
Abstract
Ethical and moral issues rule out the use of human induced pluripotent stem cells (iPSCs) in chimera studies that would determine the full extent of their reprogrammed state, instead relying on less rigorous assays such as teratoma formation and differentiated cell types. To date, only mouse iPSC lines are known to be truly pluripotent. However, initial mouse iPSC lines failed to form chimeric offspring, but did generate teratomas and differentiated embryoid bodies, and thus these specific iPSC lines were not completely reprogrammed or truly pluripotent. Therefore, there is a need to address whether the reprogramming factors and process used eventually to generate chimeric mice are universal and sufficient to generate reprogrammed iPSC that contribute to chimeric offspring in additional species. Here we show that porcine mesenchymal stem cells transduced with 6 human reprogramming factors (POU5F1, SOX2, NANOG, KLF4, LIN28, and C-MYC) injected into preimplantation-stage embryos contributed to multiple tissue types spanning all 3 germ layers in 8 of 10 fetuses. The chimerism rate was high, 85.3% or 29 of 34 live offspring were chimeras based on skin and tail biopsies harvested from 2- to 5-day-old pigs. The creation of pluripotent porcine iPSCs capable of generating chimeric offspring introduces numerous opportunities to study the facets significantly affecting cell therapies, genetic engineering, and other aspects of stem cell and developmental biology.
Introduction
T
However, only mouse iPSCs (miPSCs) have passed the stringent pluripotency test of contributing to tissues in the developing fetus and offspring after injection into preimplantation embryos [12]. The fact that teratomas could be generated from miPSC without using NANOG, but not chimeric offspring, raises the question whether these same or additional reprogramming factors are required in other species to generate chimeric offspring [10]. Besides answering this specific question, the pig is a logical choice to develop iPSC cells as the development of porcine iPSCs (piPSCs) offers significant opportunities to perform complex genetic modifications (multiple gene knockouts or knockins) for biomedical and agricultural purposes to study stem-cell-based therapies in large animal models, and potentially eliminates the need for animal cloning for agricultural purposes. Similarities between human and porcine anatomy and physiology have been frequently cited indicating them to be a more biologically relevant system than other models [15 –18], thus potentially leading to studies of stem cell therapies or creation of disease-specific mutants with significant human implications. Here we describe the production of porcine iPSC that efficiently produce chimeric offspring and contribute to the development of organs representing all 3 germ layers, trophectoderm, and potentially the germline. Now examining the contributions of various combinations of integrating and nonintegrating factors to determine their reprogramming effects and tumorigenicity is possible in a porcine model.
Materials and Methods
Cell culture and lentiviral transduction
Porcine mesenchymal stem cells (pMSCs) previously [19] derived in our lab from bone marrow were maintained in the MSC medium consisting of MEM Alpha (Gibco), 10% fetal bovine serum (HyClone), 2 mM
Alkaline phosphatase staining, immunocytochemistry, and flow cytometry
Alkaline phophatase (AP) staining was performed according to manufacturer's (Vector Laboratories) instructions. Immunocytochemistry was conducted by passaging piPSCs onto glass 4-chamber slides (BD Falcon) and fixed with 4% paraformaldehyde for 15 min. Antibodies were directed against POU5F1 (1:500; Santa Cruz), SOX2 (1:100; R&D Systems), TRA 1-81 (1:500, Millipore), and SSEA4 (1:20; Hybridoma Bank). Primary antibodies were detected using secondary antibodies conjugated to Alexa Fluor 488 or 594 (1:1000; Molecular Probes). Cell observations were made using the Ix81 with Disc-Spinning Unit (Olympus) and Slide Book Software (Intelligent Imaging Innovations).
Cells were fixed for flow cytometry with 4% paraformaldehyde for extracellular marker and 57% ethanol for intracellular marker staining for 10 min. Cells were blocked in 4% horse serum for 45 min. Primary antibodies were directed against POU5F1 (1:250), SOX2 (1:100), TRA 1-81 (1:500), and SSEA4 (1:20). Primary antibodies were detected using fluorescently conjugated secondary antibody Alexa Fluor 405 (1:1000; Molecular Probes). Cells were analyzed using a Dakocytomation Cyan (Beckman Coulter) and FlowJo Cytometry analysis software (Tree Star).
Population doubling time and embryoid body differentiation assays
Population doubling time was determined by plating piPSCs, BGO1 hESCs, and pMSCs at a concentration of 250,000 cells per 35-mm Matrigel–coated dish in the mTeSR1 medium with subsequent cell quantification every 12 h for 48 h. A nonlinear regression (curve fit) was performed using GraphPad Prism 5 (GraphPad Software) to establish population doubling time. Two-way ANOVA and Tukey's Pair-Wise comparisons were done utilizing SAS 9.1 statistical software (SAS Institute) contrasting doubling times between each population (N = 3), with P-values <0.05 considered significantly different.
Embryoid bodies (EBs) were formed by plating 1.2 × 106 cells in the mTeSR1 medium and Y-27632 ROCK inhibitor (10 μM; Calbiochem) into a single well of an AggreWell plate (Stemcell Technologies) and centrifuged at 100 g for 3 min per manufacturer's instruction. After 24 h, EBs were maintained in Petri-dish (BD Falcon) suspension culture in 20% KSR medium without fibroblast growth factor-2 and allowed to differentiate for 10 days. Each experiment was replicated a minimum of 4 times.
Polymerase chain reaction
For reverse transcription polymerase chain reaction (RT-PCR) analysis, total RNA was isolated from iPSCs using the Qiashredder and RNeasy mini kits (Qiagen) according to the manufacturer's protocol. The RNA quality and quantity was determined using the NanoDrop 8000 (Thermo Scientific). Total RNA (500 ng) was reverse-transcribed using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems). Reactions were initially incubated at 25°C for 10 min and subsequently at 37°C for 120 min. PCR amplification was performed using GoTaq Green (Promega). Primers used for RT-PCR were as follows: hPOU5F1 (for exogenous gene), forward 5′-GAGAAGGAGAAGCTGGAGCA-3′ and reverse 5′-TCGGACCACATCCTTCTCG-3′; porcine POU5F1 (pPOU5F1) (for endogenous gene), forward 5′-ACAAGGAGAAGCTGGAGCCG-3′ and reverse 5′-CGCGGACCACATCCTTCTCT-3′; α-fetoprotein (Endoderm), forward 5′-TGCTGCAAGTTGCCCACCACA-3′ and reverse 5′-TCCGCTTGGCCAGTGCTTGG-3′; brachyury (Mesoderm), forward 5′-GCAAAAGCTTTCCTTGATGC-3′ and reverse 5′-GCAGGATACCTCTCACAGCC-3′; neurogenic differentiation (Ectoderm), forward 5′-GACTTGCGTTCAGGCAAAAGC-3′ and reverse 5′-GGGCGACTGGTAAGAGTAGG-3′; GAPDH (internal housekeeping gene), forward 5′-CTCAACGACCACTTCGTCAA-3′ and reverse 5′-TCTGGGATGGAAACTGGAAG-3′. PCR reactions were performed by initially denaturing cDNA at 95°C for 3 min followed by 35 cycles of denaturing at 95°C for 60 sec, annealing at 60°C (64°C for pPOU5F1) for 30 sec, and polymerization at 72°C for 30 sec, and a final 10-min extension. PCR products were loaded into 2% agarose gels containing 0.6 μg/mL ethidium bromide and run in Tris-acetate-ethylenediaminetetraacetic acid buffer for 45 min. The U:Genius (Syngene) gel documentation station was used to observe PCR products.
Collection and micromanipulation of porcine embryos
Donor females were scheduled for embryo recovery using Altrenogest (Matrix), 1250 IU pregnant mare serum gonadotropin (Sigma-Aldrich), and 750 IU hCG (Chorulon). Donors were mated once 34 h after Chorulon injection. Embryo recipients were synchronized with Matrix, P.G. 600® (200 IU hCG, 400 IU PMSG; Intervet), and Chorulon.
The uterus, oviducts, and ovaries of donor pigs were exposed through a caudal mid ventral laparotomy. A total of 80 mL of PorcPro E-Flush (Minitube of America) was used to flush the uterus on day 4.5 postinsemination. Embryos were searched and evaluated for correct morphology and developmental stage. Microinjection of piPSCs was performed in PorcPRO E-hold medium (Minitube of America) under oil at 39°C on a Zeiss Axiovert 200 microscope equipped with Narishige micromanipulators. Up to 15 passage, 7 piPSCs of homogenous size and appearance were slowly injected into each embryo. Injected embryos were then cultured in the PorcPRO E-Cleave medium (19982/3010; Minitube of America) until transfer into the recipient.
piPSC-injected pig embryos were surgically transferred by transoviductal uterine catheterization [20] into uteri of asynchronous recipient female pigs (−24 h) into the distal tip of a single uterine horn using a 3.5-Fr. TomCat™ Catheter (Sherwood Medical). The uterus was placed back into the peritoneal cavity, and the recipient animals were sutured and placed into postoperative recovery. During gestation, real-time ultrasonography examination was used to confirm and monitor pregnancy using an Aloka 900 Ultrasound Scanner (Aloka Co. Ltd.) with an attached 5.0-MHz trans-abdominal probe. Recipient husbandry was maintained as for normal gestating and farrowing sows.
Tissue isolation and DNA extraction
To isolate fetal pigs 57 days after transplant, the selected female was prepared for mid ventral laparotomy as described above. Briefly, a 2-cm incision was made near the head of each fetus. After rupture of placental membranes, the exposed fetus was removed, and the umbilical cord was severed and placed immediately on ice. Organs/tissues, including brain, skin, liver, pancreas, stomach, intestine, lung, heart, kidney, muscle, spleen, placenta, and gonad, were harvested from individual pigs and snap frozen in liquid nitrogen. Genomic DNA (gDNA) was isolated from these tissues and piPSC and pMSC controls using the DNeasy Blood and Tissue Kit (Qiagen). Genomically integrated hPOU5F1 in these samples was detected by PCR analysis of isolated gDNA done as above using the validated human-specific POU5F1 forward 5′-GAGAAGGAGAAGCTGGAGCA-3′ and reverse 5′-TCGGACCACATCCTTCTCG-3′ primers (same as previously used for RT-PCR).
To ascertain whether furrowed pigs were chimeric, ear and tail tissue samples were collected from 36 two- to five-day-old pigs, and ear tissue samples were collected from 3 adult nonchimeric control pigs with all samples being snap frozen in liquid nitrogen. gDNA was isolated using the DNeasy Blood and Tissue Kit, and detection of genomically integrated hPOU5F1 and human-specific NANOG (forward 5′-CGCGCTACTTCGAGCGGAGG-3′ and reverse 5′-TGGAAAGCGGGCAGTGAGCG-3′ primers) was once again tested by PCR analysis.
Results
Production of porcine iPSCs
Porcine bone-marrow-derived mesenchymal cells (pMSCs; Fig. 1A) previously isolated and purified [19] were tested for POU5F1 (Fig. 1B–D), SOX2, SSEA4, and TRA 1-81 stem cell marker expression at multiple passages and were found to be negative. These cells were then reprogrammed using lentiviral transduction with 6 human pluripotency factors, POU5F1, SOX2, NANOG, LIN28, KLF4, and C-MYC, under elongation factor 1α promoter (Supplementary Fig. S1, available online at
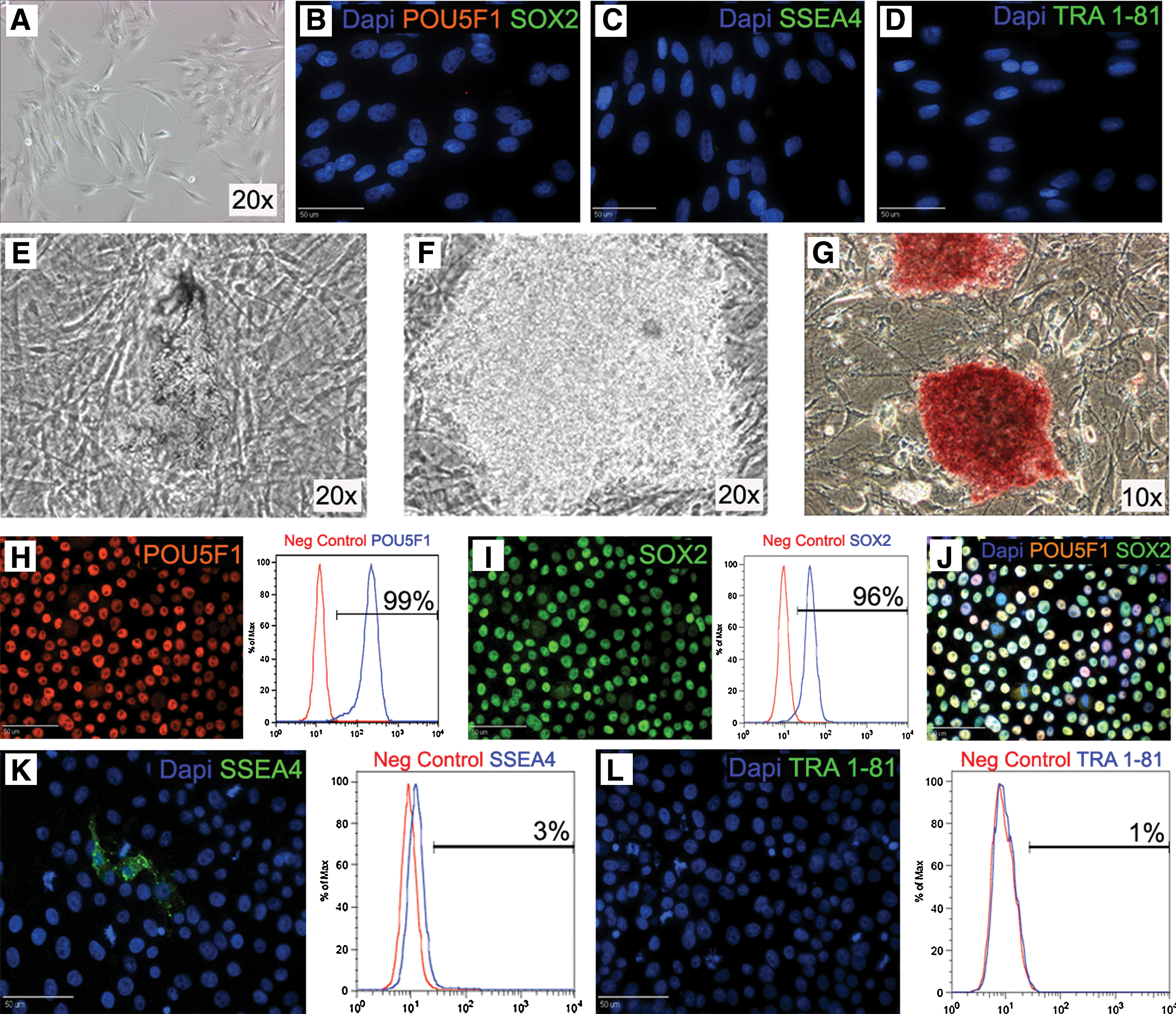
Formation of porcine induced pluripotent stem cells (piPSCs). Porcine mesenchymal stem cells (pMSCs) (
piPSCs express pluripotency markers and form all three germ layers during EB differentiation
After initial isolation, piPSCs were transferred from undefined feeder-based conditions to defined feeder-free conditions, mTeSR1 medium, and Matrigel-coated plates. Transfer to feeder-free conditions appeared to further purify piPSCs with high levels of initial cell death for the first 4–6 passages, resulting in increased homogeneity of cells with iPSC morphology. Immunostaining of piPSCs demonstrated protein expression of POU5F1 (Fig. 1H) and SOX2 (Fig. 1I; Merge J) with flow cytometry showing that >95% of cells were positive for both markers (Fig. 1H, I). However, piPSCs were mostly negative for pluripotency markers SSEA4 (Fig. 1K) and TRA 1-81 (Fig. 1L). Reprogramming of the pMSCs with exogenous factors also resulted in reactivation of endogenous porcine POU5F1 (Fig. 2A), which was not expressed in pMSCs or hiPSC controls. piPSCs maintained a normal karyotype for 20 passages (Fig. 2B) and have undergone >50 passages without loss of piPSC phenotype and immunoreactivity. piPSCs doubling time (22.3 h) was significantly (P < 0.05) lower than both hESCs (28.2 h) and pMSCs (32.2 h; Fig. 2C). These results indicate that piPSCs are a highly proliferative population that can be continually expanded without karyotypic abnormalities creating an excellent cell source for cell therapies and for genetic manipulation of livestock. piPSCs then underwent 10 days of EB (Fig. 2D) differentiation to ascertain their ability to differentiate into the 3 germ layers: ecto-, endo-, and mesoderm. Day 10 EBs expressed the ectoderm gene neurogenic differentiation (NeuroD; Fig. 2E), the endoderm gene α-fetoprotein (AFP), and mesoderm gene brachyury (T), whereas piPSCs were negative for these markers. Ten days of EB differentiation did not result in the termination of hPOU5F1 or pPOU5F1 gene expression (Fig. 2A). This is probably due to resistance to silencing caused by lentiviral integration [21,22].
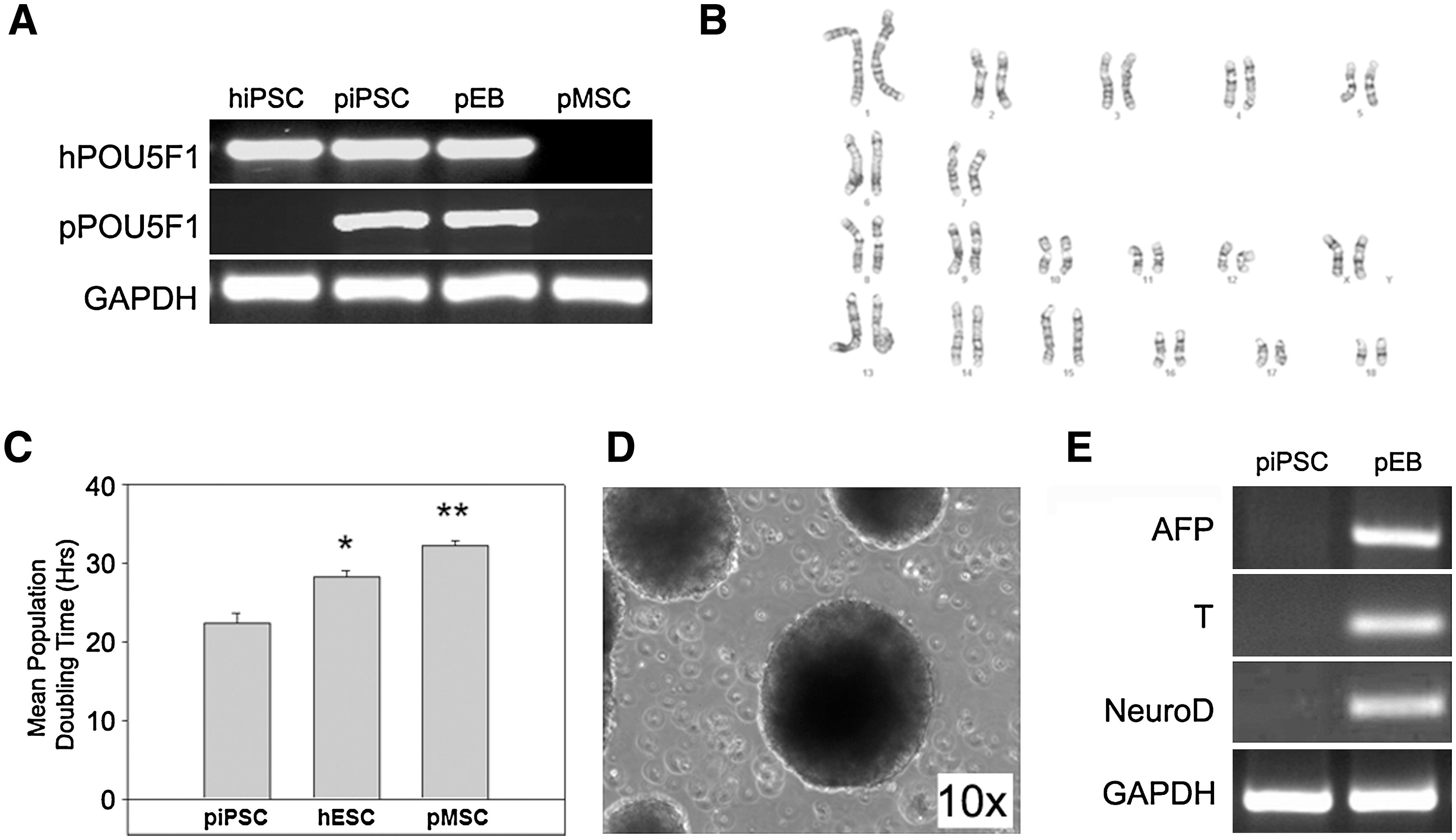
Expansion and embryoid body (EB) differentiation of piPSCs. Polymerase chain reaction (PCR) analysis using porcine-specific (p) and human-specific (h) POU5F1 primers indicated that reprogramming of pMSCs resulted in activation of endogens porcine POU5F1 (
piPSCs contribute to the formation of late-stage chimeric embryos and live offspring
In vitro differentiation of ESCs and iPSCs is a useful assay to determine the plasticity of a given cell type. However, the formation of chimeras and assessing the lineage contribution of iPSCs is a significantly more robust and definitive pluripotency assay. To this end, in vivo developing morula- and blastocyst-stage porcine embryos were collected from reproductive tracts at day 4.5 postinsemination. Then, 10–15 piPSCs were injected into each embryo (Fig. 3A, B; white arrows indicate piPSCs) and chimeric embryos were immediately transplanted into 6 synchronized recipients after injection. After 28 days, 5 recipients were determined to be pregnant via ultrasonography examination, and at day 57, 10 fetuses (9 developing and 1 deceased) were collected from a single recipient. PCR analysis of genomic-integrated hPOU5F1 was used to determine contribution of piPSCs in 13 tissues representing ectoderm (brain and skin), endoderm (liver, pancreas, stomach, intestine, and lung), mesoderm (heart, kidney, muscle, and spleen), trophectoderm (placenta), and potentially germline (gonad) of each fetus with the exception of fetus 8 (deceased fetus) where neither the spleen nor gonad could be isolated. Eight out of 10 fetuses possessed chimeric tissue (Fig. 3C), including all 3 germ layers, trophectoderm, and gonad, with 2 fetuses demonstrating piPSC contribution to all sampled tissue types (Fig. 3C: fetuses 1 and 9). All fetal pigs, with the exception of the single deceased individual (Fig. 3C: fetus 8), appeared to be developing normally and no overt differences were observed between chimeric (Fig. 3C: fetuses 1–4 and 7–10) and nonchimeric (Fig. 3C: fetuses 5 and 6) individuals. Expression of hPOU5F1 in 5 out of 10 fetal gonads suggests that piPSCs are potentially capable of contributing to the germline (Fig. 3C: fetuses 1, 3, 7, 9, and 10). However, further study is needed to confirm germline transmission. The formation of 8 late-stage chimeric fetal pigs and contribution to all 3 germ layers, trophectoderm, and gonad strongly indicate that piPSCs are pluripotent.
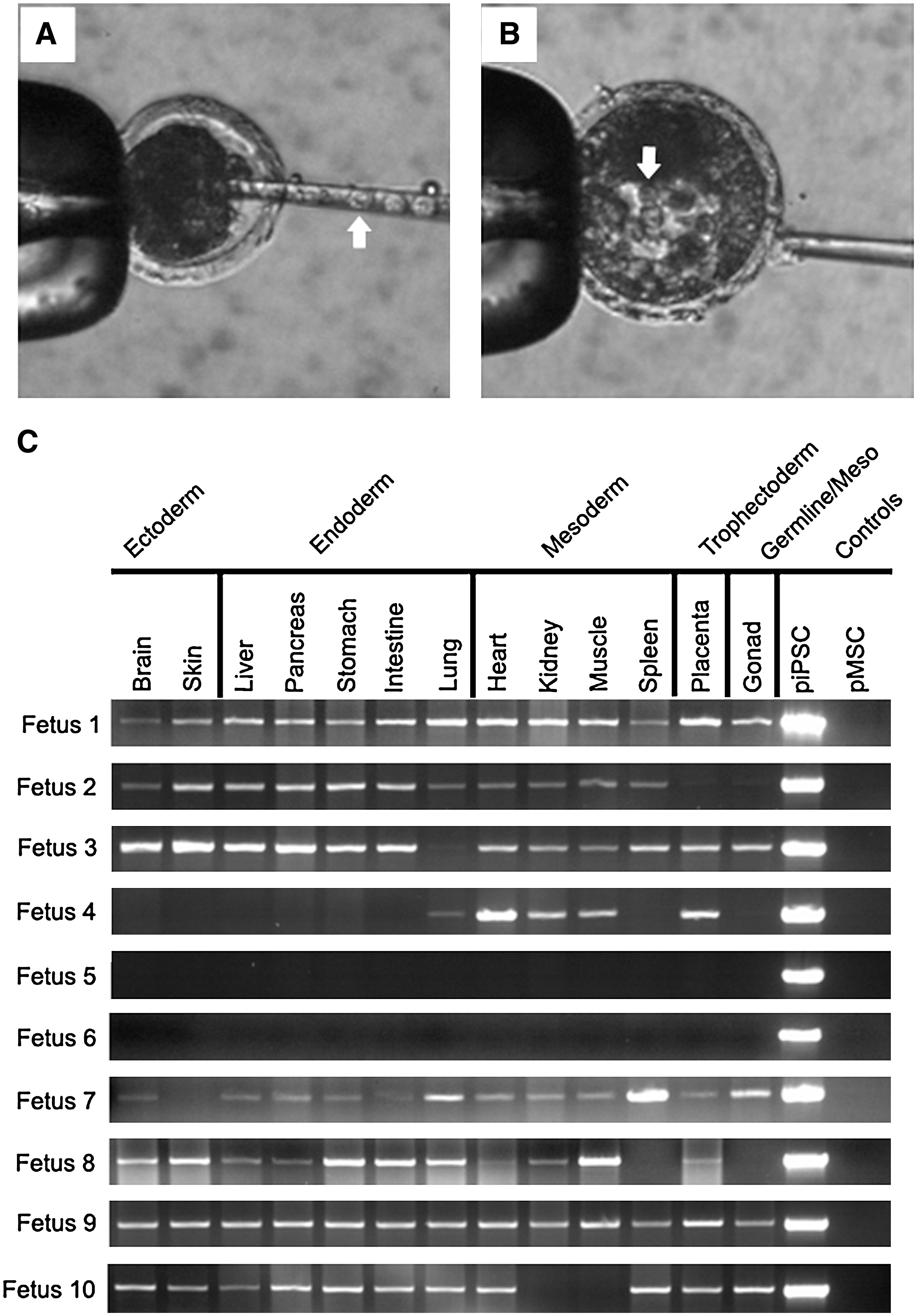
piPSCs produce fetal chimeras. Blastocyst- and morula-stage porcine embryos were freshly collected and injected with 10–15 piPSCs each (
Early publications describing iPSC formation in the mouse showed iPSC contribution to fetal development, but did not yield live chimeric offspring, suggesting incomplete reprogramming [10,14]. To ascertain whether piPSCs are fully reprogrammed, 4 recipient animals were allowed to carry chimeric embryos to term. After a normal gestation period, 34 live offspring and 2 stillborns were farrowed. Of the live births, only 2 appeared abnormal, 1 possessing a cleft pallet and the other being significantly smaller than average. Ear and tail biopsies were taken from all 36 pigs and were tested for the presence of human-specific POU5F1 and NANOG by PCR analysis. As hPOU5F1 and hNANOG were introduced independently of each other in separate vectors, the presence of both introduced genes in individual tissues strongly suggests piPSC contribution. The results demonstrated multiple hPOU5F1/hNANOG+ pigs (Fig. 4A) and that 85.3% or 29 of 34 live births were chimeric (Fig. 4B). The 27 chimeric pigs have shown no outward abnormalities and demonstrate typical growth and development (Fig. 4C) relative to their nonchimeric counterparts. Both stillborns (Fig. 4A: P35) and abnormal animals were also hPOU5F1/hNANOG+; however, the number of abnormal offspring was not unusually high, suggesting that this may not be a direct result of chimerism. PCR analysis was performed on 3 nonchimeric pigs, which were all found to be hPOU5F1/hNANOG− (Fig. 4A: NCP1-3). The live birth of 27 normal chimeric pigs further supports the full reprogramming of these piPSCs.
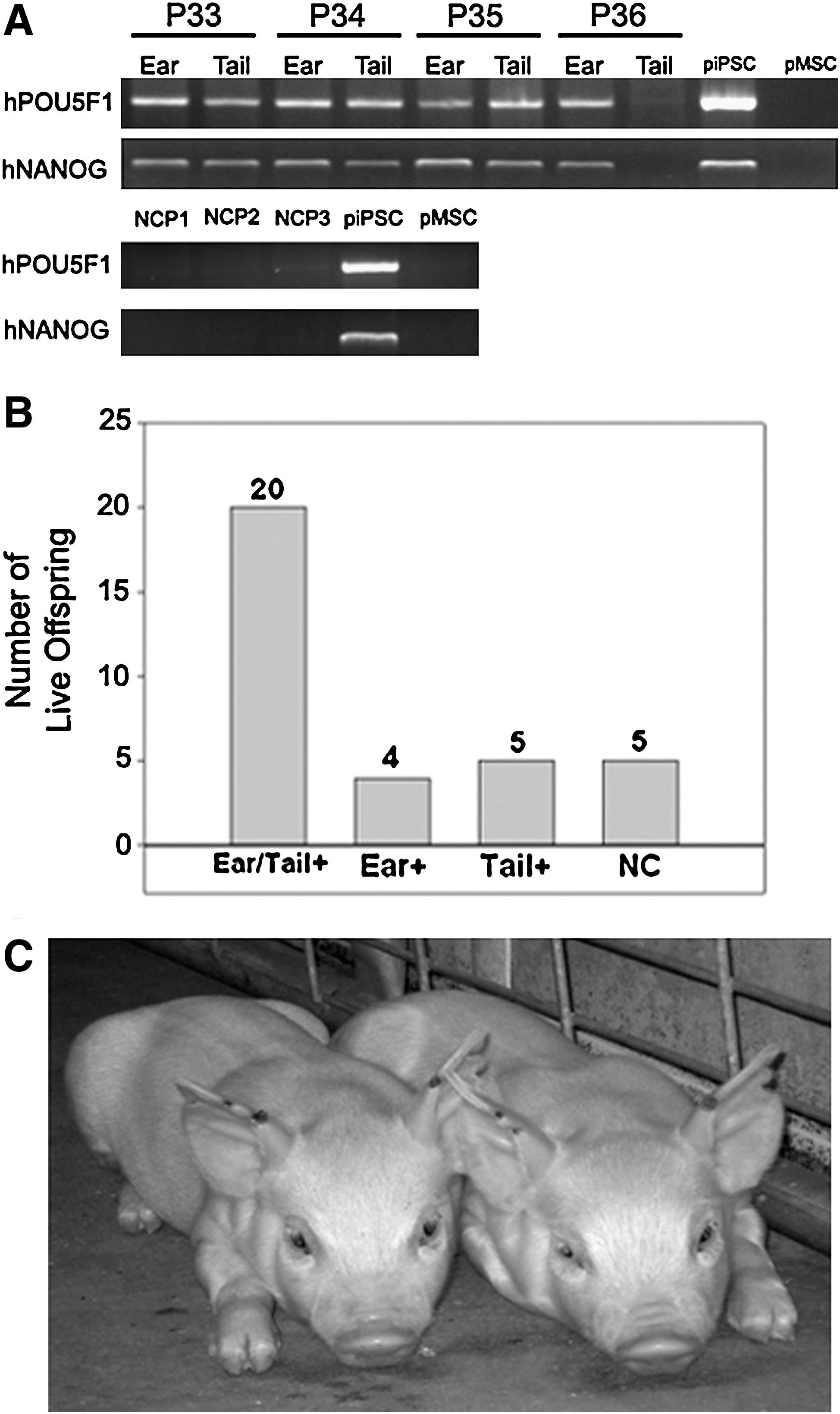
Live offspring born from chimeric embryos. PCR analysis of ear and tail biopsies from 34 live and 2 stillborn pigs demonstrated multiple hPOU5F1/hNANOG+ offspring (
Discussion
In this study we demonstrate for the first time in a nonmurine species that porcine iPSCs are capable of forming chimeras contributing to all 3 germ layers, trophectoderm, and the gonad, and producing live offspring with high efficiency (85.3%). Recent studies have shown the formation of porcine iPSCs that possess morphological immunoreactivity and in vitro differentiation potential similar to pluripotent iPSCs [23 –25]. However, the plasticity of piPSCs remained in question as the ability of these cells to form chimeras indicating complete reprogramming has not been tested. Chimeric formation, not just in vitro differentiation, is an essential assay of pluripotency, as early studies of mouse iPSCs only transduced with Pou5f1, Sox2, C-myc, and Klf4 (as are most current piPSCs [23,24]) displayed many pluripotent characteristics, but were unable to produce adult chimeras, indicating only partial reprogramming [10,14]. The lack of chimerism is potentially due to absence of sufficient NANOG expression, as mouse iPSCs stringently selected on NANOG expression were later able to form chimeras [12]. Further supporting this theory, all chimeras found in the study to be hPOU5F1+ were also hNANOG+. These studies clearly demonstrate the need for chimeric validation to claim a pluripotent status.
The production of live chimeric offspring validates the speculated potential of piPSCs as a tool to study stem cell therapies in a large animal model or for the creation of complex transgenic animals for agricultural and biomedical purposes. These highly proliferative cells with a doubling time of 22.3 h that can be continually expanded for >50 passages without showing any indication of senesces represent a robust cell source capable of differentiating into all 3 germ layers. piPSCs can be coaxed to undergo lineage-specific differentiation and be transplanted into porcine models to study conditions ranging from neural [15] injuries to diabetes to atherosclerosis [16–17] to acute myocardial infarction [18]. Understanding the dynamic cell signaling process between transplanted cells and their damaged or diseased environment leading to successful regeneration and integration is an important aspect of cell therapy that must be determined before cell therapies can be fully utilized. In addition, this system also provides a large animal model to validate efficacy and safety of iPSC cell transplant therapies. Porcine iPSCs were created in this study as the pig has been frequently noted as a superior model (as reviewed in Refs. [26 –29]) relative to murine counterparts due to comparable anatomy and physiology to humans, making them a more biologically relevant model.
The use of iPSC technology in livestock presents a significant tool to conduct complex genetic modifications such as homologous recombination and the knockin and knockout of genes. The use of starting material that is amenable to genetically manipulate, such as pMSCs [19,30] used in this study, can be easily reprogrammed into iPSCs. This would be particularly useful for the production of complex biomedical models that require the manipulation of multiple genes or for agricultural purposes to introduce genes that may confer resistance to diseases or enhance desirable traits. The fact that fetal tissue demonstrated gonad chimerism suggests that genetic manipulations may undergo germline transmission; however, this remains to be validated. In this study, piPSCs also demonstrated contribution to the developing trophectoderm, specifically the placenta. This suggests that piPSCs may possess a level of plasticity even beyond ESCs, which have been traditionally thought to only give rise to lineages derived from the inner cell mass [31,32]. ESCs have only recently demonstrated a limited ability to undergo trophectoderm differentiation [33,34], yet it is frequently a result of extensive genetic manipulation [35 –37]. Aberrant trophectoderm development and maternal–fetal crosstalk in humans commonly result in pregnancy complications (i.e., preeclampsia and intrauterine growth restriction) ending in the loss of the fetus and/or mother [38 –40]. However, the mechanisms causing these diseased states are unclear, potentially creating a niche for piPSCs as a trophectoderm and early developmental model. piPSCs used to create chimeric animals in this study were early passage cells (passage 7), and previous studies in mouse ESCs have demonstrated reduced chimeric contribution with extended culture. Most murine gene targeting studies use early passage cells (<25). Similarly, future genomic modification studies using piPSCs will likely require that pluripotency is maintained at higher passages than 7, but extended cultures will likely be deleterious.
In addition to the formation of chimeras, piPSCs displayed immunoreactivity and morphology similar to murine [10,12,14,41,42], nonhuman primate [43], human [11,13,44], and porcine [23–24] iPSCs from other studies, which are indicative of pluripotency. piPSCs expressed the essential pluripotent transcription factors POU5F1 (both human and porcine) and SOX2, and high levels of AP as do hiPSCs [11,13]. Expression of the pluripotency markers SSEA4 and TRA 1-81 in piPSCs has varied among previous reports. In agreement with Ezashi et al. [24], our piPSCs were weakly positive or negative for SSEA4 and TRA 1-81. However, others have suggested that piPSCs are positive for both SSEA4 and TRA 1-81 [23,25]. In light of these conflicting data, it appears that these 2 markers are not consistent porcine PSC surface markers. Since mouse and human PSC differ in SSEA 1, 3, and 4 marker expression, it is not unexpected that porcine may differ, but all species are consistent in their expression of early transcription factors (POU5F1, NANOG, and SOX2).
piPSCs were derived from MSCs, a unique cell type that is capable of transdifferentiation and can even produce chimeric offspring; however, the degree of MSC transdifferentiation and ability to form chimeric animals from MSCs is limited [45 –48]. It is possible that not all of the pMSCs were fully reprogrammed and therefore contributed to the chimeric offspring. However, the piPSCs appear to possess a high level of plasticity as demonstrated by their contribution to 13 different organ systems spanning all 3 germ layers, extraembryonic tissue, and potentially the germline. This high level of chimerism suggests that it is likely that piPSCs were reprogrammed. Porcine MSCs and piPSCs had stark morphological differences. Reprogrammed cells possessed ESC morphology, expansion, and growth patterns, whereas nonreprogrammed pMSCs lacked the characteristic large nucleoli, high nuclear-to-cytoplasmic ratio, and colonial growth patterns associated with true iPSCs and ESCs. Previous studies have shown that in some instances MSCs are capable of spontaneous expression of POU5F1; however, immunostaining and RT-PCR data showed that pMSCs were POU5F1 negative at multiple passages [45,49,50]. In total, the results suggest that expression of pluripotent markers POU5F1 and SOX2 in piPSCs was a result of the reprogramming process.
piPSCs in this study were also capable of being maintained under hESC feeder-free conditions, which was not previously shown [23 –25], further supporting their ESC character. In fact, feeder-free expansion appeared to be a key step purifying cultures of nonreprogrammed cells—cells with granular or abnormally large cell morphologies. High chimerism efficiency is partially attributed to transplant of these purified and fully reprogrammed cells and the lack of contaminating feeder cells. Porcine iPSCs demonstrate a significant deviation in emergence timing relative to other species. Multiple studies, including this one, have demonstrated earlier than expected emergence of true piPSCs—harvestable nongranular or irregular cells—ranging from 7 to 8 days [23,25]. This is considerably earlier than typical emergence seen in the mouse [10,12] and human [11,13] iPSCs and may represent a species-specific difference. Differences were also noted in doubling time of piPSCs, with one group [24] reporting a significantly faster doubling time (∼17 h) than the piPSCs derived in this (22.3 h) or other (∼24 h) studies and hESCs (28.2 h). Rapid proliferation is a characteristic of ESCs and piPSCs. However, abnormally high proliferation is indicative of a cancerous state and is a significant concern in iPSC formation with the introduction of multiple cancer-associated genes [51 –54] and definitive proof of high levels of tumorigenicity in iPSC chimeric mice [14,55]. To date, tumors have not been noted in chimeric pigs; however, future studies will determine the true tumorigenicity of these cells.
The live birth of 29 chimeric offspring and contribution to all 3 germ layers and potentially the germline demonstrates complete reprogramming of porcine mesenchymal cells into bona fide porcine iPSCs. This is the first report of chimeric offspring being produced in a nonmurine system. Many facets still remain to be explored, including the production of viral-free piPSCs, germline transmission, and the mechanism by which reprogramming occurs. Nonetheless, piPSCs introduce numerous opportunities to study the factors significantly affecting cell therapies, genetic engineering, and other aspects of stem cell and developmental biology.
Footnotes
Acknowledgments
We thank Bradley Didion, Richard Koppang, Ann Marie Paprocki, Charity Syverson, Greg Mell, Gordy Gunderson, and Eric James for their surgical and micromanipulation assistance in the development of chimeric embryos, and Ludwig and Becky Simmet for their generous support. We are also grateful to Erin Jordan and Amber Young, who aided in the isolation of organs/tissues. This work was supported by the Georgia Research Alliance Endowment and the Korea Research Foundation Grant funded by the Korean Government (KRF-2008-357-F00032).
Author Disclosure Statement
No potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.