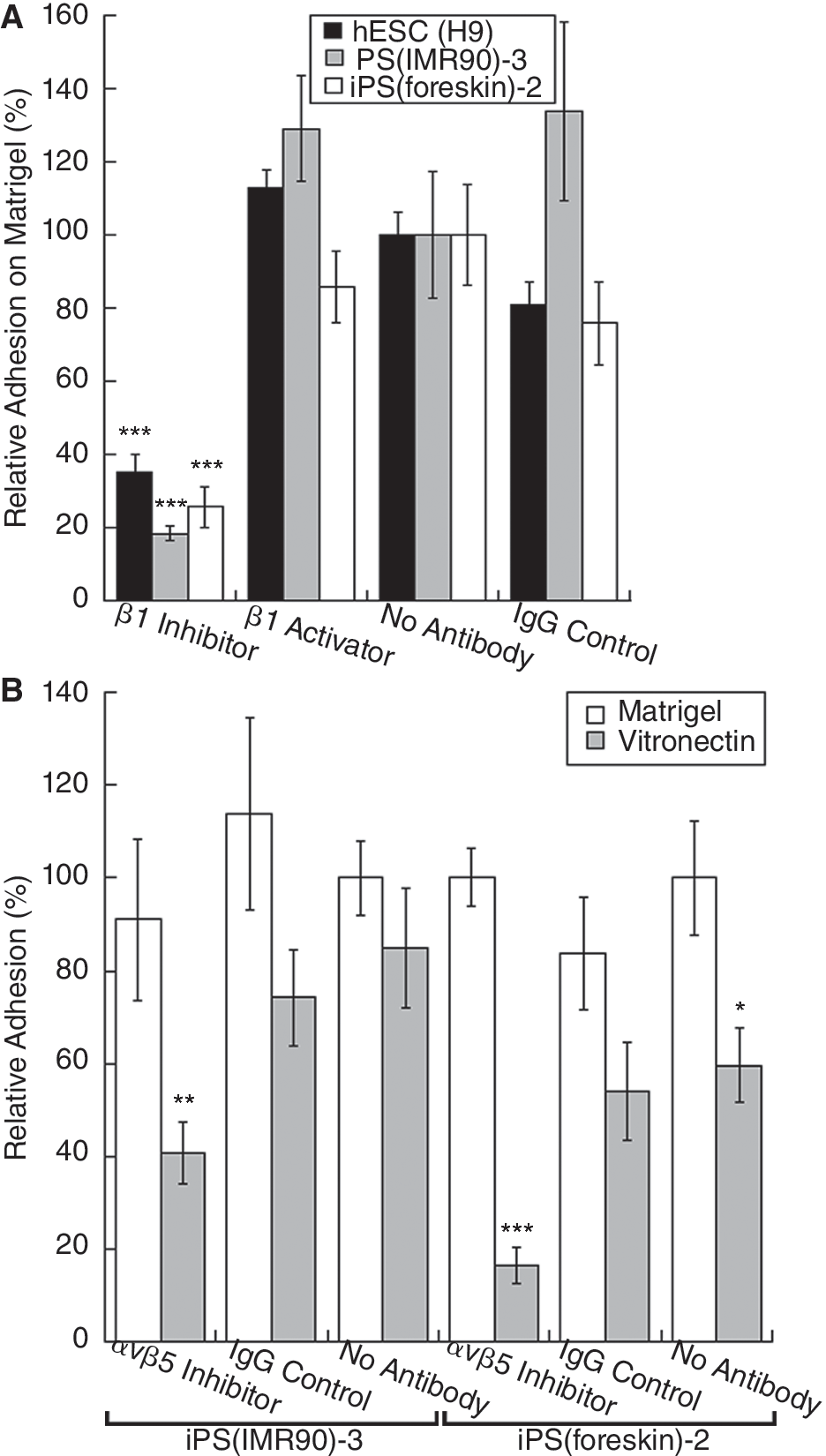
Function of β1 and αvβ5 integrins in induced pluripotent stem cell (iPSC) adhesion to Matrigel and vitronectin. Cells were seeded on Matrigel with β1 inhibiting or activating antibodies or an isotype control antibody (
αvβ5 integrin heterodimer is required for iPSC adhesion on vitronectin
Vitronectin was recently shown to be a defined substrate system that, with CM or mTESR1 media, a defined hESC media [4], allows for the culture of hESCs with retention of a normal karyotype and pluripotency [7]. This recent study also suggested that vitronectin promotes hESC growth primarily through the αvβ5 integrin heterodimer [7]. Investigations with culturing iPSCs on vitronectin have not been reported. To test the function of the αvβ5 heterodimer in iPSC adhesion to vitronectin, we performed an adhesion assay with iPS(IMR90)-3 and iPS(foreskin)-2 cells on vitronectin with an αvβ5 function inhibiting antibody. In both iPSC lines, the addition of the αvβ5 antibody significantly decreased cell adhesion relative to the no antibody control (Fig. 2B). Blockade of β1 did not affect adhesion to vitronectin (Supplementary Fig. 4). In contrast, the αvβ5 inhibiting antibody did not decrease adhesion to Matrigel (Fig. 2B); this is expected as Matrigel does not contain detectable vitronectin [5 –8]. In these experiments, adhesion was slightly higher on Matrigel than vitronectin. Because iPSCs were cultured on Matrigel immediately prior to the assay, it is possible that they developed a preference for Matrigel. To test this, we transitioned iPSCs from MEFs to vitronectin we purified, and cultured them for 2 passages (16 days) before performing an adhesion assay. The vitronectin-cultured cells were seeded onto Matrigel or vitronectin. These cells showed very similar levels of adhesion on both Matrigel and vitronectin (7.6% variance) (Supplementary Fig. 5).
β1 integrin subunit is required for iPSC and hESC proliferation on Matrigel
Next, we investigated β1 integrin subunit involvement in iPSC and hESC proliferation on Matrigel. To distinguish effects on proliferation versus adhesion, cells were seeded first and allowed to establish adhesion before addition of integrin function inhibiting or activating antibodies at 24, 48, and 72 h post-seeding. Live cells were imaged every 24 h over a period of 96 h. The anti-β1 inhibiting antibody significantly decreased the number of live cells observed in the H9 and iPS(IMR90)-3 lines, which continued to decrease over the following 48 h (Fig. 3A and 3B). The iPS(foreskin)-2 line response was delayed, with no significant decrease in the number of live cells observed with the addition of the anti-β1 until 96-h post-seeding (Fig. 3C), when a very distinct decrease in the number of live cells was seen. This shows some variation between the iPSC lines tested, though variations have been previously reported [1,8], and may be related to inherent differences between the iPS(IMR90)-3 and iPS(foreskin)-2 lines, such as the maturity level and tissue of origin of the donors cells (fetal lung fibroblasts and mature foreskin fibroblasts, respectively). As with the adhesion assays, the addition of the β1 activating antibody did not have a significant effect. To test for an αvβ5 role in iPSC proliferation on Matrigel, the αvβ5 inhibiting antibody was added to iPS(IMR90)-3 cells grown on Matrigel, but no decreased proliferation was observed (Fig. 3D).
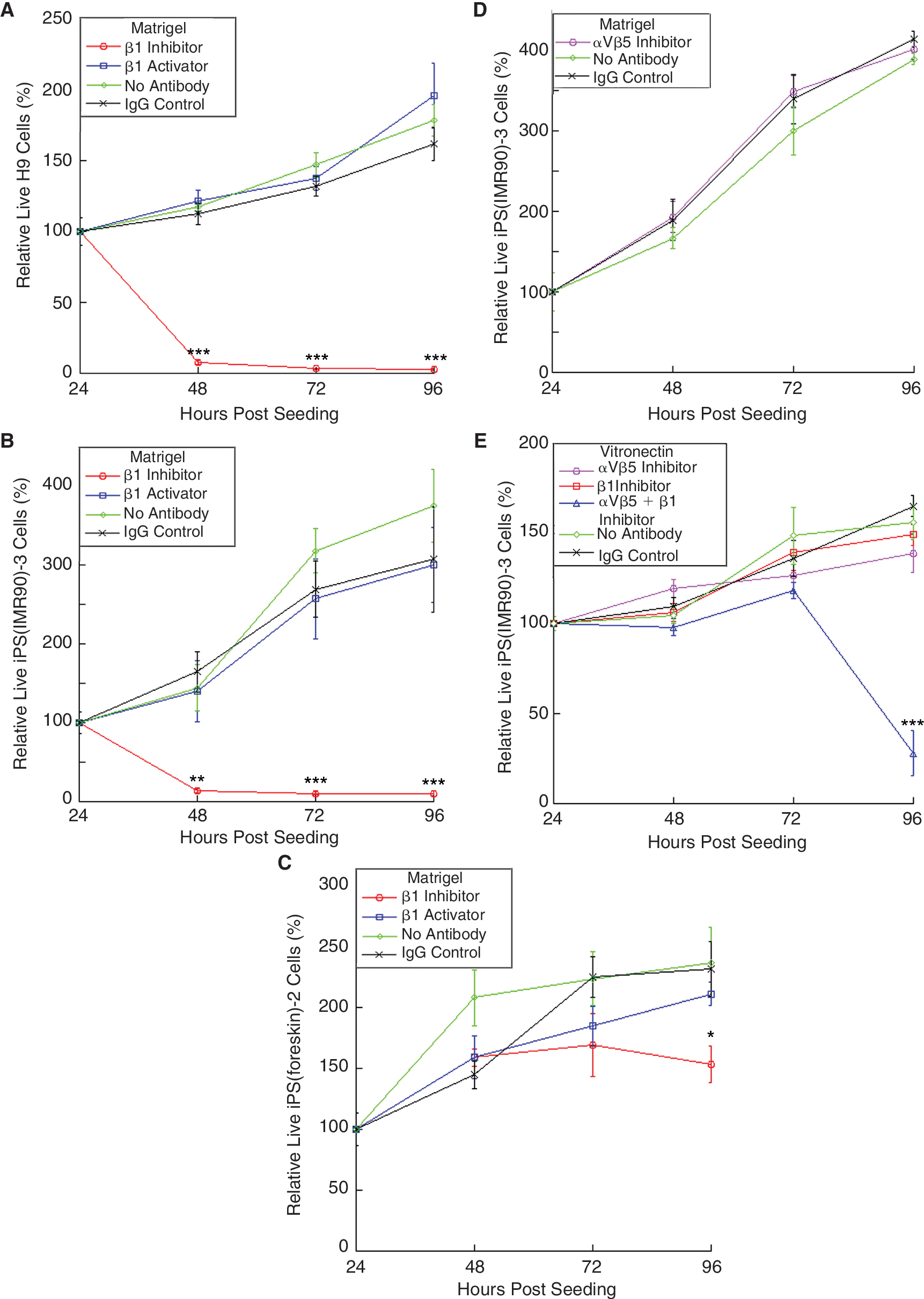
Integrin-mediated hESC and iPSC proliferation on Matrigel and vitronectin. hESC H9 (
αvβ5 and β1 integrins mediate iPSC proliferation on vitronectin
After observing that αvβ5 was required for adhesion to vitronectin, we investigated its role in proliferation of iPSCs on vitronectin, focusing on the iPS(IMR90)-3 line because it was the most robust. Surprisingly, no significant decrease in iPS(IMR90)-3 proliferation was observed with the addition of the αvβ5 inhibiting antibody on vitronectin at any point over the 96-h proliferation assay (Fig. 3E). This lack of effect was also seen with the iPS(foreskin)-2 line (data not shown). Likewise, anti-β1 did not affect iPS(IMR90)-3 cell proliferation (Fig. 3E). However, when anti-β1 was combined with anti-αvβ5, a significant decrease in proliferation was observed at 96-h post-seeding (Fig. 3E). iPSCs (previously cultured on MEFs) were observed to proliferate slightly better on Matrigel relative to vitronectin; the doubling rate on Matrigel (37 h) was 1.4 times faster than on vitronectin (54 h).
Long-term growth of hESC and iPSC on vitronectin
It has been shown that some hESC lines can be maintained for at least 8 passages on vitronectin in mTESR1 media and retain normal karyotype and pluripotency [7]. We tested whether WiCell hESCs, the most commonly used hESC lines, which were not previously examined, and iPSCs could likewise be maintained on vitronectin. iPS(IMR90)-3 and H9 cells previously cultured on feeders or Matrigel and H14 cultured on feeders were transitioned to vitronectin for 3 or 4 passages with CM and then imaged (Fig. 4A). These cells displayed normal colony morphology, appearing similar to the same cells grown on Matrigel with CM (Supplementary Fig. 6A), sometimes with fibroblast-like cells at the colony edges. No correlation between the original culture system (either from feeders or Matrigel) and morphological appearance on vitronectin was observed. Next, iPS(IMR90)-3 and H9 cells were cultured on vitronectin with mTESR1 media, together creating a defined, feeder-free system, without CM from feeders. H9 cells were cultured for 10 passages on vitronectin and iPS(IMR90)-3 cells for 9 passages. Controls were performed on Matrigel with mTESR1 media using the same cells and for the same duration. iPSC and hESC colonies grown on vitronectin (Fig. 4B) appeared morphologically similar to those grown on Matrigel (Supplementary Fig. 6B). After 9 or 10 passages on vitronectin, all were found to have a normal karyotype (Fig. 4C and Supplementary Fig. 6C). To confirm that the cells retained hESC marker expression, immunocytochemistry was performed. iPSC colonies stained positive for the hESC pluripotency surface markers SSEA-4, TRA-1-81, and TRA-1-60, and transcription factor Oct-3/4 (Fig. 4D and Supplementary Fig. 6D). To analyze differentiation potential, cells were induced to differentiate via embryoid body formation and labeled for markers from the 3 germ layers. All samples had cells that stained positive for βIII-tubulin (ectoderm), smooth muscle actin (mesoderm), and α-fetoprotein (endoderm) (Fig. 4E and Supplementary Fig. 6E). Additionally, we investigated whether hESCs spontaneously differentiate more on vitronectin or Matrigel. hESCs cultured on feeder fibroblasts were seeded onto vitronectin, Matrigel, or a 1:1 mixture of vitronectin and Matrigel. No significant difference was observed in the percent of undifferentiated colonies that arise, as determined by the mean Oct-3/4 and Sox-2 expression (Supplementary Fig. 7), which matches our observations that hESCs and iPSCs have similar amounts of differentiated cells over multiple passages on Matrigel or vitronectin.

iPSC and hESC long-term culture on vitronectin. hESC and iPSC lines, previously cultured on feeder fibroblasts or Matrigel, were cultured on vitronectin for 3 or 4 passages with CM and representative images taken (
Discussion
In this study, we show that integrin expression profiles are similar between hESCs and iPSCs, and that integrins play central roles in adhesion and proliferation of both hESCs and iPSCs in feeder-free systems. Specifically, we report that iPSC lines express α5, α6, αV, β1, and β5 integrin subunits, which multiple hESC lines have also been reported to express [3,6,7,12 –15]. However, not all iPSC lines are identical; several show significantly higher expression of some integrin mRNAs (eg, iPS(IMR90)-5, Supplementary Fig. 2) and integrin proteins (eg, iPS(foreskin)-2, Fig. 1A). These lines were reprogrammed from adherent cells (foreskin fibroblasts and fetal lung cells) with high integrin expression, and it is possible that some have retained this feature of the parental cell type. In addition, our data show that β1 integrins play a key role in adhesion and proliferation of both hESCs and iPSCs on Matrigel, while the αvβ5 heterodimer is important in iPSC vitronectin adhesion, but both αvβ5 and β1 are important for proliferation on vitronectin. Furthermore, we show that both WiCell hESCs and iPSCs can be successfully cultured long-term on vitronectin, with retention of normal karyotype and pluripotency. These conditions may form the basis of a well-defined feeder-free system.
While Matrigel adhesion was blocked by anti-β1, it was not affected by blockade of α5 or α6 integrin subunits, and the identity of β1 heterodimers involved remains to be determined. It is surprising that inhibition of the α6 integrin subunit did not result in decreased cell adhesion to Matrigel because of the high laminin content in Matrigel [3,8]. This could indicate that other β1 integrins are important for hESC and iPSC adhesion to Matrigel, such as α2β1, α3β1 [3,7,12,14], α7β1 [12,14], and α11β1 [7], which all interact with components of Matrigel: laminin-111 (α2β1, α7β1), and/or collagen IV (α2β1, α3β1, α11β1). However, we found α2 and α11 to be expressed at very low levels in iPSCs (Supplementary Fig. 2). Although the microarray data show expression of α3 and α7, previous studies have reported much lower mRNA and protein expression levels of these subunits in hESCs [3,7,12,14] than the subunits investigated in this study. It is possible that inhibiting the function of any single β1-dimerizing α integrin subunit, or even multiple α subunits, will not significantly inhibit hESC or iPSC adhesion or proliferation on Matrigel due to the ability of the β1 subunit to form laminin- and collagen-binding integrin heterodimers with several different α subunits. Consequently, several α subunits expressed at even relatively low levels may compensate for inhibition of a more highly expressed subunit in hESCs and iPSCs, rescuing adhesion to Matrigel through redundant laminin- and collagen-binding integrins.
While the αVβ5 integrin heterodimer is necessary for hESC and iPSC initial adhesion to vitronectin, both β1 and αVβ5 appear to function in continued cell proliferation on vitronectin. In hESCs, the αVβ5 integrin heterodimer has been shown to be important for initial cell attachment and adhesion to vitronectin [7], though its role in hESC proliferation was not reported. Previous studies using human carcinoma and melanoma cell lines have shown that while both the αVβ5 and αVβ3 integrin heterodimers are central to initial cell attachment and adhesion on vitronectin, only the αVβ3 heterodimer may be necessary for cell spreading and migration, although the relative importance of these 2 vitronectin-binding heterodimers in adhesion and proliferation can vary between cell lines [25,26]. It is likewise possible that in hESCs and iPSCs, αVβ5 is important for initial cell adhesion, but that other vitronectin-binding factors, such as the αVβ3 or αVβ1 integrin heterodimers, may then be involved in continued cell proliferation. Studies have reported very low or a complete absence of mRNA and protein expression of the β3 integrin subunit in hESCs [12,15] and some iPSCs (Supplementary Fig. 2). Other vitronectin-binding factors may also be involved in maintaining proliferation, such as the urokinase plasminogen activator receptor, which has been shown to be sufficient to change cell morphology and migration in HEK293 cells when interacting with vitronectin [27,28], and plasminogen activator inhibitor-1, which may have similar effects [27]. This may also help explain the delay observed in decreased proliferation when both β1 and αVβ5 inhibiting antibodies were added (Fig. 3E). Alternatively, the iPSCs may be secreting an adhesive factor that replaces the need for vitronectin interactions [7,12,14,15], or factors in the CM may contribute to the substratum [7]. If ECM proteins recognized by β1 integrins are secreted by iPSCs (or are pre sent in the CM), this could explain why blockade of both αVβ5 and β1 is required to inhibit proliferation.
Although we found that iPSCs and hESCs can be cultured on vitronectin and appear indistinguishable from the same cells maintained on Matrigel by morphology, pluri-potency, and karyotype, we repeatedly found slightly better performance of cells on Matrigel over vitronectin in our adhesion and proliferation assays. iPSCs cultured on MEFs prior to proliferation experiments displayed slightly better growth on Matrigel, suggesting that the Matrigel preference in these assays cannot be due to prior conditioning on Matrigel. However, because MEFs secrete a variety of ECM proteins, including fibronectin, laminin, and collagen [7], MEFs most likely provide an ECM environment more similar to Matrigel than to purified vitronectin [15]. Consequently, prior culturing of cells on MEFs may make the cells develop a preference for Matrigel over vitronectin. Since we found that iPSCs cultured on vitronectin for multiple passages have similar adhesion to both Matrigel and vitronectin, this further supports the idea that iPSCs can adapt to the surface they are cultured on, but do not necessarily significantly prefer one system over the other. As cells are transferred from feeders to Matrigel or vitronectin, it is also possible that a subpopulation of cells is selected, as some cell death is observed in both cases. Although overall iPSC adhesion and growth on Matrigel may be slightly improved compared to on vitronectin, it is remarkable that the 2 systems yield such similar assay results overall, since vitronectin is a single ECM protein while Matrigel is a rich mixture of ECM proteins and growth factors.
The findings reported here reveal the functional importance of integrins in established and novel feeder-free hESC and iPSC culture systems and may lead to development of improved protocols for stem cell growth. The long-term culture of iPSCs and WiCell hESCs on vitronectin reported here was done using mTESR1, a recently developed defined hESC media [4], and vitronectin purified from human plasma. Although it can be purchased commercially, vitronectin can also be cheaply purified in large quantities. However, recombinant vitronectin, which lacks any potential plasma contaminants, has been shown to be similar to plasma vitronectin in supporting long-term hESC culture [7]. Because the primary integrin-binding peptide in vitronectin is RGD [11], the results of this study support the use of RGD-based peptides in synthetic scaffold designs for undifferentiated hESC and iPSC culture [20 –22].
Variation in expression profiles and function of iPSC lines has been previously reported [1,8] as well as observed here. As the iPSC lines used in this study were derived through reprogramming with Oct-3/4, Sox-2, Nanog, and Lin-28 [1], additional studies should be performed to confirm consistency of integrin expression and function in other iPSC lines generated with alternative factors and methods. Recent progress in iPSC technology, such as the creation of iPSCs using non-integrating vectors [29] and recombinant, cell-penetrating proteins [30], may make the translation of iPSC to clinical applications more feasible.
Footnotes
Acknowledgments
We thank James A. Thomson and Jessica Antosiewicz-Bourget for the cell lines and zbFGF, Poornima Kolhar for help with the adhesion assay design, Amy Friedrich for assistance with cell culture, Qirui Hu for help with embryoid body differentiation design, Brian Matsumoto for technical support in the UCSB Integrated Microscopy Facility, and David Buchholz for critically reviewing the manuscript. The C1A9, MJ7/18, and BIIG2 monoclonal antibodies developed by Lori L. Wallrath, Eugene C. Butcher, and Caroline H. Damsky, respectively, and P4G11 and P5D2 monoclonal antibodies developed by Elizabeth A. Wayner were all obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by the University of Iowa, Department of Biological Sciences, Iowa City, IA 52242. This research was supported by grants from the Army Research Office, The California Institute for Regenerative Medicine, and Sigma Xi (Grant No. G200710111048337724).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.