
Abstract
Mesenchymal stem cells (MSCs) found in bone marrow (BM)-MSCs are an attractive source for the regeneration of damaged tissues. Alternative postnatal, perinatal, and fetal sources of MSCs are also under intensive investigation. MSCs from the Wharton's jelly matrix of umbilical cord (WJ)-MSCs have higher pancreatic and endothelial differentiation potentials than BM-MSCs, but the underlying mechanisms are poorly understood. We compared the gene expression profiles, enriched canonical pathways, and genetic networks of BM-MSCs and WJ-MSCs. WJ-MSCs express more angiogenesis- and growth-related genes including epidermal growth factor and FLT1, whereas BM-MSCs express more osteogenic genes such as RUNX2, DLX5, and NPR3. The gene expression pattern of BM-MSCs is more similar to osteoblasts than WJ-MSCs, suggesting a better osteogenic potential. In contrast, WJ-MSCs are more primitive because they share more common genes with embryonic stem cells. BM-MSCs are more sensitive to environmental stimulations because their molecular signatures altered more significantly in different culture conditions. WJ-MSCs express genes enriched in vascular endothelial growth factor and PI3K-NFκB canonical pathways, whereas BM-MSCs express genes involved in antigen presentation and chemokine/cytokine pathways. Drylab results could be verified by wetlab experiments, in which BM-MSCs were more efficient in osteogenic and adipogenic differentiation, whereas WJ-MSCs proliferated better. WJ-MSCs thus constitute a promising option for angiogenesis, whereas BM-MSCs in bone remodeling. Our results reveal systematically the underlying genes and regulatory networks of 2 MSCs from unique ontological and anatomical origins, as well as the resulted phenotypes, thereby providing a better basis for cell-based therapy and the following mechanistic studies on MSC biology.
Introduction
S
The umbilical cord vessels and the surrounding mesenchyme [including the Wharton's jelly (WJ) connective tissue matrix] are derived from extraembryonic mesoderm and embryonic mesoderm. Several reports showed that MSCs with pluripotent differentiation potential can be isolated from the matrix part of WJ [5,6]. Postnatal stem cells derived from WJ can give rise to various lineages of cells, including adipocytes, chondrocytes, osteoblasts, and cardiomyocytes [6 –8]. In addition, fibroblast-like cells of WJ can be induced into neural-like cells expressing neuron-specific enolase, as well as other neural cell markers [5]. Several groups further showed that WJ-derived MSCs (WJ-MSCs) can be induced to dopaminergic neurons in vitro and have the potential for the treatment of Parkinson's disease [5,9,10] or spinal cord trauma [11]. WJ-MSCs are also capable of pancreatic and endothelial differentiation [12,13]. WJ-MSCs form larger islet-like cell clusters than BM-MSCs [12], and endothelial differentiated WJ-MSCs have significantly higher total tubule length, diameter, and area than differentiated BM-MSCs [13]. Also, WJ-MSCs have higher proliferative potential than BM-MSCs [13]. Although WJ-MSCs can differentiate into cells of the mesenchymal lineage [5,6], their osteogenesis ability has not been compared with that of BM-MSCs on a side-by-side manner.
Although the immunophenotypic profiles of MSCs from different tissues are very similar [2,5,6], the disparate characteristics of MSCs from different anatomic locations should be considered to better apply MSCs clinically, because the origin or source of MSCs will determine their fate and functional characteristics. Monitoring genome-wide characters offers an opportunity to discover unique property and underlying regulatory networks in stem cells and holds great promise for the future of regenerative medicine. Comparative analysis between MSCs derived from BM, umbilical cord blood, and adipose tissue has been conducted [4,14]. The gene expression profiles, as well as their comparisons, of various human MSCs have also been reported by quite a few studies [3,15 –18]. Differences in gene expression profiles and cellular function between human WJ-MSCs and BM-MSCs remain yet to be explored. A very recent stem cell marker analysis on long-term cultured MSCs showed that human WJ-MSCs express more ESC markers (such as NANOG, DNMT3B, and GABRB3) and some early endodermal markers at both early and late passages [19]. WJ-MSCs thereby possess properties of true stem cells, which they retain even after extended in vitro culturing.
In this study we applied systems biology tools to investigate the global transcriptome as well as the functional differences between BM-MSCs and WJ-MSCs and to provide the responsible genes and functional genetic networks. We show by functional module analysis why WJ-MSCs may differentiate into endothelial cells more efficiently, and drylab analyses further suggest BM-MSCs to be a better source for osteoblasts. The basic biological potentials (such as osteogenic differential potential) of different MSCs remain unchanged in 2 tested culture conditions, although the expression of certain genes may change in response to in vitro culture conditions. Drylab conclusions can be verified by wetlab differentiation assays. Our data thereby contribute new insights into a refined molecular picture and a better understanding of these 2 MSC subtypes. Results obtained will benefit cell-based therapy, as well as subsequent mechanistic studies in angiogenesis and osteogenesis.
Materials and Methods
Isolation and cultivation of human MSCs
This research follows the tenets of the Declaration of Helsinki and informed consent was obtained from the donor patients. All human MSCs used for experiment were cultured for <8 passages in the MesenCult™ medium (StemCell Technologies) in the presence of 5% CO2, or in Dulbecco's modified Eagle's medium (DMEM; Cat. No. 12100-061; Gibco-BRL) supplemented with 10% fetal bovine serum (FBS; Cat. No. 12003; JRH Bioscience). For BM-MSC isolation, the mononuclear fraction of BM was isolated by Histopaque-1077 gradient (1077-1; Sigma) and plated in the culture medium. Nonadherent cells were discarded on next day. For array analysis, BM-MSCs were isolated from 2 male and 2 female donors with age under 40, and those used in reverse transcription polymerase chain reaction (RT-PCR) and differentiation experiments were isolated from both male and female donors with age 40–70.
MSCs from WJ were collected as published [9,20]. Briefly, umbilical cords (UCs) were processed within 24 h after delivery. WJ tissues were isolated from UCs and then digested with collagenase and trypsin. Cells were washed twice with phosphate-buffered saline (PBS) and plated on regular culture dishes in fresh culture medium; nonadherent cells were discarded on next day. Approximately 4 days to 1 week after explantation, fibroblast-like adherent cells were detached using trypsin–ethylenediaminetetraacetic acid solution and then reseeded in fresh culture medium without dilution for further expansion [passage 1 (P1)].
Array probe preparation and data processing
Osteoblast, adipocyte, and chondrocyte array data were downloaded from the Gene Expression Omnibus (
Genetic network analysis
Differential gene expression profiles were imported into the Ingenuity Pathways Analysis (IPA) software (Ingenuity Systems;
Immunofluorescent assay and flow cytometry
For flow cytometry, 5 × 105 MSCs in PBS were incubated with monoclonal antibodies of anti-CD29 (Cat. No. 303004; Biolegend), CD34 (Cat. No. MCA547PE; Serotec), CD44 (Cat. No. 312306; Biolegend), CD45 (Cat. No. 304006; Biolegend), CD73 (Cat. No. 550257; BD Pharmingen), or anti-CD90 (Cat. No. MCA90F; Serotec) on ice for 1 h. Unstained MSCs were used as a negative control.
RNA isolation and real-time PCR
Total mRNA was extracted using RNeasy mini kit (Cat. No. 74106; Qiagen) and 100 ng to 1 μg of total RNA was reverse transcribed using First cDNA Synthesis Kit (Cat. No. K1612; Fermentas). For quantitative real-time PCR analysis, human premessenger RNA sequence was obtained from the NCBI AceView program (
Proliferation assay
BM-MSCs and WJ-MSCs were started at passage 3 and plated at a density of 2 × 103 cells/cm2 in MesenCult medium. Cells were subcultured every 5 days and counted using a hemocytometer. Cumulative population doublings were calculated as published [25].
Multilineage differentiation assays
Osteogenic differentiation
MSCs were plated at 5 × 103 cells/cm2 in 6-well plates and treated with osteogenic medium for 4 weeks. Osteogenic medium consists of 0.1 μM dexamethasone, 10 mM β-glycerolphosphate, and 50 μM ascorbic acid in DMEM supplemented with 10% FBS (all chemicals were purchased from Sigma-Aldrich). Osteogenic differentiation was determined by alkaline phosphatase staining (Cat. No. 86R-1KT; Sigma). For evaluation of mineralized matrix, differentiated cells were treated with 2% silver nitrate (Cat. No. S6506; Sigma) under UV lamp for 1 h. Cells were then fixed with 2.5% sodium thiosulphate (Cat. No. S7026; Sigma) for 5 min and washed with water. Finally, cells were counterstained with 1% neutral red (Cat. No. N2880; Sigma) for 1 min.
Adipogenic differentiation
About 5 × 103 MSCs/cm2 were induced in adipocytic differentiation medium (1 μM dexamethasone, 0.5 mM 3-isobutyl-1-methylxanthine (IBMX), 10 μg/mL insulin, and 100 μM indomethacin in DMEM with 10% FBS). The medium was changed twice a week. Adipogenic differentiation was determined by Oil Red O staining. The cells were fixed with 10% formalin in PBS for 5 min at room temperature. After washing with 60% isopropanol, the cells were stained by Oil Red O solution (Cat. No. O0625; Sigma) (0.2% Oil Red O in 60% isopropanol) for 10 min at room temperature. All differentiation experiments were repeated 3 times using 3 batches of MSCs from 3 different donors. Around 90% of BM-MSCs could be differentiated into adipogenic cells at week 3 in each batch of cells.
Immunoblotting
Proteins from total cell lysates were resolved by sodium dodecyl sulphate–polyacrylamide gel electrophoresis, transferred to polyvinylidene fluoride (PVDF) membrane, and then blocked with 5% nonfat milk in PBS/Tween-20 or 5% bovine serum albumin in Tris-buffered saline–Tween-20, according to the instructions for antibodies. For immunoblotting, the following antibodies were used: phosphor-NFκB-p65 (Ser536) (No. 3031; Cell Signaling), NFκB p65 (GTX107678; GeneTex), IκB-α (C-21) (sc-371; Santa Cruz), and glyceraldehyde 3-phosphate dehydrogenase (MAB374; Millipore). Band intensity was measured by the ImageQuant software.
Macrophage preparation and coculture with MSCs
Human CD14+ monocytes were purified from peripheral blood mononuclear cells and monocyte-derived macrophages were cultured as described previously [26]. About 3 × 104 MSCs (upper inserts) and 4–5 × 105 macrophages (lower chamber) isolated from unrelated donors were cocultured using 12-well Transwell plates (No. 3460; Corning) for 2 days, and 1 μg/mL E.coli lipopolysaccharide (LPS; Sigma) was then added to the lower chamber for activating macrophages. Sixteen hours later, tumour necrosis factor-α and interleukin 6 pro-inflammatory cytokines expressed by macrophages were analyzed by quantitative (q)RT-PCR.
Results
Characterization of MSCs from WJ or human BM
To access molecular signatures for MSCs from different sources, primary MSCs from human BM or umbilical cord WJ were collected. Both MSC types shared a similar fibroblastoid-shaped morphology (Fig. 1A). Flow cytometry analyses of cells less than passage 8 revealed that both BM-MSCs and WJ-MSCs expressed known MSC markers such as CD29 and CD73 (SH3/4), but were negative for hematopoietic CD14 marker and CD34 (Fig. 1B, C). Both MSCs were also positive for CD44 but negative for CD45 (not shown).
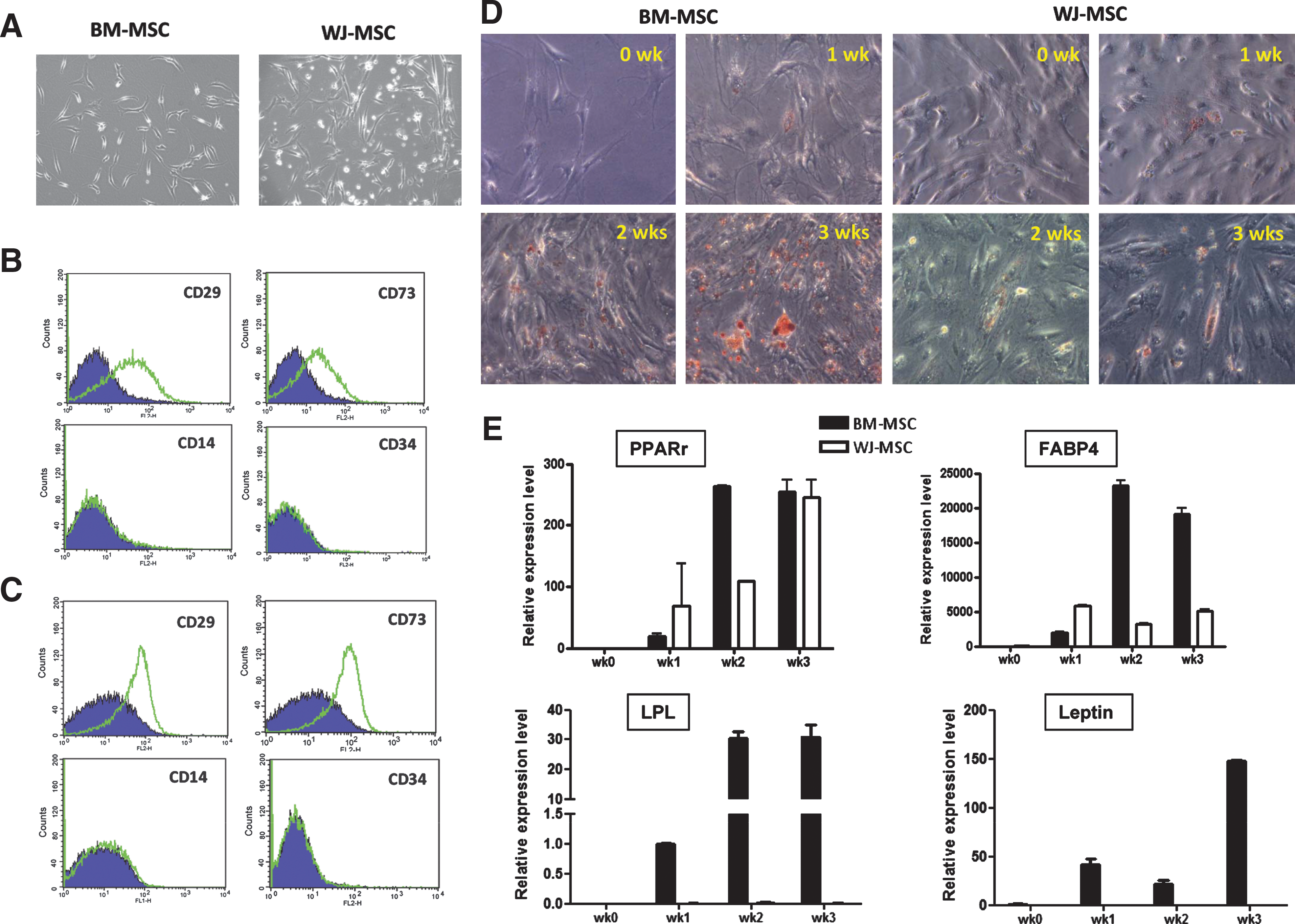
Characterization of 2 mesenchymal stem cell (MSC) subtypes. (
As a pilot study to test whether there is any functional difference between these 2 MSC subtypes, we tested their adipogenic potentials. Adipogenic differentiation was judged as Oil Red O-positive cells and was also analyzed by qPCR, in which the adipocytic marker genes PPARγ and FABP4 were expressed in the adipogenic formula-treated cells. Differentiated adipogenic cells could be observed in both MSCs at week 1, but lipid accumulation was faster in BM-MSCs (Fig. 1D). More differentiated BM-MSCs stained positive for Oil Red O at weeks 2 and 3 (Fig. 1D). Four examined adipogenic markers, PPARγ, FABP4, LPL, and Leptin, were also more abundantly expressed in BM-MSCs than in WJ-MSCs during differentiation (Fig. 1E).
Functional transcriptome analysis on different MSCs
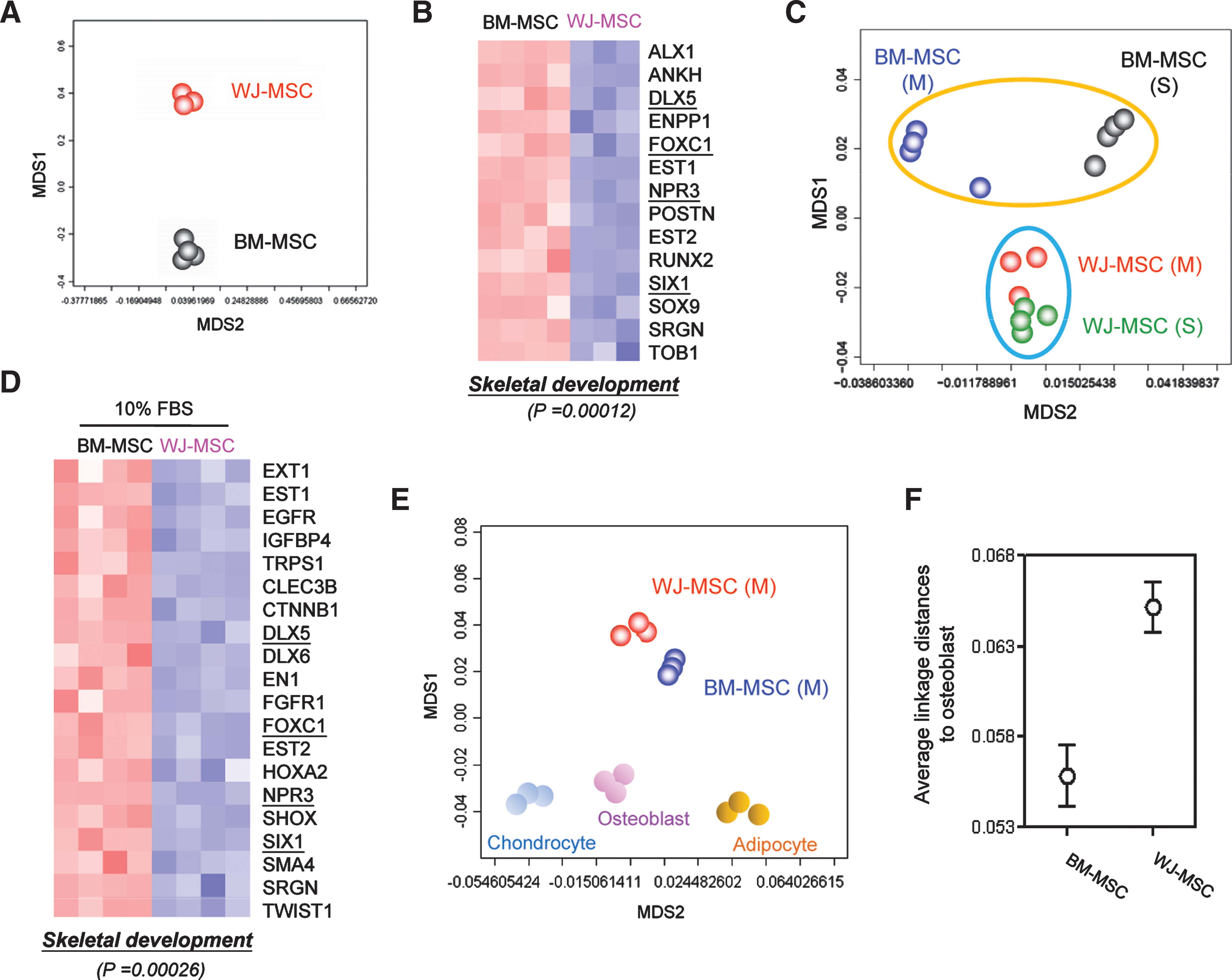
To provide more insights into the functional differences of different MSCs, we performed transcriptome analysis on them. Genes differentially expressed between each cell type (the molecular signatures) were identified according to a statistical pipeline used [21,22]. There were 762 probe sets (corresponding to 454 unique genes) abundantly overexpressed in WJ-MSCs, whereas another 334 probe sets (175 genes) in BM-MSCs [with a positive false discovery rate (pFDR) threshold q < 0.001]. An MDS plot using these genes illustrated the grouping power of them (Fig. 2A). The top 50 known genes most strongly expressed in each MSC are listed in Table 1. In WJ-MSCs, known neurogenic genes such as somatostatin receptor 1 [27], NRXN3 (neurexin 3) [28], RTN1 (reticulon 1) [29] and spondin 2 (M-spondin) [30] are expressed by those 50 genes (Table 1A, underlined). Also in the list is natriuretic peptide precursor B (NPPB), which is expressed in the early embryonic nervous system and may play region- and stage-specific roles during the development of the peripheral and central nervous systems [31]. Genes involved in cell proliferation, including cyclin D2 (CCND2), epidermal growth factor (EGF), and FLT1 (Fms-related tyrosine kinase 1; vascular endothelial growth factor receptor), are also enriched in WJ-MSCs. ICAM1 (intercellular adhesion molecule 1; CD54), an endothelial marker involved in cell adhesion, is also overexpressed in WJ-MSCs (Table 1A, underlined). Genes involved in blood vessel development, such as FOXF1 and NPPB, are enriched in WJ-MSCs.
BM-MSCs express more genes involved in osteogenesis. (
In BM-MSCs, known angiogenic genes (such as ANGPT1 and PGF) are present (Table 1B, underlined). CXCL12 (also known as stromal cell-derived factor-1, SDF1) is involved in the migration and trafficking of BM-MSCs, endothelial progenitor cells, and muscle progenitors [32,33]. It is known that migration of BM and cord blood MSCs in vitro is regulated by the SDF1-CXCR4 and the hepatocyte growth factor-c-Met axes [33]. PRRX1 (paired related homeobox 1; alias PHOX1) is a homeobox-containing protein that is expressed in the mesenchyme of facial, limb, and vertebral skeletal precursors during embryogenesis. PRRX1 is required for early events of skeletogenesis as well as in molar tooth morphogenesis [34,35]. Also enriched in BM-MSCs is NPR3 (natriuretic peptide receptor C or guanylate cyclase C), which is an important anabolic regulator of endochondral bone growth [36,37]. The presence of these known genes in our gene lists supports the reliability of our results.
To gain more insights into the functional consequences from differential gene expression and to provide quantitative evidence, signature genes were subjected to the Gene Ontology (GO) database search [38] to find statistically overrepresented functional groups within gene lists. The DAVID 2008 web-based tool, a graph theory evidence-based method to agglomerate gene/protein identifiers [24], was applied for this task. A P value was assigned for each category based on the EASE cooccurrence scoring system, a Fisher's exact-based method that identifies functional categories overrepresented in a gene list [24]. Given that the whole human transcriptome was represented in a microarray, this analysis was not biased toward the coverage of the microarray. We addressed whether there were genes enriched in the differentiation categories according to the GO database and found BM-MSCs significantly enriched with genes involved in skeletal development and osteoblast differentiation (P = 0.00012; Fig. 2B). Fourteen BM-MSC genes (ALX1, ANKH, DLX5, ENPP1, FOXC1, EST1, EST2, NPR3, POSTN, RUNX2, SIX1, SOX9, SRGN, and TOB1) were present in this category (Fig. 2B).
Conserved biological properties, but not gene expression patterns, in MSCs cultured in different conditions
The above gene profiling data suggest differential biological potentials of WJ- and BM-MSCs cultured in the defined MesenCult medium. As culture conditions may affect cell properties dramatically and many laboratories culture their MSCs in DMEM supplemented with 10% FBS, we tested whether different culture media may alter the biological potentials of MSCs. BM- and WJ-MSCs cultured in DMEM+ 10% FBS for less than 6 passages were collected, and their gene expression patterns were again analyzed. When the whole transcriptome of each cell type was applied for MDS relationship analysis, we found that different culture conditions caused only minor changes in the gene expression patterns of WJ-MSCs (Fig. 2C). In contrast, BM-MSCs were more sensitive to environment cues as their molecular patterns were significantly altered by varied cellular conditions (Fig. 2C).
We then explored the possible functional consequences of altered gene expression patterns. A total of 984 probe sets were relatively abundant in WJ-MSCs cultured in 10% FBS, whereas another 808 probe sets were found in BM-MSCs (q < 0.001). These probe sets were again subjected to the GO database analysis to find statistically enriched functional groups. Genes involved in skeletal development were still predominant in BM-MSCs (P = 0.00026; Fig. 2D). DLX5, FOXC1, NPR3 and SIX1 were present in BM-MSCs cultured in both conditions (Fig. 2B, D, underlined).
The overexpression of osteogenic genes in BM-MSCs indicates they may possess a more similar transcriptome pattern to that of osteoblast. We then included osteoblasts, adipocytes, and chondrocytes array data in the MDS analysis. BM-MSCs cultured in either MesenCult medium (Fig. 2E) or DMEM + 10% FBS (not shown) showed a closer relationship to osteoblasts. To provide more quantitative evidences, we calculated the distances between osteoblasts and different MSC subtypes by employing an average linkage distance analysis to assess the similarity between 2 groups of gene expression profiles as described previously [21]. The distance between osteoblasts and BM-MSCs was smaller than that between osteoblasts and WJ-MSCs (Fig. 2F), supporting a superior osteogenic potential of BM-MSCs.
Pluripotent marker analysis on perinatal and adult MSCs
UC-MSCs were shown to have higher proliferative and colony-forming unit-fibroblast-forming ability than BM-MSCs in 2-dimentional monolayer cultures, and perinatal MSCs were suggested to be more primitive than adult ones [39]. To analyze whether this hypothesis could be reflected on a genomic level, we compared WJ- and BM-MSC transcriptomes with that of ESC. As shown in Fig. 3A, WJ-MSCs and ESCs shared a closer relationship, and such impression could be verified by average linkage distance analysis (Fig. 3B). Unsupervised hierarchical clustering also grouped WJ-MSCs and ESCs in the same leaf, while BM-MSCs and all 3 other mesenchymal cells in another (Fig. 3C).
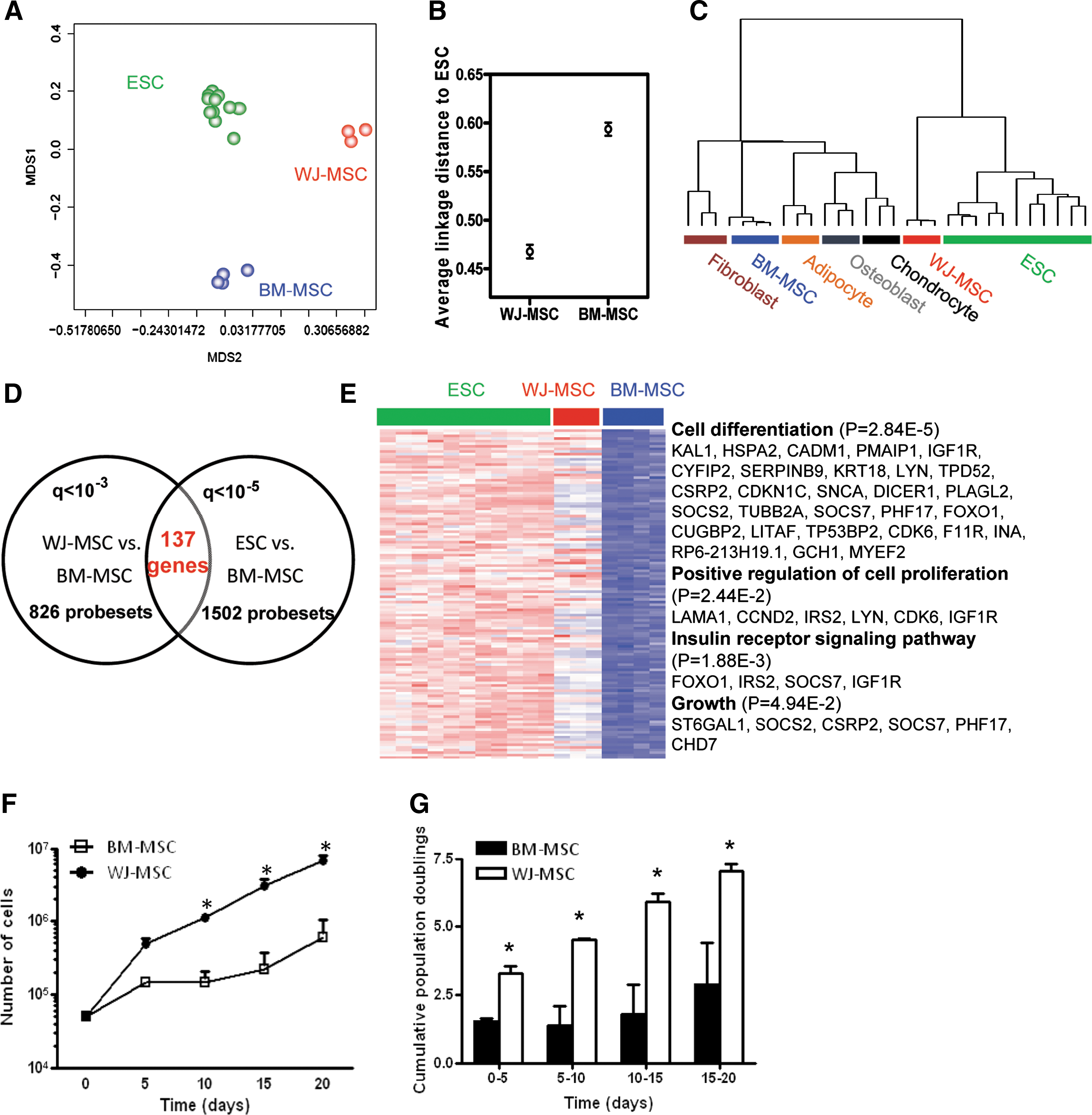
Comparative analyses of BM-MSCs, WJ-MSCs, and ESCs. (
We then investigated genes shared between ESCs and WJ-MSCs and checked the possible outcomes of the preferential ESC gene pattern in WJ-MSCs. A total of 137 genes were found commonly in both cell types when compared with BM-MSCs (Fig. 3D). Enriched among those 137 genes were those involved in cell differentiation (P = 0.0000284), positive regulation of cell proliferation (P = 0.0244), insulin receptor signaling pathway (P = 0.00188), and growth (P = 0.0494) (Fig. 3E). Also enriched in these 137 genes are those involved in antiapoptosis (FOXO1, SOCS2, IGF1R, SNCA, and SERPINB9; P = 0.029), cell adhesion (KAL1, PTPRF, LAMA1, CXADR, JUP, CADM1, DDR1, CYFIP2, DSC2, DSG2, CDK6, PCDH7, MLLT4, and F11R; P = 0.0024), and striated muscle development (CUGBP2, CXADR, CSRP2, and MYEF2; P = 0.032) (not shown).
The above analysis suggested a more multipotent status and preferential growth advantage of WJ-MSCs over BM-MSCs. We examined this hypothesis by culturing cells for 15 days. Cell number was counted every 5 days, and WJ-MSCs were found to proliferate faster than BM-MSCs (Fig. 3F). After day 5, WJ-MSCs increased their rate of growth relative to BM-MSCs, which corresponded to a significant increase in cumulative population doublings (Fig. 3G). Throughout the assay period, both cells types maintained similar cell morphology (ie, polygonal and fibroblastoid phenotypes) (data not shown).
Activated canonical pathways in different MSCs
It is likely that MSCs of different origins express different genes by utilizing unique canonical pathways. We then employed an additional tool, the IPA software, for investigating the biological pathways that may be differentially activated between 2 MSCs. The database behind IPA contains more canonical pathways culled from the literature. A 1-tailed Fisher's exact test was applied in IPA to determine the probability if the pathway assigned to the dataset is explained by chance alone (see Materials and Methods section). The canonical pathways associated to each dataset are ranked according to the significance of that pathway to the queried dataset.
Supplementary Fig. S1A, B (available online at
In BM-MSCs, more genes were involved in immune response, such as antigen presentation pathway (the top 1 enriched pathway) and chemokine, IL4/IL9/IL22, and GM-CSF signaling pathways (Supplementary Fig. S1B). Antigen presentation genes, including HLA-DMA, HLA-DRB4, HLA-DRA, HLA-DRB1, CD74, and HLA-DPA1, were abundant in BM-MSCs (Table 1B). Ephrin receptor and BMP signaling pathways were also active (Supplementary Fig. S1B). In both WJ- and BM-MSCs, the ILK signaling pathway was active (Supplementary Fig. S1).
Genetic networks of different MSCs
Increasing evidence showed that genes do not act as individuals but collaborates in genetic networks. To better understand how genes enriched in different stem cells are related and, most importantly, how those interactions are related to cellular function, we performed genetic network analysis for signature genes. Filtrated probe sets were inputted into the IPA software to construct network modules. The knowledge base behind IPA summarized known molecular interactions evidenced in published literature. The term “network” in IPA is not the same as a biological or canonical pathway with a distinct function (ie, angiogenesis), but rather is a reflection of all interactions of a given protein as defined in the literature.
In WJ-MSC a major network consisting 137 genes was identified (Fig. 4A). This network includes several growth factors or cytokines: CLEC11A, CXCL2, CXCL5, EGF, HBEGF, NGFB, and NTF3. Among those genes, EGF and HBEGF can enhance the migration of human and rabbit MSCs [40], and EGF has been also reported to stimulate the differentiation of human MSCs into bone-forming cells [41]. This network also revealed genes with significant biological roles in WJ-MSCs. Some genes have higher connectivity to others (regarded as “hubs”) or resided in a position among submodules in the major network (“bridge” genes) (Fig. 4A, panels 1–5). Deregulation of hub or bridge genes may eventually lead to the disruption of the genetic network and the malfunction of cells [42]. ICAM1, NFKB1 (NFκB), NGFB (nerve growth factor, beta polypeptide), and EGF are bridges connecting different submodules in the major network component (Fig. 4A, panels 1–4). Central to the network there are significant hubs, including ACTG2, CDK6, CCND2, CDKN1A, COL18A1, FOXO1A, FOXO3A, IGFBP5, KSR1, MYH9, NOS3, NFKBIA, PRKCD, and SERPINE1 (Fig. 4A, panel 5). Of these hub genes, CDK6, CCND2, COL18A1, IGFBP5, and SERPINE1 are all involved in enhancing cell growth. Two forkhead transcription factors (FOXO1A and FOXO3A) involved in brain development [43] are also present.
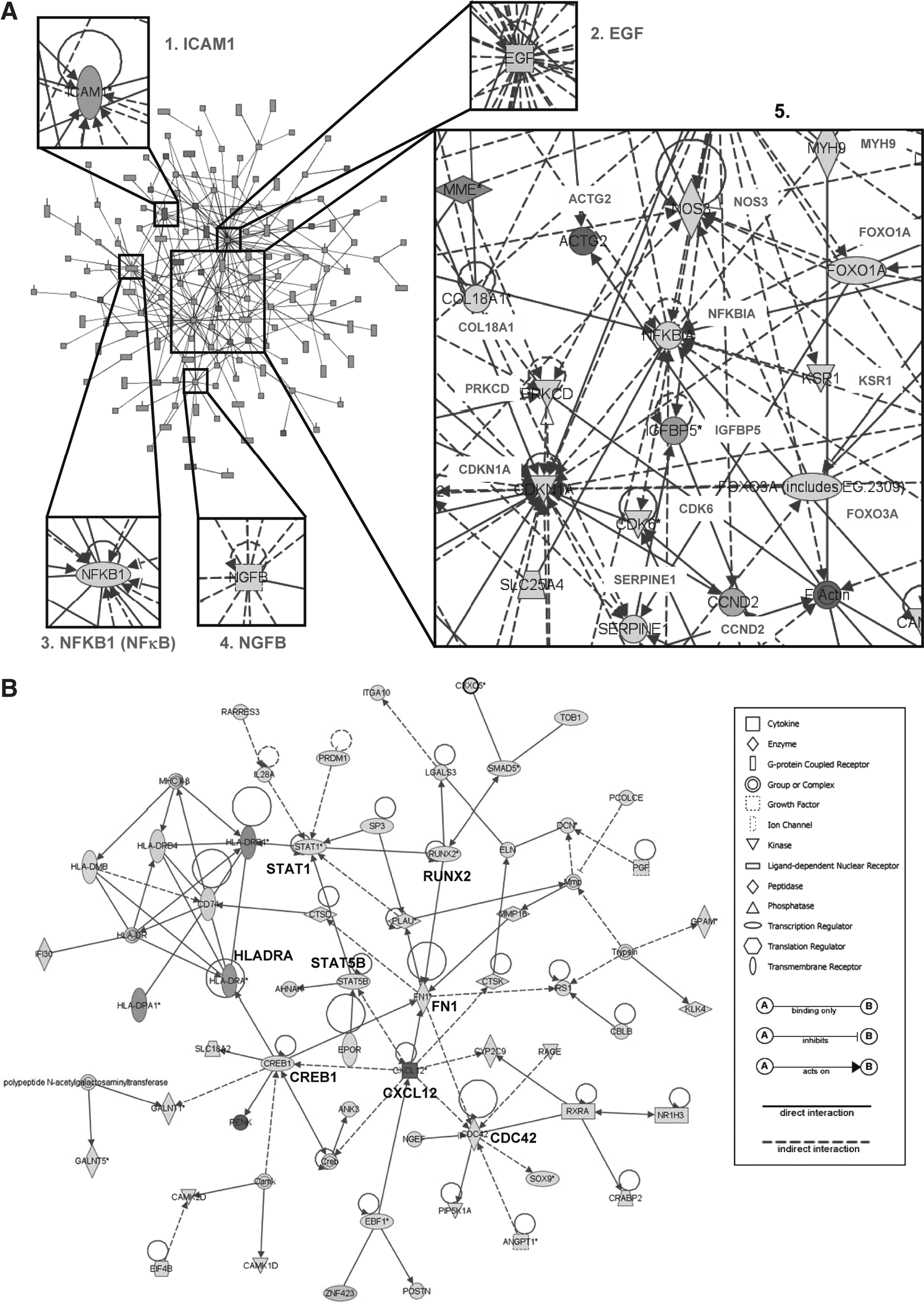
Interaction network analysis as a framework for the interpretation of MSC biology. (
When a similar functional network analysis was conducted on BM-MSC genes, a major network consisting 57 genes was identified (Fig. 4B). Extracellular proteins ANGPT1, CXCL12, DCN, ELN, IL28A, KLK4, LGALS3, MMP16, PCOLCE, PENK, PGF, PLAU, and POSTN are present in this network. HLADRA, CREB1, and RUNX2 link submodules to the main network (Fig. 4B). Also crucial in this network (ie, crucial in BM-MSCs) are transcription factors STAT1, STAT5B, CDC42, and cytokine CXCL12 (Fig. 4B).
Verification of in silico analysis results
The differential expression of genes involved in these functional groups was examined by qRT-PCR and immunoblotting in Fig. 5. Cell proliferation genes such as FLT1 and EGF were more abundant in WJ-MSCs (Fig. 5A). The dominant expression of TEK, a PI3K-AKT pathway gene, in WJ-MSCs could be verified by qPCR (Fig. 5B). As NFκB is one of the major downstream effectors of PI3K signaling, we examined by immunoblotting the levels of NFκB and IκB-α proteins in different MSCs. Clearly, WJ-MSCs expressed less IκB-α but possessed more active form p65 NFκB (phosphorylated at S536) (Fig. 5B). Genes involved in neurogenesis, such as RTN1, NPPB, and NRP2, were also more abundant in WJ-MSCs (Fig. 5C).
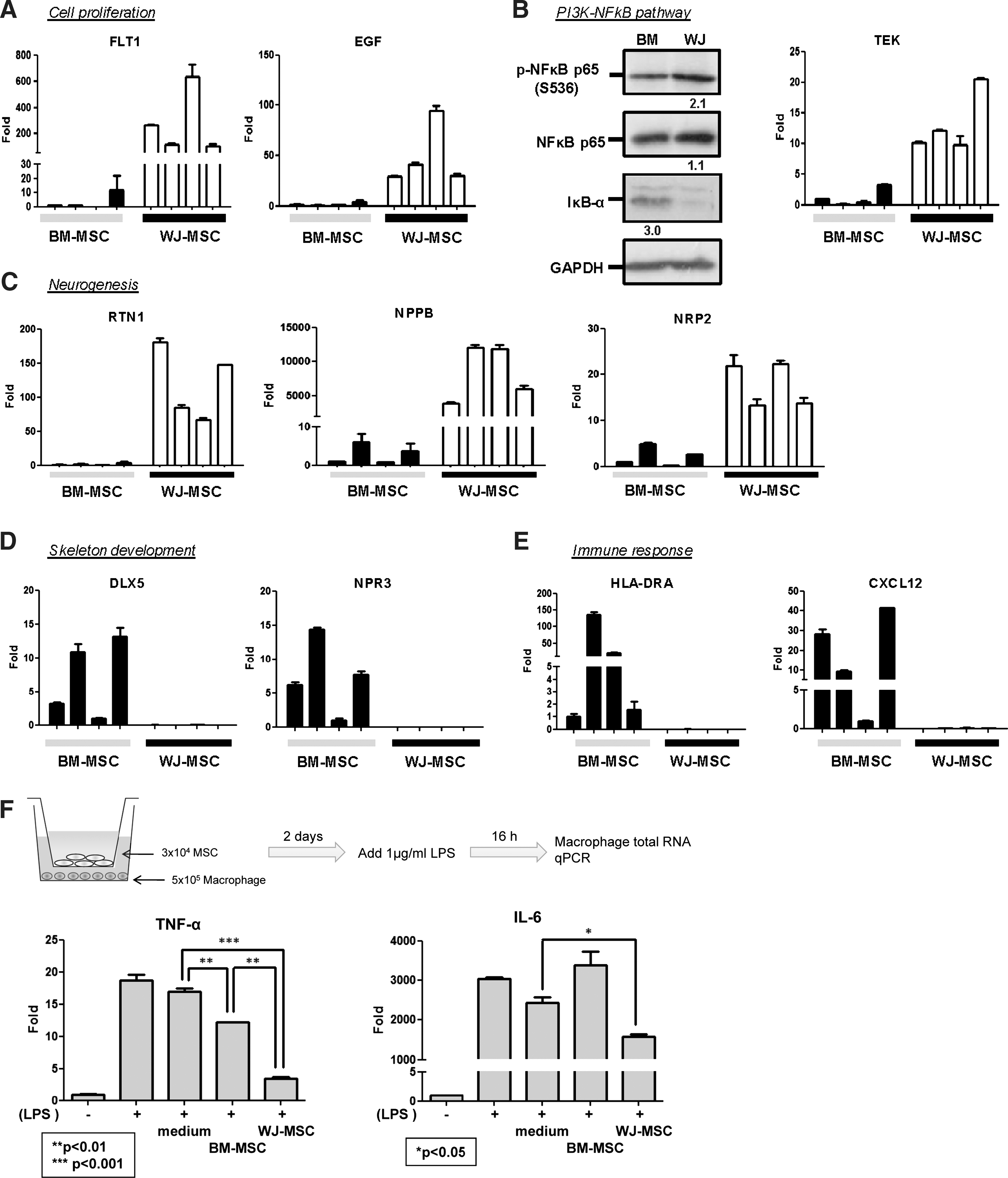
Validation of array data by qRT-PCR (
In BM-MSCs, genes involved in skeletal development were more abundant (Fig. 5D), consistent with drylab analysis data (Fig. 2). Also abundant in BM-MSCs were genes involved in immune response, such as CXCL12 and HLADRA (Fig. 5E), indicating WJ-MSCs maybe a better choice in allogeneic transplantation. The superior allogeneic transplantation potential of WJ-MSCs was verified by examining their immunosuppressive effects over allogenic macrophages. WJ-MSCs prevent cocultured macrophages from producing inflammatory cytokines tumor necrosis factor-α and IL6 more significantly (Fig. 5F).
Osteogenic differentiation potentials of WJ- and BM-MSCs
Genomics analyses suggested different osteogenesis characters of different MSCs as well as the involved genes. We examined drylab data by wetlab experiments. The osteogenic differentiation potentials of both MSCs cultured in either defined medium or DMEM + 10% FBS were tested by culturing the cells in osteogenic medium. Under osteogenic induction, BM-MSCs cultured in the defined MesenCult medium expressed higher alkaline phosphatase (ALP) activity than WJ-MSCs (Fig. 6A, B). ALP was expressed in more than 95% of BM-MSCs at week 3 but in only few WJ-MSCs even at week 4 (Fig. 6B). Mineralization of differentiated cells was further tested by von Kossa staining, which showed a greater extent of mineralization with more detectable bone nodules in differentiated BM-MSCs (Fig. 6C). Bone nodule could be observed clearly at week 3 in differentiating BM-MSCs but could not be observed even at week 4 in differentiated WJ-MSCs (Fig. 6C). The expression of 2 other osteogenic markers [osteopontin (SPP1) and RUNX2] was examined by qPCR. As shown in Fig. 6D, these marker genes were more abundantly expressed in BM-MSCs than in WJ-MSCs early during osteogenic differentiation. Similar results were found when MSCs were cultured in 10% FBS: ALP, osteopontin, and RUNX2 were expressed more dominantly in differentiated BM-MSCs (Fig. 6E). ALP staining data from osteogenic differentiated MSCs also showed that when MSCs were cultured in 10% FBS, BM-MSCs still held a better osteogenic ability (Supplementary Fig. S2, available online at
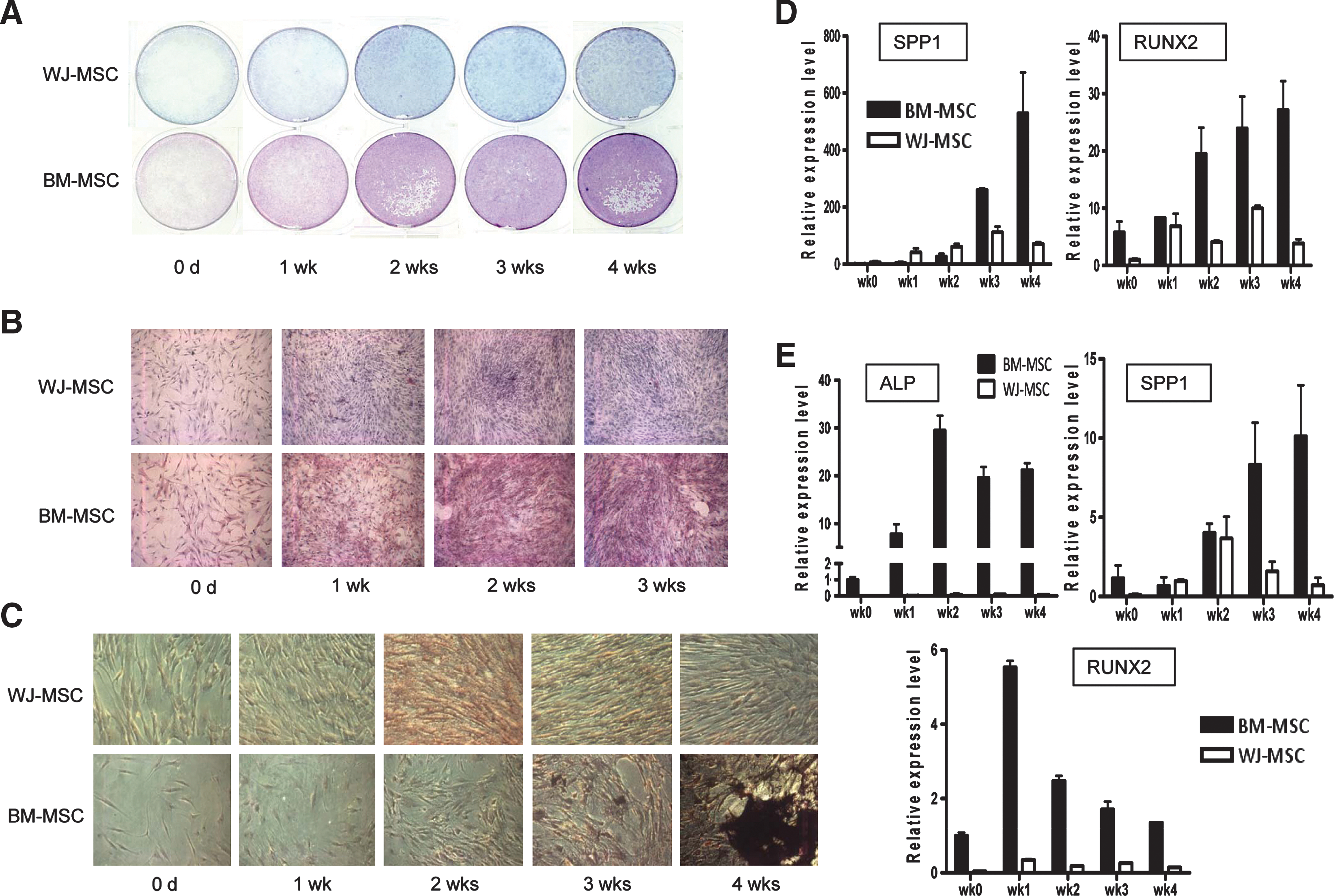
Different osteogenic capacities of WJ-MSCs and BM-MSCs. (
Discussion
Stem cells have drawn immense research interest because they hold great promise for future therapeutic applications. Although much progress has been made in stem cell biology, the clinical application of these cells is stymied by an inadequate or insufficient understanding of the functional integration of complex interactive genetic processes. Further characterization of these stem cells via a genomics approach will be critical for a better understanding of the molecular events involved in stem cell properties as well as the utilization of these cells. The differential gene expression patterns was indicated by immunophenotype analysis that unlike BM-MSCs, small populations of WJ-MSCs express endoglin (SH2, CD105) and CD49e at passage 8 [10]. Here we identify global transcriptome differences and functional variations between WJ- and BM-MSCs, and some of the functions revealed may correlate to clinical applications. We confirm drylab data by wetlab experiments and found that MSCs from BM are a better source for osteogenesis, whereas WJ-MSCs are more primitive and more similar to ESCs. These findings shed light on the myth of different MSCs, provide the underlying mechanisms, and will contribute in cell-based therapy.
The gene expression profiles, as well as their comparisons, of various human MSCs have been reported by quite a few groups [3,4,14 –18]. Nevertheless, interpreting genomics data into biological significance is always an overwhelming task. Analyzing gene expression data by assessing the biological significance of filtrated gene sets, rather than individual genes, has become a main approach in microarray data analysis and this has promisingly derived new biological interpretations of microarray data. In principle, this approach measures the gene–phenotype correlations of every gene in a predefined gene set, usually according to the GO [38] categories or the sets of genes representing biological pathways in the cell, and then give a score to represent its changes associated with the phenotype. The DAVID database we used and the Gene set enrichment analysis [44], for example, are among the most mentioned and used gene set-based approaches. In addition to dividing filtrated genes into functional subgroups, systems biology approaches (such as connecting signature genes into networks) also reveal genetic modules inside cells. In systems biology the overall molecular events are viewed as a biological system for a given biological process, where we can find important hub and betweenness controller genes, which could not be readily extracted from 1-dimensional gene list analysis [21,45,46]. By applying these 2 approaches (functional grouping and genetic network construction), we successfully generated hypotheses from bioinformatics analyses. For example, we suggested that BM-MSCs may hold a better osteogenic potential, whereas WJ-MSCs may proliferate quicker. These hypotheses could be verified by the following wetlab experiments (Figs. 3F, G and 6).
The functional comparison between WJ- and BM-MSCs has been performed by several groups [12,13]. However, the underlying mechanisms and responsible genes are yet unclear. The greater proliferative potential of WJ-MSCs may be due to the abundant expression of telomerase [5] or embryonic stem cell (hES) genes such as NANOG, DNMT3B, and GABRB3 [19]. Additionally, genes (including GAB2, NFKBIA, FOXO1A, YWHAB, FOXO3A, CDKN1A, MAP3K8, NFKB1, NOS3, DDR1 and TEK) involved in the phosphoinositide 3-kinase (PI3K)-AKT survival/proliferation pathway are also higher in WJ-MSCs (Supplementary Fig. S1, Figs. 4A and 5A, B). NFκB is also more active in WJ-MSCs (Fig. 5B), which partly explains why WJ-MSCs divide faster than those from human BM (Fig. 3F, G). One of several effector arms of PI3K-AKT signaling are the highly related members of the mammalian FoxO transcription factor family, FoxO1, FoxO3, and FoxO4 [47]. FOXO1 (FOXO1A) and FOXO3A are both key hub genes maintaining the stability of the whole genetic network in WJ-MSCs (Fig. 4A). RUNX2, a transcription factor crucial in osteogenesis, is a hub gene in BM-MSCs (Fig. 4B), suggesting a dominant osteogenic ability of BM-MSCs. We could confirm this by wetlab experiments (Fig. 6). Our transcriptome analysis on these 2 MSC subtypes therefore provides a basic roadmap for future in-depth exploration of the molecular mechanisms governing multilineage differentiation and stemness of MSCs.
On top of providing molecular mechanisms for reported phenotypes, our functional genomics analyses further provide new hypotheses. For example, WJ-MSCs might induce less rejection response in allogeneic transplantation because they express less antigen presentation genes (eg, HLA-DRA; Supplementary Fig. S1 and Fig. 5E) and suppress more host inflammatory cytokines (Fig. 5F). HLA-DRA is a member of MHC class II and is involved in immune recognition. Lower expression of HLA-DRA in WJ-MSCs may decrease the risk of transplantation rejection. Another hypothesis is that WJ-MSCs, although still can, may possess less osteogenic ability than BM-MSCs. We proved this point by wetlab osteogenic differentiation experiments (Fig. 6). A very recent study suggests that WJ-MSCs predominantly expresses genes engaged in osteogenic differentiation, and the tendency of WJ-MSCs to differentiate into osteoblast is similar to BM-MSCs [48]. However, a side-by-side functional comparison between WJ-MSCs and BM-MSCs was not performed. It was therefore unclear whether WJ-MSCs shares the same, or less, osteogenic potential than BM-MSCs. Our study thus provides the first parallel mesenchymal lineage differentiation analyses for these 2 MSC subtypes. WJ-MSCs and BM-MSCs have been compared for their cartilage tissue engineering potential in a 3-dimentional culture scaffold: type II collagen staining was more intense for hBM-MSCs than hUC-MSCs, suggesting a worse chondrogenic differentiation potential of WJ-MSCs [49]. As to adipogenic differentiation, Karahuseyinoglu et al. compared the adipogenesis abilities of WJ-MSCs and BM-MSCs and found that WJ-MSC-derived adipocytes were less mature compared with those from BM-MSCs [50]. This conclusion is consistent with our observations (Fig. 1D, E). WJ-MSCs may, thus, constitute a promising option for angiogenic or neural cells, whereas BM-MSCs in bone remodeling in regenerative medicine.
Several reports seem to report contradicting results that MSCs purified from human umbilical cord exhibited better osteogenic potential than BM-MSCs [39,51]. However, the MSCs they purified were actually from the perivascular portion of the umbilical cord, but not the WJ part. We used WJ-MSCs in comparative genomics research, and data obtained therefore are different. Several reports have shown that, except from the matrix part of WJ, MSCs or somatic stem cells with pluripotent differentiation potentials can also be isolated from the perivascular portion of the umbilical cord [52], or from the vasculature endothelial surface of the umbilical cord [53,54] (reviewed by Troyer et al. [55]). It has been shown that MSCs from WJ possess the least ability of osteogenesis when compared with those from umbilical veins or arteries [20]. Moreover, in the absence of osteogenic induction, MSCs from arteries also expressed osteocytic marker alkaline phosphatase [20]. Additionally, MSCs from WJ and perivascular regions have different marker expressions. For examples, pancytokeratin (an ecto/endodermal marker) and α-smooth muscle actin are only expressed in hUCPVCs, which are thought to be more differentiated myofibroblasts, whereas cells in WJ are more immature [50,56].
Using a knowledge-based database, only 29.1% of WJ-MSC genes and 32.6% of BM-MSC genes was found to be involved in networks, respectively. This is due to the fact that we applied a knowledge-based strategy to construct genetic networks, so that only genes with known interactions will be included. Stemness and differentiation are most complex processes governed by a highly coordinated regulation of distinct genetic programs, and biological function of genes enriched in different MSCs remains mostly unknown. The genetic networks revealed are therefore still incomplete. More comprehensive, integrated studies enabling the determination of all interactions will offer additional insights into how such a complex interaction map may contribute to unique stem cell behaviors. The global genetic networks among stemness genes are therefore needed to be further constructed via bioinformatics algorithms or data-driven approaches [21].
An improved understanding of MSCs of different sources can help the applications of these cells and the control of their differentiation directions in vivo, thereby furthering their potentials in clinical applications. Studying genes or pathways differentially expressed in various MSCs may eventually decipher the mechanisms of neurogenesis, neuroplasticity, angiogenesis, and osteogenesis. Thus, this study provides novel strategies for further functional or mechanistic studies on stem cells. The approaches taken here can also serve as a model for the comparison of other complex biological systems, or the study of the function of a given gene.
Footnotes
Acknowledgments
The authors thank Dr. Szu-Ting Chen from Prof. Shie-Liang Hsieh's laboratory for providing macrophages, and Dr. Tom K. Kuo for his critical reading of the manuscript. The authors acknowledge the technical services provided by the Microarray and Gene Expression Analysis Core Facility of the National Yang-Ming University VGH Genome Research Center. The Gene Expression Analysis Core Facility was supported by the National Research Program for Genomic Medicine, National Science Council (NSC). This work was supported by the NSC (NSC97-3111-B-010-004 and NSC98-2320-B-010-020-MY3), the Yen Tjing Lin Medical Foundation (CI-96-11), the National Health Research Institutes (NHRI-EX97-9704BI), the Mackay Memorial Hospital (MMH-HB-98-04), Department of Health (Cancer Excellence Center Plan), and a grant from the Yang-Ming University (Ministry of Education, Aim for the Top University Plan).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.