
Abstract
Bone marrow-derived mesenchymal stem cells (MSCs) possess multilineage differentiation potential and can be used for the treatment of diabetic heart failure. However, hyperglycemia can affect the function of MSCs adversely and merits the requirement for a strategy to correct this anomaly. MSCs were isolated from the tibias and femurs of C57BL/6 wild-type mice at 60 days after induction of diabetes by streptozotocin. MSCs were characterized by flow cytometry for CD44 (97.7%), CD90 (95.4%), and CD105 (92.3%) markers and were preconditioned with insulin-like growth factor-1 (IGF-1) (50 ng/mL) and fibroblast growth factor-2 (FGF-2) (50 ng/mL) in combination for 1 h in serum-free Iscove's modified Dulbecco's medium. This was followed by hypoxic and high glucose insults to mimic diabetic heart microenvironment and to study the effect of preconditioning. Diabetic MSCs after treatment showed upregulation of IGF-1, FGF-2, Akt, GATA-4, and Nkx 2.5 and downregulation of p16INK4a , p66shc , p53, Bax, and Bak. Under hypoxic stress, preconditioned diabetic MSCs showed high superoxide dismutase activity (52.3%) compared with untreated cells (36.9%). This was concomitant with low numbers of annexin-V-positive cells, high in vitro tube-forming ability, and high chemotactic mobility to stromal cell-derived factor-1α after preconditioning in diabetic MSCs. Upregulation of Ang-I and VEGF and downregulation of p16INK4a were also observed in preconditioned cells under conditions of high glucose insult. Therefore, preconditioning with IGF-1 and FGF-2 in combination represents a novel strategy to augment MSC function affected by diabetes and holds significance for future strategies to treat diabetic heart failure.
Introduction
D
Development of oxidative stress compromises stem cell proliferation and mobilization and has been well documented [5,6]. In this respect, different strategies have been used to protect stem cell function from oxidative stress and promote proliferation and differentiation [7,8]. Preconditioning involves the use of growth factors, hypoxic shock, or anti-aging compounds [9,10] and represents an option for augmentation of stem cell potency [11]. Recent studies suggest that preconditioning of stem cells with growth factors recovers their cytoprotective effects, resulting in improved efficiency of the cell therapy [10]. Similarly, studies have also proclaimed antioxidant preconditioning to protect against oxidative stress by the activation of cellular antioxidant defenses [12].
Mesenchymal stem cells (MSCs) possess multilineage differentiation potential [13,14] and provide a good source for cell-based therapies [15,16]. Nevertheless, effect of diabetes on the function of MSCs has not been previously documented and is an area of great interest with implications for the repair of diabetic heart failure. In the present study, effect of diabetes on MSCs was investigated and found to enhance senescence and apoptosis and downregulate survival factors. Moreover, a preconditioning strategy based on insulin-like growth factor-1 (IGF-1) and fibroblast growth factor-2 (FGF-2) was employed to augment proliferation and tolerance to hypoxic and high glucose insults in diabetes-affected MSCs.
Materials and Methods
Animals
This investigation conforms with the Guide for the Care and Use of Laboratory Animals published by the U.S. National Institutes of Health (NIH Publication No. 85-23, revised 1985). All animals were treated according to procedures approved by the Institutional Review Board at the National Center of Excellence in Molecular Biology, Lahore, Pakistan.
Mouse model for diabetes
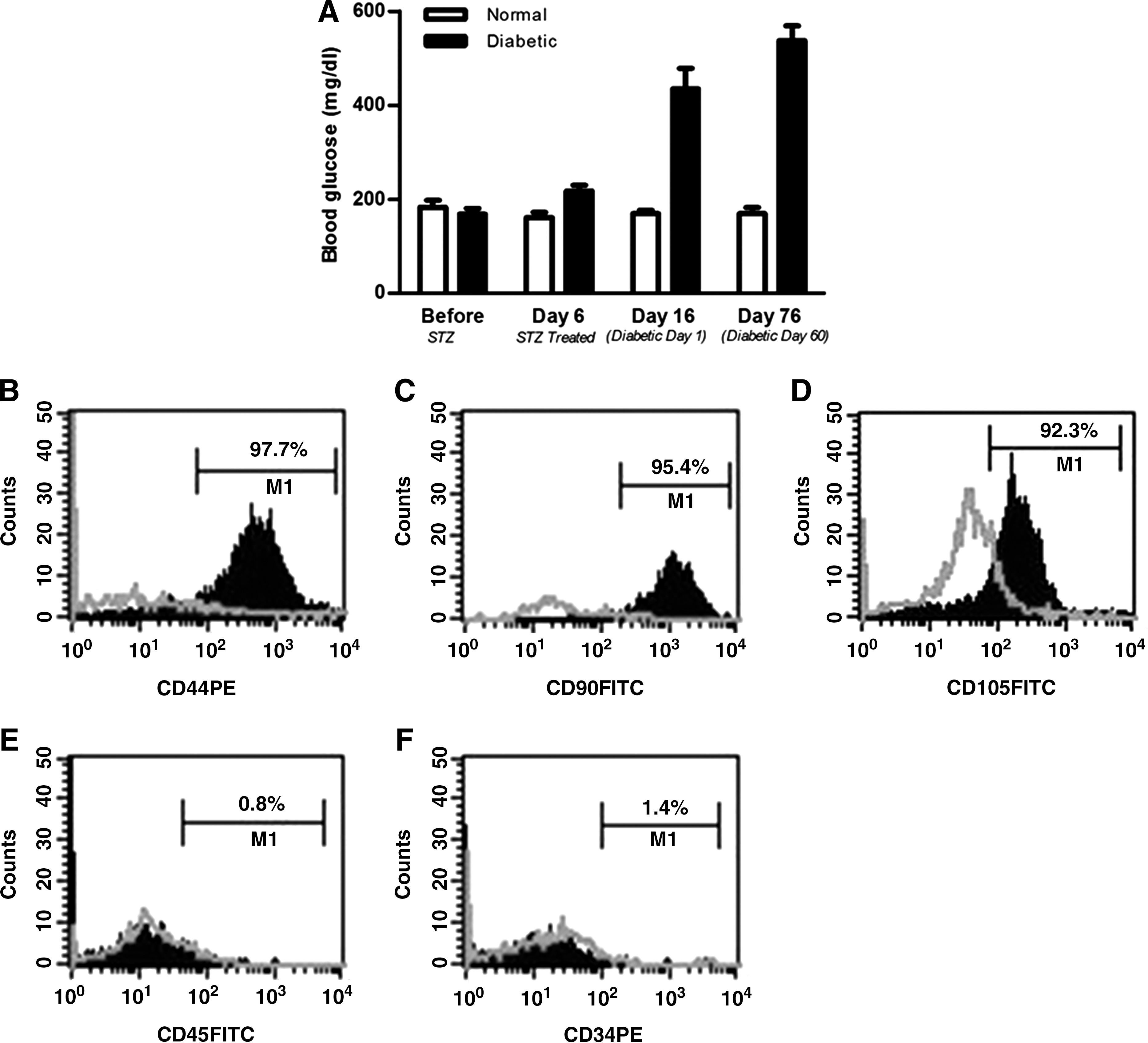
Diabetes was induced in C57BL/6 wild-type mice (6–8 weeks old) by intraperitoneal injection of streptozotocin (55 mg/kg) for 5 consecutive days, as described by Leiter and McNeill [17]. As streptozotocin metabolizes within 24 h, mice were housed with absorbent bedding and food and water ad libitum. At day 16 after the initial injection, mice were weighed and their blood glucose level was measured. Only streptozotocin-induced diabetic mice with blood glucose levels >300 mg/dL were included in the study.
Cell culture
MSCs were isolated as previously described [18]. Cells from tibia and femur of wild-type and 60-day diabetic C57BL/6 mice were cultured in Iscove's modified Dulbecco's medium (IMDM) supplemented with 20% fetal bovine serum (FBS), penicillin (100 U/mL), and streptomycin (100 μg/mL) at 37°C in humid air and 5% CO2. The medium was replaced every 3 days.
Flow cytometry
For flow cytometry analysis, MSCs from diabetic animals (N = 6) were washed with PBS (pH = 7.4) and incubated in the dark for 30 min at room temperature with CD45FITC, CD34PE, CD44PE, CD90FITC, and CD105FITC antibodies (all from BD Biosciences). The specific fluorescence of 10,000 cells was analyzed on FACScalibur (Becton Dickinson) using Cell Quest Pro software.
Preconditioning of diabetic MSCs
Diabetic MSCs were treated with IGF-1 (Santa Cruz Biotechnologies) and FGF-2 (Santa Cruz Biotechnologies) in different concentrations alone and in combination. To assess the optimal treatment regime, cell proliferation assay was performed with the use of XTT (sodium 3-[1-{phenylaminocarbonyl}-3,4-tetrazolium]-bis{4-methoxy-6-nitro} benzene sulfonic acid hydrate) according to the manufacturer's protocol (Roche). Briefly, 4 × 103 diabetic MSCs/well were plated in a 96-well plate containing IMDM with 20% FBS and left overnight in an incubator at 37°C and 5% CO2. Cells were exposed to different combinations (ng/mL) of IGF-1 and FGF-2 for 1 h under serum-free conditions and cell proliferation was measured after 48 h of growth factor treatment. Absorbance values were measured using Spectra max PLUS 384 (Molecular Devices) at 450 nm, with 650 nm as reference wavelength.
Parallel experiments showing the effect of other factors, such as vascular endothelial growth factor (VEGF) (Santa Cruz Biotechnologies), potentially involved in protection from hyperglycemia were done and cell proliferation was measured by real-time polymerase chain reaction (PCR) analysis for proliferating cell nuclear antigen (PCNA) on a 7500 Real-Time PCR system (Applied Biosystems).
Gene expression profiling of MSCs
RNA was extracted from normal, diabetic untreated, and diabetic MSCs preconditioned with either IGF-1/FGF-2 or VEGF using trizole reagent (Invitrogen Corporation) and quantified using ND-1000 spectrophotometer (NanoDrop Technologies). cDNA synthesis was carried out from 1 μg of RNA sample using Moloney Murine Leukemia Virus (M-MLV) reverse transcriptase (Invitrogen Corporation). Reverse transcriptase (RT-PCR) analysis for IGF-1, FGF-2, Akt, p16INK4a , p66shc , p53, Bax, Bak, GATA-4, and Nkx 2.5 was carried out using a GeneAmp PCR system 9700 (Applied Biosystem) with glyceraldehyde 3-phosphate dehydrogenase as internal control. Gels were quantified using Quantity One® 1-D Analysis Software, version 4.4 (Bio-Rad Laboratories, Inc.), according to the instructions given in user guide, with quantity of the ladder (Fermentas) used as standard. The sequences (5′–3′) for the primer pairs and their product lengths (bp) have been mentioned in Table 1.
Hypoxic stress
To analyze the effect of preconditioning 2 sets of diabetic MSCs, one without any treatment and another after preconditioning were subjected to hypoxic stress with H2O2 (100 μM) for 90 min under serum-free conditions. This was followed by 1 h recovery in serum-supplemented medium after which diabetic MSCs were analyzed.
Superoxide dismutase assay
Activity of superoxide dismutase (SOD) produced by both sets of MSCs was determined by the SOD activity colorimetric assay kit (Abcam) according to the manufacturer's protocol. Briefly, protein was isolated using lysis buffer from both sets of MSCs and SOD activity was measured using 10 μg of the total protein extract after 24 and 48 h of recovery. Absorbance values were measured using Spectra max PLUS 384 (Molecular Devices) at 450 nm.
Apoptosis
The number of apoptotic cells in both sets of MSCs was evaluated with fluorescein isothiocyanate Annexin-V kit according to the manufacturer's protocol (Abcam) and quantified by flow cytometric analysis on FACScalibur (Becton Dickinson).
In vitro tube-forming assay
Matrigel was thawed on ice to prevent premature polymerization. Aliquots of 50 μL were plated into individual wells of a 48-well tissue culture plate (Corning) and allowed to polymerize at 37°C for at least 30 min. To investigate the effect of preconditioning on the tube-forming ability of diabetic MSCs, cells were removed with trypsin/ethylenediaminetetraacetic acid and plated on Matrigel-coated wells in a concentration of 1.5 × 104 cells/well. The formation of tubular structures in the respective wells were examined at 6, 24, and 48 h after incubation of cells at 37°C in 5% CO2 with a phase-contrast microscope (IX-51 Olympus).
Chemotactic attraction
The effect of preconditioning on the ability of diabetic MSCs to mobilize in response to chemotactic signals was analyzed as described by Kucia et al. [19]. For this purpose, 5 × 104 cells/well were plated in a 6-well transmembrane culture plate (Costar). In the lower chamber, 2 mL of serum-free medium supplemented with 50 ng/mL stromal cell-derived factor-1α (SDF-1α; Upstate Biotechnology) was added. Both sets of MSCs after hypoxia were plated in the upper chambers. Wells without SDF-1α-supplemented medium were considered as control. The number of migrated cells was counted in 10 high-power fields after 24 h by staining with 4′,6-diamidino-2-phenylindole using an inverted microscope IX-51 (Olympus) equipped with a digital camera DP-71 (Olympus).
Glucose stress
Both sets of MSCs were exposed to glucose stress (30 mmol/L) under serum-free conditions to determine the effect of preconditioning on the ability of cells to tolerate hyperglycemia. The cells were allowed to recover for 1 h in serum-supplemented medium. RNA was isolated from the cells using trizole reagent (Invitrogen Corporation) for analysis of Ang-I, VEGF, and p16INK4a expression. Quantitative PCR (qPCR) was done on IQ5 Multi color Real-Time PCR Detection System (Bio-Rad) using Maxima SYBR Green qPCR Master Mix (Fermentas) in a reaction volume of 25 μL.
Statistical analysis
All data are expressed as mean ± standard error of the mean. Analysis of cell proliferation, densitometry, SOD activity, apoptosis, and chemotactic ability was performed using Student's unpaired t-test (P value of <0.05 was considered statistically significant).
Results
Characterization of MSCs
Experimental animals at 60 days after streptozotocin administration and with blood glucose >300 mg/dL (Fig. 1A) were used in the study. MSCs from these animals were cultured and analyzed by flow cytometry for the expression of stem cell markers. Cells were found to be positive for MSC markers CD44 (97.7%), CD90 (95.4%), and CD105 (92.3%) (Fig. 1B–D) and negative for hematopoietic markers CD45 (0.8%) and CD34 (1.4%) (Fig. 1E–F).
Stimulation of cell proliferation in preconditioned diabetic MSCs
Diabetic MSCs were preconditioned with different concentrations of IGF-1 and FGF-2. Cell proliferation was measured after 48 h of preconditioning to determine the best cytokine regimen. Highest cell proliferation was observed in diabetic MSCs treated with 50 ng/mL IGF-1 and 50 ng/mL FGF-2 in combination compared with untreated diabetic cells (1.61 ± 0.11 vs. 0.92 ± 0.08). Proliferative ability after treatment with 50 ng/mL IGF-1, 50 ng/mL FGF-2, 100 ng/mL IGF-1, 100 ng/FGF-2, and combination of 1 ng/mL FGF-2/50 ng/mL IGF-1 was 1.10 ± 0.11, 1.35 ± 0.12, 1.37 ± 0.05, 1.38 ± 0.09, and 1.44 ± 0.07, respectively. However, it was observed that 50/50 ng/mL IGF-1 and FGF-2 together showed highest cell proliferation (Fig. 2A) and therefore were used as the preconditioning strategy for subsequent experiments. Diabetic MSCs were also preconditioned with VEGF (50 ng/mL) and cell proliferation as PCNA expression was compared with IGF-1/FGF-2 treatment (Fig. 2B). The results demonstrated that IGF-1/FGF-2 treatment shows significantly higher PCNA expression than VEGF treatment.
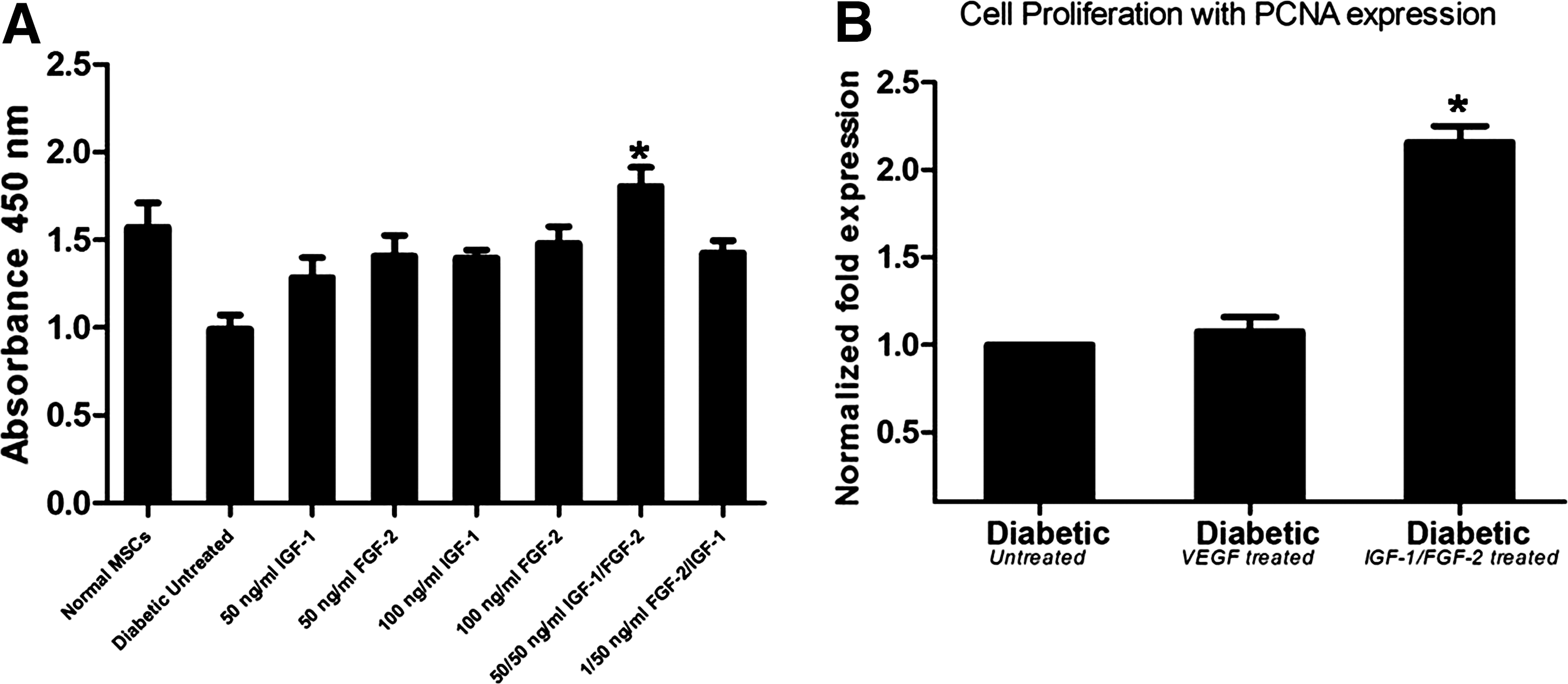
Cell proliferation after preconditioning of diabetic MSCs.
Gene expression profiling of MSCs
Gene expression profiling was done on normal, diabetic untreated, diabetic VEGF-treated, and diabetic IGF-1/FGF-2-treated MSCs. Figure 3A and B shows elevated levels of p16INK4a , p66shc , p53 (senescent markers), Bax, and Bak (apoptotic markers) and decreased levels of IGF-1, FGF-2, and Akt prosurvival factors in diabetic untreated MSCs compared with cells from normal animals. Diabetic MSCs after preconditioning with 50 ng/mL VEGF and 50/50 ng/mL IGF-1/FGF-2 showed improvement in gene expressions observed at various levels. Prosurvival proteins such as IGF-1, FGF-2, and Akt and differentiation markers GATA-4 and Nkx 2.5 showed low levels in diabetic untreated MSCs, whereas they were significantly increased after preconditioning. Diabetic MSCs showed high levels of Bax and Bak and their expression was significantly reduced after preconditioning. Similarly, markers of senescence such as p16INK4a , p66shc , and p53, which were elevated in diabetic untreated MSCs, showed significant reduction in expression after preconditioning with IGF-1 and FGF-2. However, there were significant differences in gene expression between VEGF-treated and IGF-1/FGF-2-treated MSCs (Fig. 3C, D). All quantification of gel bands was done by densitometry (Fig. 3E).

Gene expression profiling of mesenchymal stem cells.
Increased SOD activity in preconditioned MSCs
A significant increase in the activity was observed after 24 h of hypoxic treatment in preconditioned MSCs (52.3% ± 2.0%, P < 0.05) compared with untreated diabetic cells (36.9.8% ± 3.1%). SOD activity in both sets of MSCs at 48 h was measured to be 23.1% ± 2.2% and 32.2% ± 2.5%, respectively. SOD activity in MSCs from normal and diabetic animals subjected to normoxia was also recorded and used as a standard (Fig. 4A).
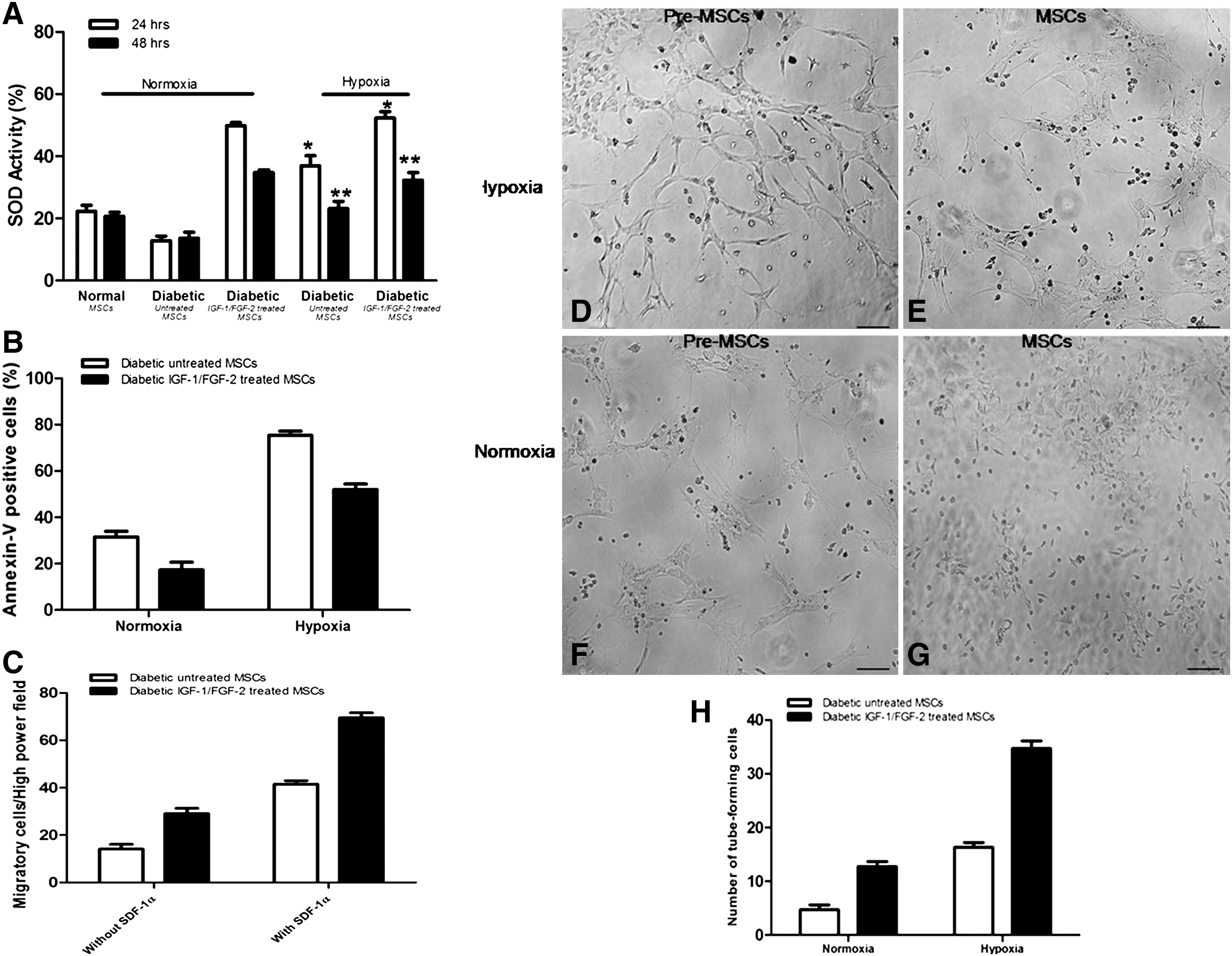
Diabetic MSCs under hypoxic stress.
Effect of IGF-1 and FGF-2 preconditioning on apoptosis
A significant decline in the number of apoptotic cells as determined by annexin-V was observed in preconditioned diabetic MSCs (51.8% ± 2.3%) after hypoxic treatment when compared with untreated diabetic MSCs (76.2% ± 1.8%) (Fig. 4B). Under normoxia, the number of apoptotic cells in both sets of MSCs was observed to be 31.5% ± 2.4% and 17.2% ± 3.3%, respectively.
Chemotactic ability of preconditioned diabetic MSCs
The ability of diabetic MSCs to respond to chemotactic signals from SDF-1α was analyzed. Preconditioned diabetic MSCs under hypoxic stress demonstrated significantly high number of cells (73.9 ± 5.3, P < 0.05) mobilized to SDF-1α when compared with untreated MSCs (31.9 ± 5.9) (Fig. 4C). Wells containing untreated and preconditioned MSCs but without SDF-1α treatment were considered as controls.
In vitro tube-forming ability in diabetic MSCs
In vitro formation of tubular structures under normoxic and hypoxic conditions were observed. Tube formation was found to be higher in preconditioned diabetic MSCs (Fig. 4D) under hypoxic conditions compared with untreated MSCs (Fig. 4E). Under normoxia, low tube formation activity was observed when compared with hypoxia in preconditioned and untreated diabetic MSCs (Fig. 4F, G). These results showed the enhanced angiogenic ability of preconditioned diabetic MSCs in response to hypoxic treatment in vitro compared with untreated diabetic MSCs. Quantification of tube formation confirmed the enhanced effect of IGF-1/FGF-2 treatment in diabetic MSCs compared with untreated cells (Fig. 4H).
Real-time PCR gene expression after glucose stress
Real-time PCR analysis for Ang-I and VEGF gene expression revealed an increase in preconditioned MSCs compared with untreated cells under high glucose conditions. Similarly, a decline in the p16INK4a expression was observed in preconditioned cells compared with untreated cells (Fig. 5).

Real-time reverse transcriptase (RT)–polymerase chain reaction gene expression under high-glucose conditions. FGF-2, fibroblast growth factor-2; IGF-1, insulin-like growth factor-1; VEGF, vascular endothelial growth factor.
Discussion
Bone marrow-derived MSCs represent one of the most widely used stem cell type for the repair of damaged myocardium [20,21]. Development of MSC-based therapies for the treatment of diabetic heart failure is an important area of research with recent studies highlighting the potential of MSCs for the repair of diabetic heart [4,22,23]. However, hyperglycemia can force stem cells into early aging, affecting their reparability [24] and severely undermining stem cell-based therapies for the treatment of diabetic heart failure. Therefore, the aim of the present study was to determine the effect of diabetes on bone marrow-derived MSCs and develop a preconditioning strategy to improve the cellular response affected by diabetes.
Our results demonstrated high expression of senescent (p16INK4a , p66shc , p53) and apoptotic (Bax, Bak) genes in diabetic MSCs compared with MSCs from normal animals. Markers for cell survival IGF-1, FGF-2, and Akt were also found to be downregulated in diabetic MSCs compared with normal MSCs. These findings highlighted the irregularities in MSCs as a result of diabetes and are in concordance with similar reports of stem cell dysfunction due to hyperglycemia [21,25]. It has been shown that diabetes can lead to elevated levels of p16INK4a , p66shc , and p53, promoting apoptosis in stem cells (24). As a correlation has been documented between elevated levels of senescent markers and a consequent decline in prosurvival factors IGF-1 and Akt, this mechanism may also seem to be involved in diabetic MSCs and may help explain our findings.
A preconditioning strategy to augment diabetes-affected impairment of MSC function was devised. IGF-1 and FGF-2 were used together in a concentration as determined by the highest cell proliferation activity. Diabetic MSCs after preconditioning demonstrated improvement witnessed at several fronts as shown in Fig. 3. There was not only significant upregulation in prosurvival genes IGF-1, FGF-2, and Akt but also downregulation of senescent (p16INK4a , p66shc , and p53) and apoptotic (Bax, Bak) genes after preconditioning in diabetic MSCs. One of the explanations for this amazing cellular response was based in our decision to employ IGF-1 and FGF-2 in combination. Recent data have implicated IGF-1 to be involved in survival pathways [26] and so IGF-1 has been widely used to enhance stem cell function [27 –29]. On the other hand, FGF-2 works by promoting cell proliferation [30] and suppressing cellular senescence [31]. Therefore, using these growth factors in combination we were able to enhance proliferation, improve the expression of cell survival genes, and more importantly, cause a decline in the expression of senescent markers.
To highlight the beneficial effect of IGF-1/FGF-2-based preconditioning, a parallel treatment option using VEGF was used. The rationale behind this approach was to compare IGF-1/FGF-2 preconditioning with a growth factor with potential effects on hyperglycemia. Our results demonstrated that VEGF treatment does help to enhance cell proliferation, augment the senescent state, and improve survival signaling in diabetic MSCs; however, this effect is less than the overall effect due to IGF-1/FGF-2 treatment. Therefore, IGF-1/FGF-2 preconditioning has a higher protective effect from hyperglycemia on MSCs and so it was employed for further analysis in subsequent experiments.
Preconditioned MSCs were subjected to hypoxic and high glucose insults in a way to mimic the diabetic heart microenvironment. The result showed low SOD activity in diabetes-affected MSCs that was significantly improved after preconditioning as evidenced in Fig. 4A. Recently, it has been shown that SOD activity is dependent on ROS levels and is affected by the change in donor age [32]. As diabetes promotes high ROS levels [7], this may explain the observed low SOD activity in diabetic MSCs. Stem cell susceptibility to apoptosis and necrosis due to ROS as a consequence of hyperglycemia is well documented [33] and is associated with altered cellular angiogenic response [34]. Figure 4B concurs with this finding as it demonstrates high number of apoptotic cells in diabetic MSCs compared with significantly reduced apoptotic cells after preconditioning and was concomitant with enhanced tube-forming ability in vitro in preconditioned MSCs compared with untreated cells.
It was also observed that the chemotactic attraction of diabetic MSCs in response to SDF-1 was significantly improved after preconditioning compared with untreated diabetic cells. Hyperglycemia impairs the SDF-1α–CXCR-4 pathway, affecting endothelial progenitor cell (EPC) recruitment in patients with coronary artery disease [35]. Among other factors, SDF-1α–CXCR-4 axis seems to be critical for recruitment and mobilization of stem cells [36] and it has been shown that different types of stem cells respond to ischemia-induced SDF-1α by upregulation of CXCR-4 receptor [37,38]. Our findings are in concordance with this idea and diabetic MSCs show low chemotactic response, which is enhanced by the treatment of these cells with IGF-1 and FGF-2.
To evaluate the tolerance of preconditioned diabetic MSCs to glucose level, we set up a simple culture system comprising of high glucose (30 mmol/L) treatment. This culture system was not only used to determine the effect of preconditioning on the ability of diabetic MSCs to tolerate high-glucose environment but also to emulate the clinical hyperglycemic condition in vitro. Further, these glucose levels may correspond to levels observed in patients with poorly controlled diabetes or under some stress conditions such as infection, inflammation, or hospitalization [39]. Our results also showed upregulation of Ang-I and VEGF coupled with downregulation of p16INK4a in preconditioned diabetic MSCs compared with untreated cells. This is of immense significance as it not only demonstrates better survival and angiogenic abilities of preconditioned cells under high-glucose conditions but also an ability to withstand cellular senescence.
Preconditioning with IGF-1 and FGF-2 can enhance cell proliferation, upregulate prosurvival, and downregulate senescent genes in the diabetes-affected MSCs. Further, this remarkable turnaround in diabetic MSCs after preconditioning resulted in an improved ability to tolerate hypoxic and high glucose insults. In future, these findings will be helpful for studies on diabetic impaired stem cells and can be employed for the treatment of diabetic heart failure.
Footnotes
Acknowledgments
The authors thank their colleagues for the review of this manuscript. This work was supported by research grants from the Ministry of Science and Technology (MoST), Government of Pakistan, Pakistan Academy of Sciences, and Higher Education Commission (HEC) of Pakistan.
Author Disclosure Statement
The authors declare no conflicts of interest.