
Abstract
Embryonic stem cells are considered to be a good in vitro tool to study the induction of various cell types including cardiomyocytes; however, induction of the pharyngeal endoderm (PE), the underlying heart-forming region, in vivo has been scarcely reported. In the present study, we found that many PE-related genes, such as Paxl, Pax9, Sixl, and Tbxl, were up-regulated in cardiomyocyte-rich embryoid bodies (EBs). The third pouch-related genes including Hoxa3, Foxn1, and Aire, which are crucial for thymus development and function, were also detected in later stages. Nkx2.5, a cardiac transcription factor gene, is known to be transiently expressed in the PE. By crossing Nkx2.5-Cre mice with Cre-dependent EGFP reporter mice, we found that Nkx2.5+ lineage exclusively contributed to thymic epithelial cell development, followed by thymus development. Gene expression analysis using Nkx2.5-EGFP ES cells also revealed that PE-related mRNAs were specifically enriched in the transiently appearing E-cadherin+Nkx2.5+ cell fraction. Interestingly, the EB-derived cells were found capable of supporting T-cell differentiation to CD4 or CD8 double-positive cells in a reaggregation organ culture in vitro. Our results suggest that EBs contain cells that resemble third pharyngeal pouch endoderm and confer a thymus-like microenvironment.
Introduction
D
In the pharyngeal region where cells express Pax1 and Pax9, lateral pockets of endoderm called pharyngeal pouches (PPs) are formed on around E9.5 during mouse development [6]. Hoxa3 is also expressed in PPs, though in a caudal domain (third and fourth PPs), and is regulated by retinoic acid signaling. In the third PP, the parathyroid and thymus are developed, and their prospective regions are marked by the expression of Gcm2 and Foxnl, respectively [7], which can be detected beginning on E9.5 and E11.5, respectively. Loss of those leads to loss of their related organ. In addition, Six1 and Eya1 have been shown to be crucial for development of the thymus and parathyroid [8]. Together, these results have been useful to clarify the genetic cascades important for thymus and parathyroid organogenesis. However, induction of the PE and PPs from embryonic stem (ES) cells has been scarcely reported.
We previously showed that transient serum removal during differentiation of ES cells efficiently enhanced the expression of cardiogenesis-associated genes in embryoid bodies (EBs) [9]. In the present study, we found that the PE-related genes Pax1, Pax9, Eya1, Six1, Tbx1, as well as the PP-related genes Foxn1, Aire, Hox3a, were also induced in cultures with cardiogenic conditions. Interestingly, those EBs were able to support T-cell differentiation in vitro. Thus, our findings indicate possible opportunities to produce thymic epithelial cells (TECs) in vitro.
Materials and Methods
ES cell maintenance and differentiation
hcgp7 (ht7-derived Nkx2.5-GFP knock-in ES cells) and EB3 (a kind gift from Dr. Niwa) cells were cultured without feeders in leukemia inhibitory factor (LIF)-supplemented medium, as described previously [10]. These ES strains carry hygromycin- or blasticidin-resistant genes in one of the foci of Oct4 (known as Pou5f1), which are used to select undifferentiated cells during maintenance. On the other hand, the hygromycin-resistant gene is fused to the herpes simplex virus thymidine kinase (HSV-TK) gene. Therefore, hcgp7-derived undifferentiated cells can be eliminated by administration of ganciclovir [11]. To differentiate the ES cells, EBs were formed by hanging drops and cultured as floating EBs in Glasgow's modified Eagle's medium (GMEM)-based differentiation medium containing 10% fetal bovine serum (FBS) and 0.1 mM 2-mercaptoethanol (2-ME). As previously reported by us and others, removal of serum on Day 4 of the culture (d4) enhanced cardiac induction on d7 (see Supplementary Fig. 1; Supplementary materials available online at

Pharyngeal endoderm-related genes expressed in embryoid bodies (EBs) and up-regulated in cardiogenesis-enhancing condition. (
Whole-mount in situ hybridization
Whole-mount in situ hybridization was performed as described previously [9]. Digoxigenin-labeled complementary RNA probes were synthesized from plasmids harboring the complementary DNA (cDNA) sequence and hybridized with mouse embryos. The following primers were used for polymerase chain reaction (PCR) assays and cloned into a pCR4-TOPO vector (Invitrogen): Pax1 sense, 5′-ACCACCCCCAATGTAGTGAA-3′ and antisense, 5′-GTCCCTTCTCGGTGTTTGAA-3′; Vgll2 sense, 5′-GAGCCATGAGCTGTCTGGAT-3′ and antisense, 5′-AGAGTGGCGG GAGAGTAGG-3′.
Flow cytometry and cell sorting
Floating or attached EBs were harvested and dissociated into single cells with trypsin (0.25% trypsin-ethylenediaminetetraacetic acid (EDTA); Invitrogen). Flow cytometry analyses were carried out using a FACS Canto flow cytometer (BD Biosciences, San Jose, CA). Cell sorting was performed using a FACS Aria cell sorter (BD Biosciences). Data were analyzed using FlowJo (Tree Star, Inc., Ashland, OR) software. The following antibodies were used: anti-E-cadherin antibody (rat monoclonal, clone ECCD-2; Takara Shuzo Co., Ltd., Otsu, Shiga, Japan), allophycocyanin (APC)-conjugated anti-rat IgG antibody (BD Biosciences), and R-phycoerythrin (PE)-conjugated anti-EpCAM antibody (rat monoclonal, clone G8.8; eBioscience, San Diego, CA).
Quantitative reverse transcription PCR
For expression studies, EBs and EB-derived cells were harvested, and total RNA was extracted with Trizol Reagent (Invitrogen). After treating the RNA samples with deoxyribonuclease, cDNA was synthesized using SuperScript III reverse transcriptase (Invitrogen). Real-time PCR was performed using an ABI7900 (Applied Biosystems, Foster City, CA) with SYBR Green Master Mix (Invitrogen) and Taqman Rodent GAPDH Control Reagents (Applied Biosystems). Expression data were normalized to GAPDH expression. The primers are described in Table 1.
Reaggregation thymus organ culture
Co-cultures of EB-derived cells and thymocytes were performed as described previously [14]. In brief, 3–5 × 105 embryonic thymocytes were mixed with an equal number of EB-derived cells and spun down. The cell slurry was placed on the surface of a polycarbonate filter, which was supported by a collagen sponge. Mouse embryonic fibroblasts (MEF) and 2′-deoxyguanosine (dGuo)-treated thymic stromal cells were used as negative and positive controls, respectively. After 1 week of incubation, the thymocytes were harvested and analyzed by flow cytometry.
Immunofluorescence
Cytospin-acquired EB-derived cells were fixed with 4% paraformaldehyde for 10 min. Cells were incubated with a rabbit polyclonal anti-Foxn1 antibody [15] for 60 min, washed twice, and incubated with an anti-rabbit IgG antibody conjugated with fluorescein isothiocyanate (FITC) for 30 min. After staining, the cells were washed twice and fluorescence was visualized with a Leica TCS SP2 confocal microscope.
Animals and transplantation of EBs under kidney capsule
For a cell lineage tracking study, Nkx2.5-Cre andCAG-loxP-CAT-loxP compound heterozygous mice were generated by crossing mice carrying each genotype [16,17]. For a transplantation study, Balb/c nu/nu mice were used as recipients. Twelve-day-old EBs derived from hcgp7 were placed under the kidney capsule with a glass capillary. To eliminate HSV-TK+ undifferentiated cells, ganciclovir was administrated intraperitoneally at 50 mg/kg from d1 to d10 [18]. One month following transplantation, thymocytes were harvested from the grafts and analyzed by flow cytometry. The animals were handled in accordance with the Guidelines for Animal Experiments of the National Cardiovascular Center.
Results
Transient serum removal induced PE genes in EBs
Previously, we found that serum removal from EB cultures efficiently enhanced cardiac differentiation [9]. Microarray analysis using cardiomyocyte-rich EBs revealed that a set of transcription factor genes important for heart development and differentiation, Nkx2.5, Gata4/5/6, Mef2c, and Myocd, were induced under this condition. Furthermore, screening of genes showing an Nkx2.5-like expression pattern identified several noncardiac genes, including Pax1 and Vgll2 (Fig. 1A). Consistent with a previous report [19], expression of those genes was detected in the pharyngeal region and somites, but not in the cardiac region, by in situ hybridization (Fig. 1B). On the other hand, our experimental culture conditions did not generate a skeletal muscle lineage, as shown by our microarray data (Supplementary Fig. 1; Supplementary materials available online at
Expression of third PP genes and proteins in EBs
The PE and one of its derivatives the third PP are known to play pivotal roles in thymus organogenesis. The Foxn1 transcription factor is crucial for development of the mature thymus and its expression begins in the thymus domain of the third PP on E11.25. In contrast to PE genes, expression of the Foxn1 gene was barely detected in floating EBs on d4 to d8. As shown in Figure 2A, a SFA culture condition tended to enhance the expression of Foxn1 in later stages (d12). Many other epithelial and/or mesenchymal genes known to be important for thymus organogenesis and function, including Aire, β5t, Ccl21, Ccl25, Dll1, and Dll4, were also detected. The expression levels determined by quantitative reverse transcription PCR (qRT-PCR) were found to be much lower than that in the E14.5 embryonic thymi (<1/100 for Foxn1), though we observed that the EpCAM+ cell fractions had enriched expressions of Foxn1 mRNA (>1/30), as well as of Pax1 and Pax9 (data not shown). To confirm that Foxn1 is expressed at the protein level, we stained cytospin preparations of EBs with the Foxn1 antibody [15]. Clear nuclear signals were detected in SFA-cultured EBs on d13, but not in floating EBs on d4, though positive cells were much less abundant than in the embryonic thymus (Fig. 2B).
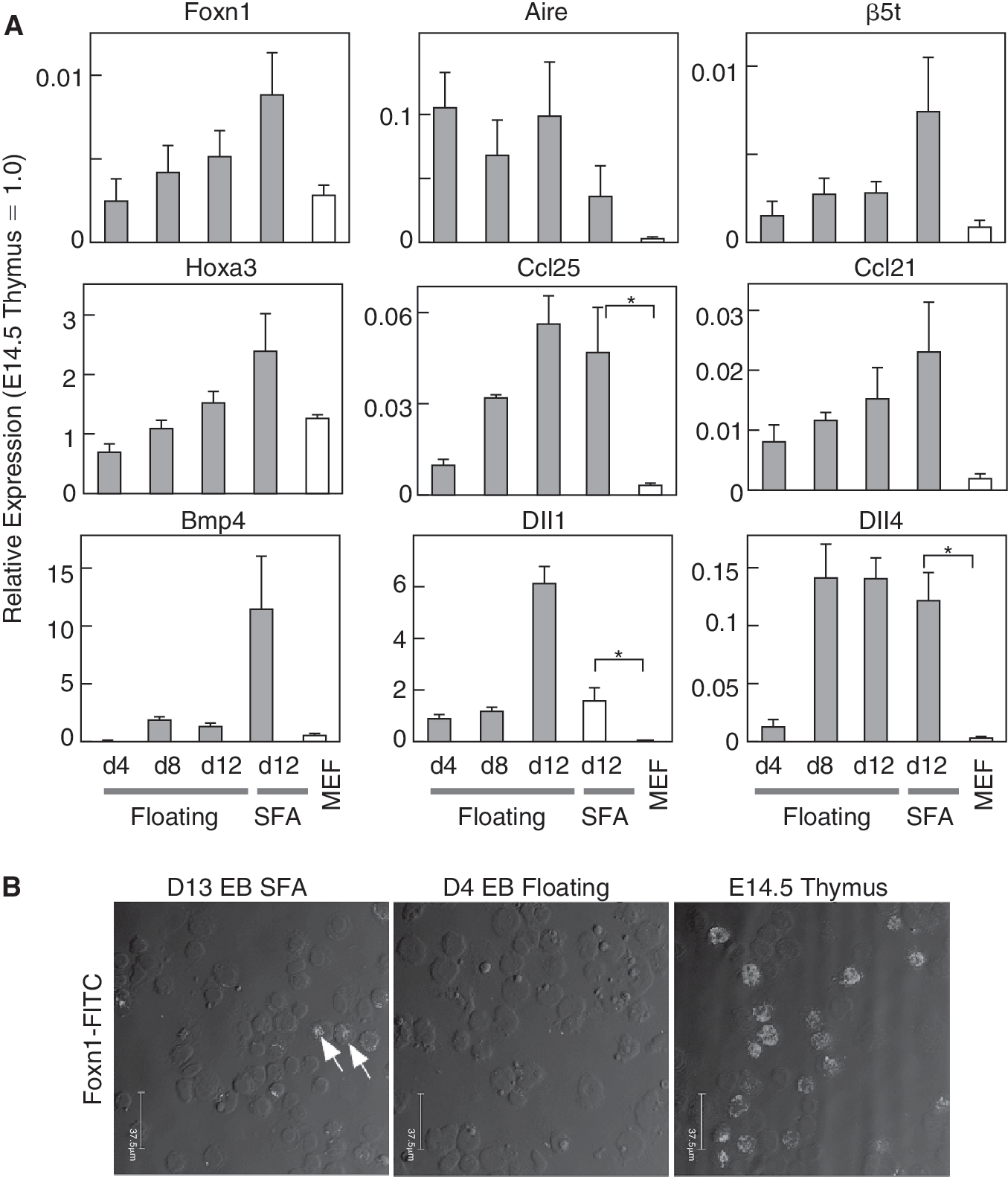
Expressions of Foxnl and other markers important for thymus function in embryoid bodies (EBs). (
Enrichment of PE gene expressing cells in Ecad+Nkx2.5+ cell fraction
Previous reports have shown that Nkx2.5 is not specifically expressed in the heart tube, whereas its expression has been detected in extracardiac tissues including the PE (Fig. 3A and [24]). In contrast to Nkx2.6, the expression of Nkx2.5 declines after formation of PPs on E9.5, though both play pivotal roles in the induction of pharyngeal genes [25]. Previously, we established hcgp7 cells (Nkx2.5-GFP knock-in ES cells, Fig. 3B) and found that the GFP+ cell fraction specifically enriched cardiomyocytes with a very high purity [10]. If PE cells are generated by EBs, they may be included in the GFP+ cell fraction, thus we used E-cadherin (Ecad) as a marker to identify such endodermal cells [26]. Flow cytometry analysis of EBs on d6 revealed that a small population of cells expressing both GFP and Ecad (1% of live cells) was transiently generated (Fig. 3C). As shown in Figure 3D, Ecad+GFP+-sorted cells preferentially expressed Pax1, Pax9, Eya1, Six1, Vgll2, and Fgf8, indicating that these genes were co-expressed in the limited cell fraction. These results confirmed that PE cells were indeed generated by the EBs.

Pharyngeal endoderm marker-expressing cells enriched in Ecad+Nkx2.5+ cell population. (
Nkx2.5+ lineage differentiates into TECs
Expression of Nkx2.5 in extracardiac tissues, such as the thyroid, spleen, and stomach, has been reported. However, the expression of Nkx2.5 in the thymus or contribution of Nkx2.5+ lineage to thymus development has not been adequately examined [17]. To investigate the fate of Nkx2.5+ lineage in the thymus, we analyzed thymic cells from Nkx2.5-Cre/loxP-CAT-loxP-EGFP double transgenic mice to detect cells that expressed Nkx2.5 (Fig. 4). Approximately 6% of all cells were GFP+ and, remarkably, all of the GFP+ cells were composed of EpCAM+, CD31−, and PDGFRα−, indicating that they were epithelial cells, and not endothelial or mesenchymal cells. These results suggest that Nkx2.5+ lineage contributes to the development of TECs in the thymus, consistent with results of its expression in the PE. Furthermore, 65% of the GFP+ cells were I-Ab+, a molecule that characterizes functional TECs. Our findings indicate that Nkx2.5+ PE-derived cells are present as TECs in the embryonic thymus.
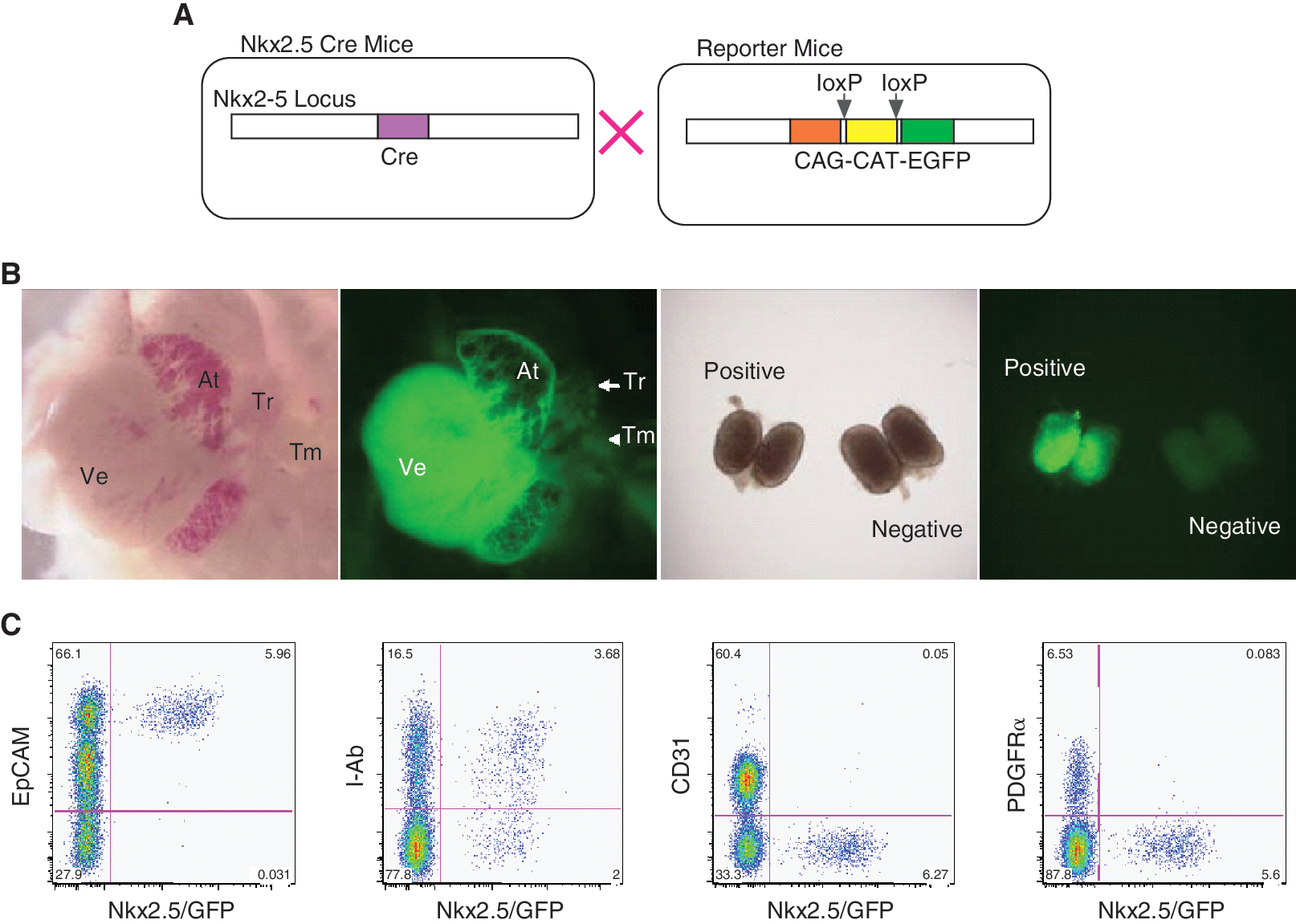
Nkx2.5+ lineage differentiated into TECs in vivo. (
EBs support thymocyte differentiation in vitro
Thymic epithelial cells are known to support differentiation of T cells when cultured as aggregates in vitro. Using the well-known co-culture system termed reaggregation thymus organ culture (RTOC), we next examined if EBs can support the differentiation of T cells. As shown in Figure 5, when immature CD4−CD8− thymocytes derived from E15.5 embryonic thymi were incubated with EBs on d12 and d13, they differentiated into CD4+CD8+ double-positive T cells, as well as CD4+ or CD8+ single positive cells within 1 week. In contrast, mouse embryonic fibroblasts (MEFs) did not have that activity. These findings are consistent with the results showing that EBs expressed TEC genes at a low level, though higher than the control MEF. Collectively, our results indicate that EBs can provide a microenvironment that supports T-cell differentiation, by inducing TEC-like cells.

Embryoid bodies (EBs) support thymocyte differentiation in vitro. Embryoid body-derived dissociated cells (d12 EBs) were dissociated and mixed with embryonic thymocytes (E15.5). After culturing as a slurry on a polycarbonate membrane for 1 week, thymocytes were harvested for analysis by flow cytometry. Adult thymocytes and embryonic thymocytes were separately cultured with 2′-deoxyguanosine-treated thymic stromal cells (dGuo-TSC) and used as positive controls, while embryonic thymocytes cultured with mouse embryonic fibroblasts (MEFs) and EBs without embryonic thymocytes were used as negative controls. Thymocytes recovered from dGuo-TSC, EB, and MEF samples numbered approximately 1 × 104, 1 × 105, and 5 × 105, respectively.
To test if the function of TECs might be detectable in vivo, we next analyzed thymocyte development in grafts from nude mice. Since unsorted EBs contain undifferentiated ES cells that form tumors after transplantation, it was important to purify the TEC-like cells or eliminate undifferentiated cells. Since EB-derived cells scarcely express I-Ab even in late stages (data not shown), we took the latter approach. The ES strain hcgp7 harbors the HSV-TK gene in one of the loci of Oct4. Thus, undifferentiated ES cells expressing HSV-TK are killed by an administration of ganciclovir. As shown in Figure 6, we observed tumors in only 1 of 8 ganciclovir-administered nude mice, while 100% (3 of 3) of the nonadministered mice manifested tumors. Administration of ganciclovir itself did not affect the survival or function of the transplanted thymic lobes. Thus, elimination of undifferentiated cells turned out to be a successful strategy. However, we were unable to detect T-cell differentiation in nude mice grafted with EB-derived cells.

EBs do not support thymocyte differentiation in vivo.
Discussion
In the present study, we found that serum removal, which enhances cardiogenesis in EB cultures, induced the expression of not only cardiac genes, but also the PE genes Tbx1, Pax1/9, Six1, and Eya1. We also detected the expression of another PE gene, Vgll2, though it did not seem to be involved in PE development, based on our recent analysis of knockout mice (data not shown). Importantly, these markers were enriched in the same cell fraction that expressed Nkx2.5 (GFP) and Ecad, suggesting that PE cells are indeed generated by EBs. Although the cardiac transcription factor gene Nkx2.5 is known to be expressed in the PE, its expression declines following pouch development and it is not detected in the thymus. Using Crelox technology, we showed for the first time that a substantial number of Nkx2.5+ lineage cells contribute to TEC development. It would be intriguing to know when PE-derived cells begin to contribute that development, and where and how Nkx2.5− TECs arise. Nevertheless, our findings provide further insight into PE and PP development.
Weak Foxn1 expression was detected in long-term cultures (12 days or more); however, there was no Gcm2 expression seen (data not shown). During mouse embryonic development, Gcm2 is first detected on E9.5 in the third pouch, about 2 days earlier than Foxn1. Although both the thymus and parathyroid are generated from the third PP endoderm, their requirements for induction may be distinct, with BMP signaling implicated in TEC differentiation and thymus organogenesis [27]. It is also known that the expression of Bmp4 is restricted to the ventral region of the third pouch endoderm, which eventually expresses Foxn1 [28]. With the culture conditions used in the present study, BMP-responsive genes, such as Nkx2.5 and Idb1/2 [29,30], were efficiently up-regulated. Thus, it is possible that the level or strength of BMP signaling suitable for cardiac differentiation may also be suitable for induction of the portion of the PE specified to develop into TECs.
Interestingly, we found that EBs were able to support T-cell differentiation in vitro, which was surprising, because the expression level of TEC markers in EBs was much lower than that of those in the embryonic thymus (ie, <1/100 for Foxn1). Thus, only a small population of TEC-like cells are likely capable of inducing T-cell differentiation. However, the EBs in the present study could not rescue T-cell differentiation in nude mice. We are uncertain if the TEC-like cells among the EBs were still immature or too small in number to function in vivo. Alternatively, the immature TEC progenitor might have been eliminated together with undifferentiated ES cells by ganciclovir treatment in the present experiment. Recent studies have found that TEC progenitors express Oct4 [31]. Thus, it is possible that Oct4 is required for the function of TECs at some points of development. Enhancing differentiation efficiency as well as the use of appropriate markers for selection are required for future studies.
Based on results of the present gene expression analysis, cells with PE identities were strongly suggested to have been developed from EB-derived cells. Our findings contribute important evidence regarding the potential of ES cells and other pluripotent stem cells (such as iPS cells) to produce thymic epithelial progenitor cells [32]. Thymic epithelial cells form the stromal microenvironment, which is crucial for thymopoiesis, but are gradually lost over time, resulting in a restricted T-cell receptor repertoire. Regeneration and repair of the thymic niche may become an important target for immune reconstitution, especially in immune-compromised individuals. Studies are now in progress to detect other differentiation markers and functional TECs derived from ES cells.
Footnotes
Acknowledgments
We express our thanks to Ms. Sachiko Suzuki and other members of the Department of Bioscience for their assistance with these experiments. This work was supported in part by the Program for Promotion of Fundamental Studies in Health Sciences of the National Institute of Biomedical Innovation (NIBIO), and by Health Labour Sciences Research Grants and Research Grants for Cardiovascular Diseases from the Ministry of Health, Labour and Welfare, Japan.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.