
Abstract
Transplacental traffic of fetal progenitor and differentiated cells is a well-known phenomenon in pregnancies. We hypothesize that intrauterine stem cell transplantation leads to microchimerism in the dams and that this is gestational age-dependent. EGFP+ fetal liver-derived mesenchymal stem cell (MSC) (105 per fetus) were injected intraperitoneally into congeneic and allogeneic recipient fetuses at E12 versus E13.5 of murine pregnancy (56 dams). Engraftment in maternal organs was evaluated using TaqMan quantitative polymerase chain reaction (PCR) and fluorescence microscopy during pregnancy (1, 3, and 7 days after in utero transplantation [IUT]) and after delivery (1 and 4 weeks after delivery). One day after IUT donor cells were mainly found in the placenta (E12: 9/10 dams vs. E13.5: 4/8 dams) and laparotomy site (E12: 5/10 dams vs. E13.5: 4/8 dams). Three days after IUT these probabilities decreased significantly in the placenta to 3/8 and 1/3, respectively, whereas it was increased within the surgical wound to 8/8 and 2/4. One week after IUT donor cells could be detected in other single maternal organs, such as bone marrow or spleen. The surgical wound was chimeric in all dams. One week after delivery the surgical wound was still a major site of engraftment in both groups. E12 IUT resulted in detectable donor cell microchimerism in the maternal bone marrow (3/4), liver (2/4), lungs (1/4), spleen (1/4), and thymus (1/4), whereas engraftment probabilities were lower following E13.5 IUT (BM: 1/4, liver: 2/4, lungs: 1/4, spleen: 1/4, thymus: 0/4). At 4 weeks after delivery persistent microchimerism was found only after E12 IUT in various maternal organs (BM: 1/4, spleen: 1/4, lungs: 1/4) and within newly created surgical wounds (3/4), but completely not in the E13.5 group. Allogeneic IUT did also not result in any detectable long-term fetal microchimerism. An earlier IUT might lead to a higher transplacental traffic of donor MSC and persistent microchimerism within maternal tissues. Even 4 weeks after delivery, these cells are present in surgical wounds.
Introduction
T
A number of reports suggest that small numbers of fetal progenitor cells may also be present in maternal blood, and may persist for decades post-pregnancy [8]. However, due to low numbers in peripheral maternal blood their isolation and culture are technically challenging [9,10]. It seems that fetal cells are easier to detect in maternal sites of inflammation and tissue regeneration, such as systemic sclerosis, Hashimoto disease, or others [11,12]. Obviously, these fetal progenitor cells not only persist and hide long term, but also can have an impact on tissue remodeling and thus probably maternal health status [13 –15].
Currently, studies on fetomaternal stem cell traffic exclusively focus on the naturally occurring fetal microchimerism [16]. In the light of evolving efforts in early prenatal diagnosis and intrauterine treatment of certain genetic diseases, such as hemoglobinopathies or enzyme storage diseases, different strategies to improve success rates have been proposed using intrauterine transplantation of fetal stem cells [17,18]. These approaches include the use of different stem cell types, such as embryonic stem cells or mesenchymal stem cells (MSCs), but also in vitro- or in vivo-manipulated stem cells [19 –21]. However, based on the above mentioned fact that there is no complete placental barrier between mother and fetus, it must be suspected that these intrauterine-transplanted fetal stem cells cross into the maternal circulation and also might persist within maternal niches for decades. In regard to the timing of in utero stem cell transplantation, it is unclear whether the optimal time point referring to the immature fetal immune system would also be ideal in terms of safety of the recipient's mother. Since it has been shown recently that gestational age has an impact on the probability of fetal microchimerism in normal murine pregnancies after syngeneic crosses, we speculated that earlier in utero transplantation (IUT) might be associated with less transplacental traffic [22,23]. Since the localization of fetal cells within the placenta changes tremendously between E12 and E13.5 (current standard time point for IUT in the mouse model) and additionally MSC niches change exactly during this time in ontogeny, we have compared probabilities, levels, and tissue distribution of microchimerism after congeneic in utero stem cell transplantation at E12 versus E13.5 [24,25]. We have also tested allogeneic MSC traffic after IUT since the immunomodulatory effects of MSC and their capacity to escape lysis by cytotoxic T cells and NK cells are currently discussed in the literature [26].
Materials and Methods
Recipient mice
Twelve-day or 13.5-day gestation (term = 20–21 days) C57/BL6 (H-2Kb) and Balb/c (H-2Kd) mouse fetuses served as recipients (RCC Ltd. Füllinsdorf, Switzerland). Virgin 8–10-week-old mice were time-mated overnight and separated the next morning. The day of the vaginal plug was considered as E0 of gestation. The experimental protocols were approved by the Local Animal Care and Use Committee Basel (Nr. 2166).
Harvest and preparation of fetal liver-derived MSC
Time-mated heterozygous C57/BL6 TgN(ACTbEGFP)1Osb pregnant mice (H-2Kb, eGFP+; SLC, Hamamatsu, Japan) at E14 were sacrificed. Immediately, fetuses were removed from their uteri and decapitation was performed. The very well-visible fetal livers were dissected and kept in sterile culture medium containing RPMI, 9% fetal calf serum (FCS), 9% HS, 9% glutamax 100×, 100 U/mL penicillin, 100 U/mL streptomycin, 10−4 β-2ME. The livers were then separately cut into small pieces and minced between two 30-μm nylon meshes. Cells were suspended in culture medium, spun down, resuspended, and seeded into separate culture dishes. MSCs were allowed to adhere to plastic for 24 h and grow nearly to confluency. Then cells were harvested using 0.05% trypsin–ethylenediaminetetraacetic acid (EDTA), neutralized with FCS, washed and resuspended in freezing medium containing 90% FCS and 10% dimethylsulfoxide (DMSO), and kept frozen in liquid nitrogen until further processing.
Characterization of fetal liver-derived MSC by flow cytometry
MSC were stained with a panel of antibodies (IgG2a and IgG2b) according to the manufacturer's instructions. Rat anti-mouse CD14, -CD31, -CD34, -CD44, -CD45, -CD105, -Sca-1, -MHC class I, and -MHC class II were purchased by eBiosciences (THP Medical Products VertriebsGmbH, Vienna, Austria). Rat anti-mouse CD29 was obtained from Research Diagnostics Inc. (Flanders, NJ); rat anti-mouse CD73 was obtained from BD AG (Allschwil, Switzerland). The following reagents were used as secondary antibodies: goat anti-rat IgG2a-FITC, goat anti-rat IgG2b-FITC, and goat anti-rat Ig-biotinylated (Serotec, Düsseldorf, Germany), Streptavidin-PerCP (BD AG Allschwil, Switzerland) and mouse anti-rat IgG2a-PE and -IgG2b-PE, respectively (Southern Biotech, BioConcept, Allschwil, Switzerland). The cells expressed CD29, CD44, CD73 (low), CD105, Sca-1, and were negative for CD14, CD31, CD34, CD45, MHC class I and class II.
Characterization of fetal liver-derived MSC by in vitro differentiation
The in vitro differentiation potential of the MSC into osteocytes, chondrocytes, and adipocytes was evaluated (see Fig. 1). Whereas differentiation into adipocytes occurred quite spontaneously, differentiation into osteocytes succeeded only after application of BMP-2 and retinoid acid [27]. Although bone marrow-derived MSC showed chondrogenic differentiation as a positive control, our fetal liver-derived MSC did not.
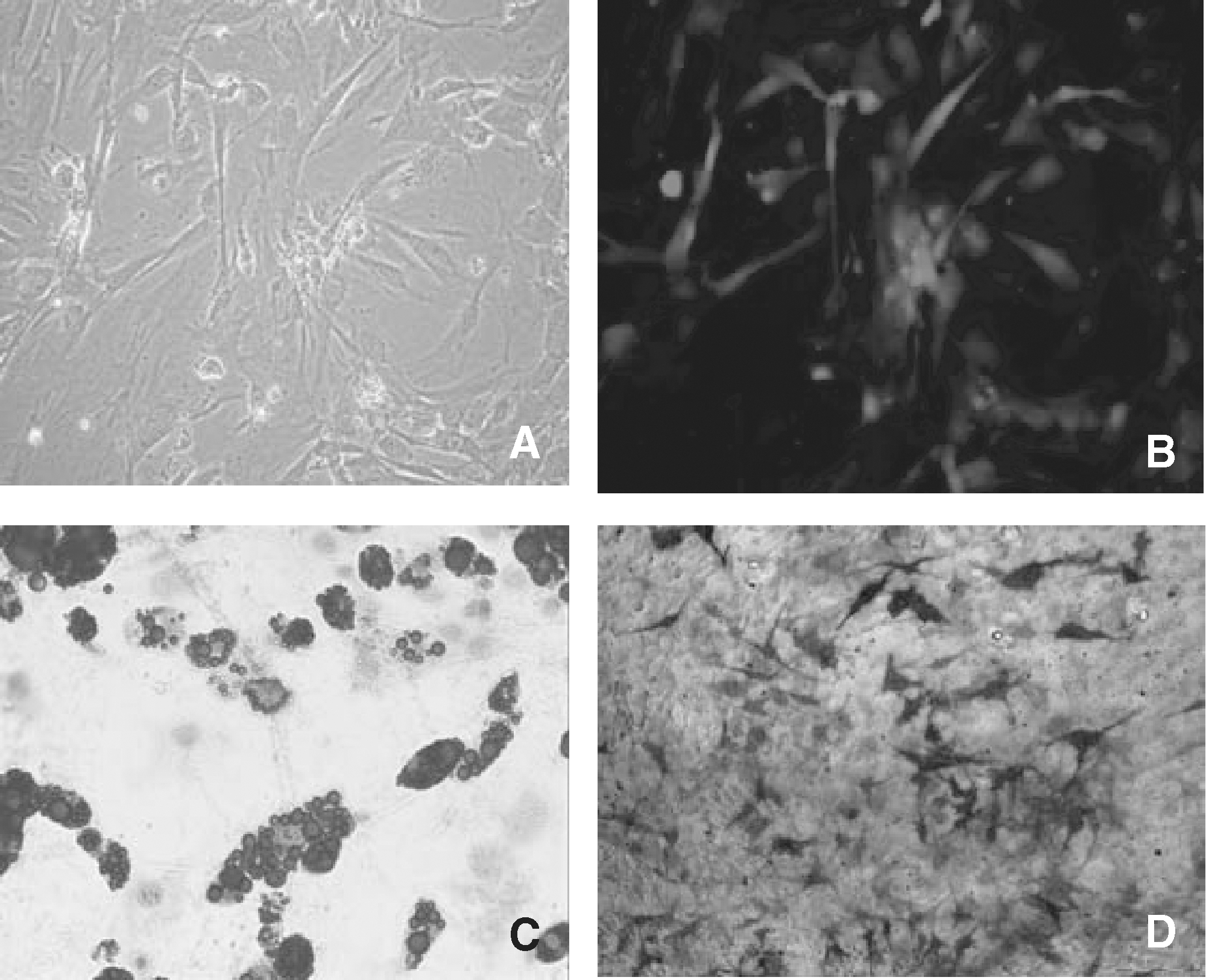
Donor mesenchymal stem cell (MSC) in (
In brief, for adipogenic differentiation expanded MSC were cultured in adipocyte-induction medium (DMEM containing FCS, Na-pyruvate, HEPES, PS, 10 μg/mL methyl-isobutylxanthine, dexamethasone 10−6 M, 10 μg/mL insulin, and 0.1 mM indomethacin for 3 days, following the culture in adipocyte-maintaining medium for 1 day (containing DMEM, FCS, Na-pyruvate, HEPES, PS as above and 10 μg/mL insulin). These treatment steps were performed 3 times. To visualize the accumulation of lipid-rich vacuoles, Oil Red O staining was used (Sigma-Aldrich, Buchs, Switzerland).
To promote osteogenic differentiation expanded MSC were cultured in α-MEM containing FCS, Na-pyruvate, HEPES, PS in the presence of ascorbic acid 0.1 mM, β-glycerophosphate 10 mM, dexamethasone 10−7 M, 50 ng/mL BMP-2, and 2.5 μM all-trans-retinoic acid. As a marker for osteogenic differentiation, alkaline phosphatase staining was used (Sigma-Aldrich, Buchs, Switzerland).
In utero transplantation
The surgical procedure was performed at either E12 or E13.5 as recently described [28]. Five microliters of PBS containing 105 MSC was injected per fetus. The exact needle position was proven by dye injection.
Recipient's sample preparation
Samples were taken at 1–7 days after IUT and at 1 resp. 4 weeks after delivery. Mice were sacrificed using carbon dioxide 85% v/v in air. Maternal organs (including placenta in cases of short-term analysis) were immediately removed and taken half for DNA extraction and half for fluorescence microscopy. DNA extraction was performed using the DNA extraction high pure preparation kit according to the manufacturer's instructions (Roche, Basel, Switzerland). The other half of the tissue was embedded into O.C.T. Compound Tissue-Tek® (Sakura, Sysmex Digitana AG, Horgen, Switzerland) and stored frozen at −70°C. Cryosection of 7 μm were used for immunohistochemistry.
Assessment of engraftment by quantitative PCR
A TaqMan assay was used to detect the EGFP gene that is expressed exclusively by the donor-derived MSC. Primers and probes were as follows: forward primer: 5′-GTCCGCCCTGAGCAAAGA-3′, reverse primer: 5′-TCCAGCAGGACCATGTGATC-3′, probe: 5′-CCCAACGAGAAGCG-NFQ-3′. Polymerase chain reaction (PCR) conditions were the following: 50°C for 2 min, 95° for 10 min, and 50 cycles at 95°C for 15 s and at 60°C for 1 min. As an indicator of the host cells, the common and well-characterized EGF promoter sequence was used (forward primer: 5′-TCCTTACTGGTCACCATCATCAATAC-3′, reverse primer: 5′-GTCCGTGAACTTCCCCTTCA-3′, probe: 5′-VIC-GTCCGTGAACTTCCCCTTCA-NFQ-3′). For quantification, the ABI PRISM® 7000 Sequence Detection System® software was used (Applied Biosystems Foster City CA). This assay is sensitive enough to detect 1 EGFP+ cell within 100,000 other cells. Samples were run in quadruplets, and considered positive if at least 2 of 4 wells had a detectable EGFP signal.
Assessment of engraftment by fluorescence microscopy
Cryosections of the recipient's tissue were fixed with acetone at room temperature, washed in TBS containing 0.05% Tween-20 and 0.2% BSA. Nuclei were counterstained using DAPI (Abbott, Baar, Switzerland). Results of direct fluorescence microscopy were confirmed using confocal microscopy with an emission peak wavelength of 509 nm.
Statistical analysis
The engraftment frequencies were compared between the E12 and E13.5 groups using chi-square test (SPSS for Windows 15.0, SPSS Inc., Chicago, IL). We did not compare the ct values of eGFP or the amount of eGFP genes between both groups since the actual amount was in most instances just above the detection limit.
Results
Placental microchimerism days after congeneic in utero transplantation (E13–E17; n = 30)
An in utero transplantation of EGFP+ fetal liver-derived MSC (see Fig. 1) at E12 resulted in placental microchimerism 1 day after transplantation in 90% of dams, whereas a later IUT at E13.5 had a lower rate of chimeric placentas being 50% (see Fig. 2). Three days after IUT the probability of detecting donor cells in the placenta significantly decreased in both groups being 37.5% in the E12 group (P = 0.019) and 30% in the E13.5 group (difference not significant). In all positive cases, we detected only single EGFP+ cells within the placenta and never cell clusters (see Fig. 3).
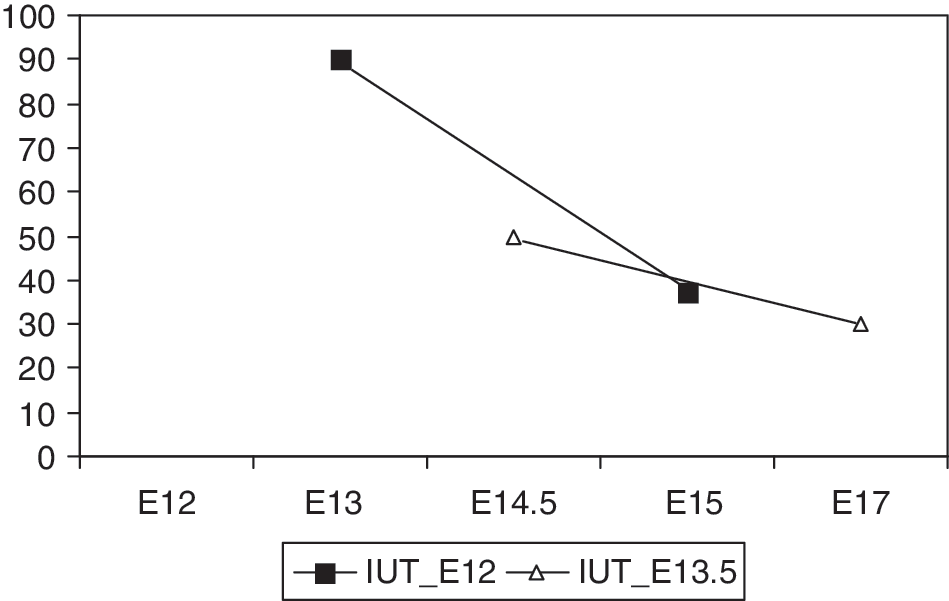
The probability of placental microchimerism is highest 1 day after E12 in utero transplantation (IUT) and decreases significantly 2 days thereafter (P = 0.019). Differences after E13.5 IUT are not significant.
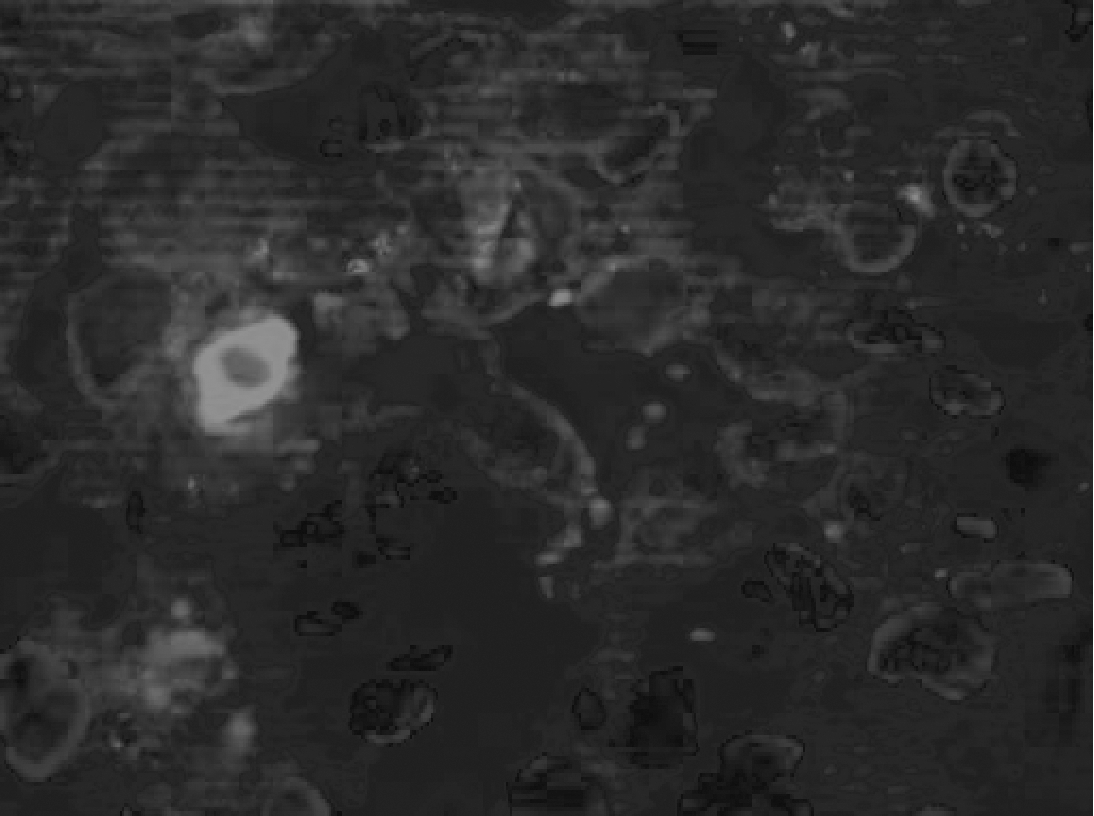
Single EGFP+ cell within the placenta 3 days after E12 IUT.
Microchimerism in maternal organs 1 week after congeneic in utero transplantation (E19–E20; n = 6)
One week after IUT, microchimerism in maternal organs was just at the detection limit of the PCR assay. We were, however, able to reliably detect EGFP+ single cells in maternal spleen (after IUT at E12) and bone marrow (after IUT at E13.5; data not shown).
Microchimerism in maternal organs 1 week after delivery following congeneic IUT (n = 8)
One week after delivery, all analyzed dams of the E12 and E13.5 groups showed microchimerism; however, organ distribution was significantly different (P = 0.047; see Table 1).After E12 in utero transplantation donor cells were found in maternal bone marrow in 3 of 4 cases, whereas this was true in only one of 4 mothers after E13.5 in utero transplantation (see Fig. 4).

Donor cell microchimerism after E12 IUT in maternal thymus (
Microchimerism in maternal organs 1 month after delivery following congeneic IUT (n = 7)
One month after delivery, donor cell microchimerism was significantly different between the E12 and E13.5 groups (P = 0.047) and could only be detected in 3 of 4 animals that had received in utero transplantation at E12. EGFP+ cells were found in the maternal bone marrow, spleen, and lungs of the recipients' mothers. We were unable to detect any donor cells in the E13.5 group (n = 3).
No persistent microchimerism in maternal organs following allogeneic IUT (n = 5)
When we analyzed Balb/c recipient mothers 1 month after delivery, we could only detect single donor cells in maternal thymus in 1 animal of the E12 group. There was no persistent microchimerism in the E13.5 group following allogeneic in utero transplantation (data not shown).
Donor MSC migrate to surgical wounds following congeneic IUT (n = 7)
Already 1 day after in utero transplantation at either E12 or E13.5 donor cells were found within the surgical wound of the midline laparotomy and were detectable at this site until 1 week after delivery (see Table 2).Four weeks after delivery, however, donor cells could not be detected at the laparotomy site. If a new surgical wound was placed by cutting a piece of the mouse tail, cells appeared again when the wound was analyzed 2 days later (see Fig. 5).
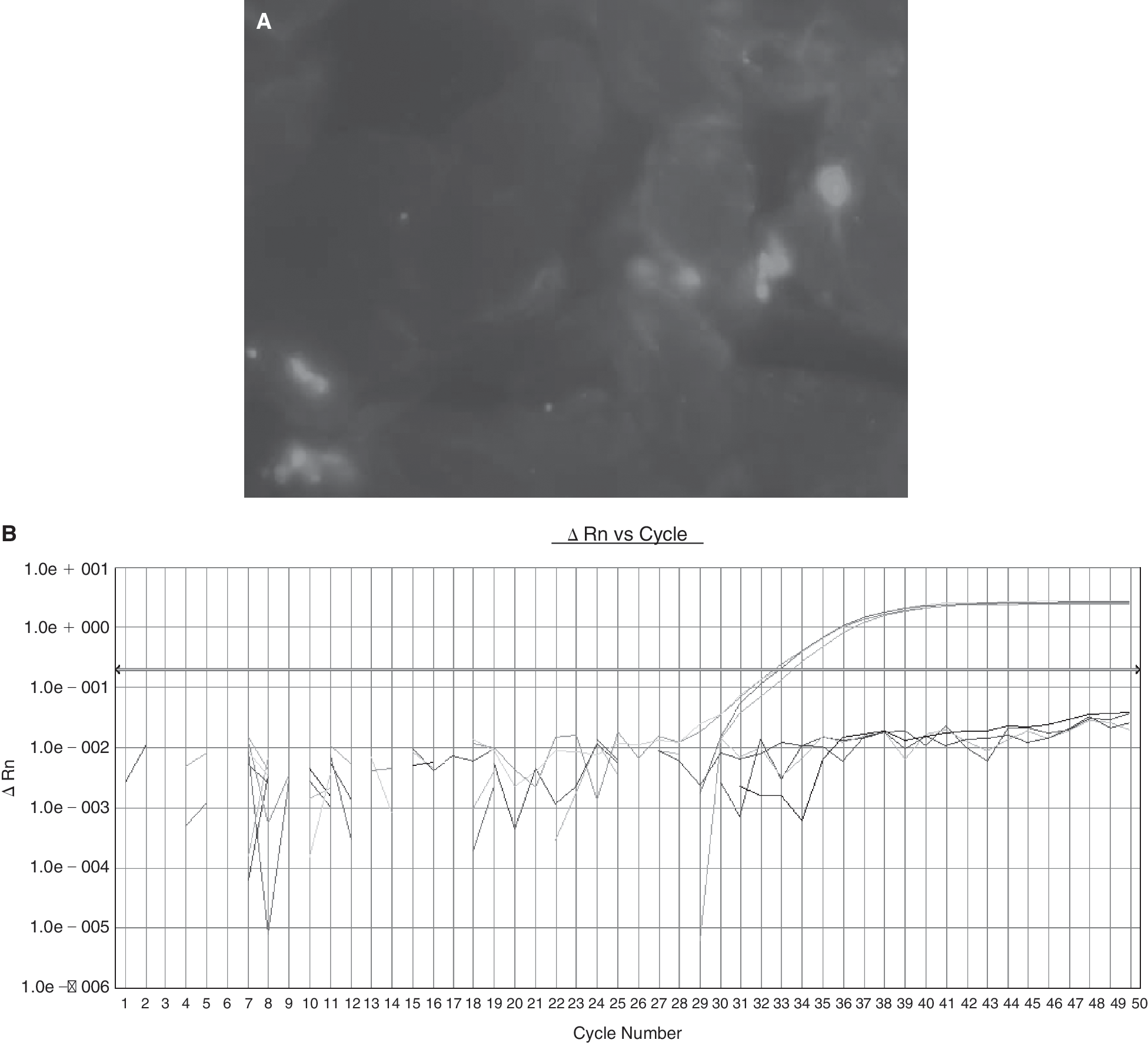
Donor EGFP+ cells at the laparotomy site 1 week after delivery detected by direct fluorescence microscopy (
Discussion
Intrauterine stem cell transplantation into immunocompetent recipients is still challenging, because engraftment levels are rather low and therapeutic effects are limited [29]. We aimed to perform an earlier transplantation (E12 vs. E13.5) to enhance engraftment levels in the recipient immunocompetent fetuses. For maternal safety purposes, however, we also wanted to examine the effect of an earlier IUT on the transplacental cell traffic and microchimerism in maternal organs.
Our study shows that intrauterine MSC transplantation at an earlier time point leads to a higher transplacental traffic of donor MSC and increased probabilities of long-term engraftment in various maternal organs, such as bone marrow, liver, lungs, and spleen. These cells might persist in maternal niches and migrate to surgical wounds even after a long-term interval. To our knowledge there are no studies on microchimerism in maternal organs following intrauterine stem cell transplantation yet.
Fetomaternal cell traffic during pregnancy and the persistence of microchimeric fetal cells for decades postpartum is accepted to be a common phenomenon in human and mammalian pregnancies [8,23]. It has also been shown that this cell traffic is increasing as pregnancy is progressing [30]. Increased fetal cell numbers are also described under certain conditions such as preeclampsia or after elective termination of pregnancy [5,7]. The effect of this persistent fetal microchimerism on maternal health status is a matter of debate. It is not clear whether the accumulation of fetal cells in certain maternal autoimmune diseases is a cause or bystander [12,31,32]. Animal experiments, however, suggest that microchimeric white blood cells of fetal origin might induce dermal fibrosis in parous, but not in virgin mice when mice are treated with vinyl chloride [33]. Although these microchimeric cells were not further characterized, the authors hypothesized that they might be lymphocytes that stem from transplacentally transferred fetal progenitor cells [34].
Based on these findings, it is important to gain more information on possible side effects of intrauterine stem cell transplantation on maternal health status. Since it has been shown lately that fetomaternal microchimerism is increasing with gestational age, we initially hypothesized that similarly an earlier time point of in utero stem cell transplantation would lead to lower levels of donor cells in the recipients' mothers [35,36]. Since it is unclear yet whether histocompatibility is an issue in this context, we tested both syngeneic and allogeneic constellations for the presence of donor cell microchimerism [37].
Using EGFP-expressing fetal liver-derived MSC as donor cells, our study shows that a relatively high proportion of recipients' mothers are microchimeric during and shortly after delivery. This observation is in agreement with earlier findings in both, human and murine pregnancies. Depending on the detection method and tissue used, the probabilities of microchimerism varied between 20% and 100% [23,36,37 –39]. Several studies suggest that cells with multilineage capacity migrate into the maternal circulation during pregnancy, persist long term, and might differentiate into various cell types [40 –42]. To our knowledge, however, this is the first study on donor cell microchimerism in the mothers after IUT.
In both groups (E12 vs. E13.5) donor cells are found in various maternal organs, such as thymus, spleen, liver, and lungs. However, the earlier intrauterine transplantation at E12 resulted in a higher proportion of chimeric maternal bone marrow. Our results are, hence, partly in agreement with the findings of Wang and colleagues who analyzed rat bone marrow after delivery and were able to detect small numbers of fetal EGFP+ cells within the maternal bone marrow in the absence of apparent injury [14]. Similarly, trafficking of fetal mesenchymal progenitors to the bone and bone marrow seems to be common in human pregnancies [41]. In contrast, in the last third of normal murine pregnancy and after delivery virtually no fetal cells could be detected in maternal bone marrow, whereas higher levels were found in maternal lungs, heart, spleen, and liver [23]. Studies show that the microchimeric fetal cells in normal murine pregnancies comprise a mixed population of progenitor and differentiated cells, with different relative proportions in different maternal organs [36]. Any invasive procedure during pregnancy, however, might lead to completely other results and has to be studied before procedures such as in utero stem cell transplantation could be staged as being low risk for the mother. In contrast to our transplantation experiments using pure EGFP+ donor MSC, analyses of “simple” mouse crossings suggest that the passage of fetal cells to the decidua begins with E10 and increases steadily. However, from their morphology it appears that these cells are rather of trophoblastic giant cell origin, than being fetal stem cells [24]. Our observation that E12 transplantation leads to a higher probability of microchimerism in maternal bone marrow could be explained by gestational age-dependent differences at the fetomaternal interface with regard to the changing expression pattern of homing-associated molecules between E12 and E14, which might be relevant not only for lymphocytes, but also for MSC [43]. Whether the SDF-1/CXCR4 axis would play a role in transplacental MSC traffic as it does for the migration of MSC to the bone marrow is unclear yet [44,45].
For long-term engraftment within the maternal environment, the gestational age-dependent dose of transplacentally transferred cells seems to be relevant. Four weeks after IUT, persistent donor cells are found only in the early (E12) transplantation group. This is true for syngeneic and to a lesser extent for allogeneic in utero transplantations. It seems that antigen dose is relevant for sustained peripheral T-cell tolerance [46]. The number of cases with allogeneic IUT in our series is low, but clearly shows that there is no persistent microchimerism after allogeneic IUT. In contrast, allogeneic crosses resulted in persistent fetal cells in maternal blood until 3 weeks after delivery [24]. Our results on syngeneic IUT and microchimerism at 4 weeks after delivery are also contrary to what is known on naturally occurring fetal microchimerism after syngeneic crosses. Although there are no differences between syngeneic and allogeneic crosses during pregnancy, syngeneic crosses had no detectable fetal cells at 2–3 weeks after delivery [23].
Transplanted progenitor cells might have an impact on tissue regeneration. In this study we have only tested surgical wounds in regard to accumulation of donor cells. During gestation and shortly after delivery, we were able to detect donor MSC at the laparotomy site. After this time period cells disappeared, since wound healing was completed. This observation is in line with data from human-injured tissues [47]. A lack in cytokines and extracellular matrix proteins might lead to a migration of the donor MSC back to their niche, for example, bone marrow or perivascular space [48,49]. In accordance with this model, we were not able to detect donor MSC in peripheral blood at any time. A new wound created by tail cut 4 weeks after delivery again recruited donor MSC to the surgical wound. However, this phenomenon could only be observed in the E12 group.
Our study has few weaknesses: we were unable to stimulate our MSC cell line to differentiate into chondrocytes. However, it seems to be a common issue that MSC of different mouse strains and origin show various differentiation potentials under similar culture conditions [50,51]. Though we have long experience in transuterine intra-abdominal puncture of mouse fetuses and also proved the correct needle position several times by dye injection, we cannot exclude that some donor cells leaked into the amniotic cavity after the procedure and did not cross into the maternal circulation through the placenta. This could only be further addressed if maternal organs are tested after other routes of administration.
With regard to maternal safety after intrauterine stem cell transplantation, we could predict from our animal model that an earlier IUT leads to a higher probability of persistent microchimerism in the recipient mothers. Since it appears that donor stem and immune cells are detectable at the sites of maternal surgical wounds, it needs to be clarified whether these cells enhance healing or lead to lesions comparable with GvHD or autoimmune inflammatory conditions. The results reported here should be taken into account before MSC are routinely used in ongoing human pregnancies, and this note of caution would even mere so apply, if embryonic or induced progenitor cells are used for any therapeutic intervention in the prenatal period. An earlier intrauterine transplantation of MSC leads to higher probabilities of donor cell microchimerism in the recipient dams.
Footnotes
Acknowledgments
This study was in part supported by the Swiss National Science Foundation NFP46 grant no. 4046-058662/1 and the Foundation “Basel Cord Blood Project—Therapy with stem cells from cord blood.” We thank Sinuhe Hahn for critically reviewing the manuscript.
Author Disclosure Statement
No competing financial interests exist.