
Abstract
Neural progenitor cells reside in many regions of the adult brain. However, their capacity to generate new neurons relies on stem cell niches, consisting of stem cells, niche support cells, and basal lamina, which maintain stem cells and direct their differentiation and migration into tissue structures. Neurospheres are thought to expose neural progenitor cells to an environment reminiscent of the stem cell niche. We show that embryonic day 14.5 ventral mesencephalon neurospheres grafted into the midbrain of 6-hydroxydopamine lesioned mice express markers of mesenchymal cells, such as CD29 and CD44, and enclose a core of host-derived proliferating cells that express nestin, polysialic acid–neural cell adhesion molecule, βIII-tubulin, and neuron-specific nuclear protein. Laminin was expressed between the grafted cells and the core of proliferating host-derived cells. Further, infusion of the anti-mitotic agent β-
Introduction
T
Studies have reported that 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced lesions of the rodent midbrain are followed by NPC proliferation and acquisition of a dopaminergic (DA) phenotype [9,12], but at substantially lower rates of neurogenesis than in the SVZ and SGZ [9,13]. Other laboratories have been unable to reproduce these findings [11,14,15]. Notwithstanding this debate, Lie et al. described a population of actively dividing progenitor cells in the adult SN which, in situ, gave rise to new mature glial cells [8]. However, in vitro, and when transplanted into the hippocampus, these proliferating cells differentiated into neurons [8]. Therefore, while NPCs are present in the region of the adult SN, they lack appropriate environmental signals to give rise to new DA neurons. Transplantation studies provide evidence that the microenvironment, or stem cell niche, not only regulates the maintenance of stem cells but also has a potent influence on the neurogenic potential of NSCs. A stem cell niche is identified by the presence of stem cells, niche support cells, and a basal lamina [16,17]. In the SVZ and SGZ stem cell niches, cells with molecular and structural characteristics of astrocytes function as stem cells [18]. However, not all astrocyte-like cells within the neurogenic niche act as stem cells. Some act as niche support cells, sensing and regulating the niche's microenvironment as well as secreting factors that stimulate neurogenesis [19]. Depletion of NSC using the anti-mitotic compound cytosine β-
In the presence of mitogens, NSC/NPC can be propagated as neurospheres [23] and evidence is emerging that whole neurospheres provide a supportive environment that is similar to a stem cell niche [24 –28]. Neurospheres produce proteins required for their proliferation and survival, including fibroblast growth factor-2 (FGF2), platelet-derived growth factor-AA, insulin-like growth factor, and neurotrophin-3, together with their receptors [29,30]. Further, expression of nestin, Musashi-1, and Sox2 in neurospheres deprived of FGF2 suggests that the stem cell niche is maintained in the core of the spheres [31,32]. Whole neurospheres transplanted into normal and injured rodent brains survive better than dissociated neurospheres [33,34] possibly due to the close physical contact of cells within the sphere creating an effective trophic environment.
Trophic factors released by neurospheres may also support proliferation and differentiation of endogenous NPCs in nonneurogenic areas. Transplantation of neurospheres derived from the ventral mesencephalon (VM) of embryonic day (E) 14.5 embryos into the midbrain of severely SN lesioned mice promoted the generation of host-derived tyrosine hydroxylase (TH+) cells in the SN [24]. TH+ cells were also found in the vicinity of the graft and ectopic to the SN, suggesting that the graft had induced endogenous cells to acquire a TH phenotype. The increase in TH+ cells in the SN of grafted mice was accompanied by a significant increase in dopamine transporter (DAT+) terminals in the striatum of the grafted side and resulting functional improvement [24]. Regardless of whether this increase in host TH+ cells is due to neurogenesis or a phenotypic shift, this study demonstrated the capability of intact neurospheres to provide a supportive environment for an endogenous response to recovery after injury. More strikingly, the graft's organization resembled a stem cell niche. The grafts expressed glial markers, including glial fibrillary acidic protein (GFAP), suggesting that the graft cells could be behaving as niche astrocytes similar to those found in the SVZ and SGZ. Given the niche-like properties of a neurosphere, we wanted to determine whether the transplantation of VM neurospheres generated an in vivo niche capable of inducing endogenous TH+ cells. Therefore, the aim of this study was to closely examine the niche-like properties of the graft.
Materials and Methods
Experimental animals
All methods conformed to the Australian National Health and Medical Research Council published code of practice for the use of animal research and were approved by the Howard Florey Institute Animal Ethics Committee. Adult male C57BL/6J mice received transplants of neurospheres prepared from transgenic mice with C57BL/6J genetic background and an enhanced green fluorescent protein (EGFP) transgene under the control of a chicken β-actin promoter and a cytomegalovirus enhancer (The Jackson Laboratory).
Generation of neurospheres
VM neurospheres were generated and propagated as previously described [24,34]. Neurospheres were derived from E14.5 embryos of C57BL/6J female mice time-mated with heterozygous transgenic GFP male mice. The VM was dissected from embryos carrying the transgene and enzymatically dissociated using serum-free conditions [24,34]. The cells were subsequently seeded at a density of 1 × 106 cells/mL in neurosphere media containing epidermal growth factor (10 ng/mL) and FGF2 (10 ng/mL; Chemicon) in uncoated T25 flasks (Falcon). Neurospheres were supplemented with a fresh medium and mitogens every 48 h. Spheres were propagated by enzymatic dissociation every 7–10 days. Neurospheres used for transplantation were passaged >5 times.
Lateral ganglionic eminence (LGE) neurospheres were generated from E12.5 GFP embryos. The generation and propagation of LGE neurospheres was carried out according to the generation and propagation of VM neurospheres.
In vitro differentiation of VM neurospheres and immunocytochemistry
VM neurospheres, expressing GFP, were differentiated on poly-
Nigral lesions and amphetamine-induced behavioral testing
Animals were anaesthetized using 4% chloral hydrate and lesions of the right SN were made in 12-week-old C57BL/6J mice by stereotaxic injection of 10 μg of the DA neurotoxin, 6-hydroxydopamine (6-OHDA; Sigma) in 0.02% ascorbic acid (anteroposterior, 3.0 mm; lateral, 1.5 mm; dorsoventral, 4.3 mm, with respect to bregma). Three weeks later, the rate of D-met-amphetamine (5 mg/kg intraperitoneally; Sigma)-induced rotation was used to select animals with large lesions. Mice making >600 ipsilateral rotations in a 90-min period were chosen for transplantation.
Transplantation of whole neurospheres
Three weeks after the 6-OHDA lesion, mice were transplanted with 8–10 GFP+ VM (or LGE) neurospheres supranigrally [24]. Neurospheres isolated after 7 days in culture were resuspended in mitogen-free neurosphere media. Neurospheres (200–250 μm diameter) were drawn up into the sterile glass transplantation cannula attached to a 100 μL Hamilton syringe. Mice were anesthetized and placed in a stereotaxic frame. A cannula loaded with neurospheres was inserted into the midbrain, just dorsal to the substantia nigra pars compacta (SNpc) using the same craniotomy and coordinates for the lesion.
Transplantation of PA6 stromal cells
Mouse PA6 stromal cells were obtained from the Riken Cell Bank and cultured in α-MEM supplemented with 10% fetal bovine serum, 2 mM glutamine, 50 U/mL penicillin, and 50 μg/mL streptomycin (all purchased from Invitrogen). Cells were allowed to reach ∼70%–80% confluency at which point mitomycin-C (10 μg/mL) was added to the media for 3 h to halt cellular proliferation. The cells were subsequently washed in phosphate-buffered saline, trypsinized, and resuspended at 100,000 cells/μL. Lesioned mice were anesthetized and stereotaxically injected with 2 μL of cell suspension (200,000 cells) supranigrally (using the same coordinates as lesion) using a 26-gauge Hamilton syringe.
Ara-C infusions and bromo-deoxy-uridine injections
Miniosmotic pumps coupled to brain infusion cannulae (Alzet) were used to administer Ara-C (2% w/v in saline; Sigma) or saline. The cannulae were stereologically implanted into the midbrain at coordinates anteroposterior 3.0 mm, lateral 0.5 mm, and dorsoventral 4.3 mm, relative to bregma. Infusion was maintained for 7 days at a flow rate of 0.5 μL/h, after which time the mice were briefly anesthetized with 2% isoflurane to remove the osmotic pumps. Mice were killed by transcardial perfusion at the time of removing the osmotic pumps (day 0) or at 2, 5, and 25 days. Bromo-deoxy-uridine (BrdU; dose 50 mg/kg) was injected intraperitoneally every 8 h during the 24 h preceding death.
Immunohistochemistry
Animals were killed at 4, 8, 12, or 20 weeks after transplantation by an overdose of sodium pentobarbitone (100 mg/kg; Rhone Merieux) and transcardially perfused as described in Parish et al. [35]. The brains were then removed and equilibrated with 30% sucrose solution at 4°C for 48 h. A 1 in 6 series of 25-μm-thick coronal sections were cut through the SN and caudate-putamen (CPu). Sections were processed for goat anti-GFP (1:500; Abcam), rabbit anti-GFAP (1:800), rabbit anti-TH (1:500), mouse anti-NeuN (1:200), mouse anti-βIII-tubulin (1:600), mouse anti-nestin (1:200) (all from Chemicon), mouse anti-5A5 [polysialic acid–neural cell adhesion molecule (PSA-NCAM), 1:200; Developmental Studies Hybridoma Bank], rabbit anti-laminin (1:100), rat anti-BrdU (1:400), rat anti-CD29 (1:50), rat anti-CD44 (1:50), rabbit anti-doublecortin (1:100), and CD11b (1:100) (all from Sigma). Sections were incubated with the respective primary antibody overnight at room temperature followed by incubation with the respective AlexaFluor secondary antibody (1:200) and observed by fluorescence or confocal microscopy. Chromogenic staining for TH was also carried out for counting. Sections were incubated with a rabbit polyclonal anti-TH antibody (1:1,000) overnight at room temperature followed by biotinylated secondary antibody and avidin–biotin peroxidase complex. Staining was developed by cobalt- and nickel-intensified diaminobenzidine reaction and counterstained with 1% neutral red.
Quantitative analysis
The number of TH+ neurons in the midbrain of mice was counted. The extensiveness of the lesions prevented accurate delineation of the SN boundaries. Hence, counting was restricted to sections of the SN containing the third nerve radical, a region that could be clearly defined in all animals. DA cells within the midbrain were identified by TH immunoreactivity and their neuronal morphology. Cells along the midline and ventral tegmental area (VTA) were omitted when counting the total number of TH+ cells in the midbrain. Cells that were considered ectopic midbrain TH+ cells if they lay outside of the SN, VTA, and midline regions. Therefore, the number of ectopic cells was obtained by subtracting the number of TH+ cells in SN region from the total TH+ cells counted. It should be noted that sections containing the third nerve represented ∼60% of the entire SN; the method used therefore may under estimate the extent of the host DA cell response. Since TH+ cells in the core of the graft lacked neuronal morphology, they were not counted.
For mice treated with Ara-C, the number of nestin, NCAM, and βIII-tubulin+ cells within each graft niche was counted. The number of nestin, PSA-NCAM, and βIII-tubulin+ cells 25 days after Ara-C was counted from every sixth section. Estimated numbers of nestin, PSA-NCAM, and βIII-tubulin+ cells in each animal were calculated by multiplying the number of cells counted by 6.
Statistical analysis
A t-test was used to compare the difference in TH+ cells in grafted and sham graft animals by means of a statistic software package (Sigma Stat version 2.03). One-way analysis of variance and Bonferroni t-test were used for all other data. Significance was set as P < 0.05.
Results
In vitro differentiation of VM neurospheres
The differentiation potential of the VM-derived NSC was determined in vitro. The trilineage potential of the VM-derived NSC was confirmed by the presence of astrocytes (GFAP), oligodendrocyte progenitor cells (NG2), and neurons (βIII-tubulin and NeuN). All differentiated neurospheres expressed GFAP, 69% of neurospheres expressed NG2, 35% expressed βIII-tubulin, and 22% expressed NeuN. The differentiation potential of VM-derived NSC has been further characterized by Moses et al. [24].
Location and morphology of the graft
Three weeks after 6-OHDA administration, the extent of DA cell loss was assessed by amphetamine-induced rotation. Subsequent histological examination confirmed that the right SN was almost completely depleted of TH immunoreactive cells in animals with high levels of rotation (Fig. 1A–C). Grafts of whole VM neurospheres derived from E14.5 GFP transgenic embryos were then implanted into the lesioned (right) mesencephalon of mice with extensive lesions, as assessed by rotation. The grafted cells, identified by GFP immunoreactivity, survived in all animals. These grafts commonly appeared as tear shape aggregates dorsal to the SN (Figs. 1B and 2). GFP+ cells were only found within these aggregates, or occasionally along the cannula tract, but were not observed in other regions.

Photomicrographs showing graft location and TH expression in midbrain.
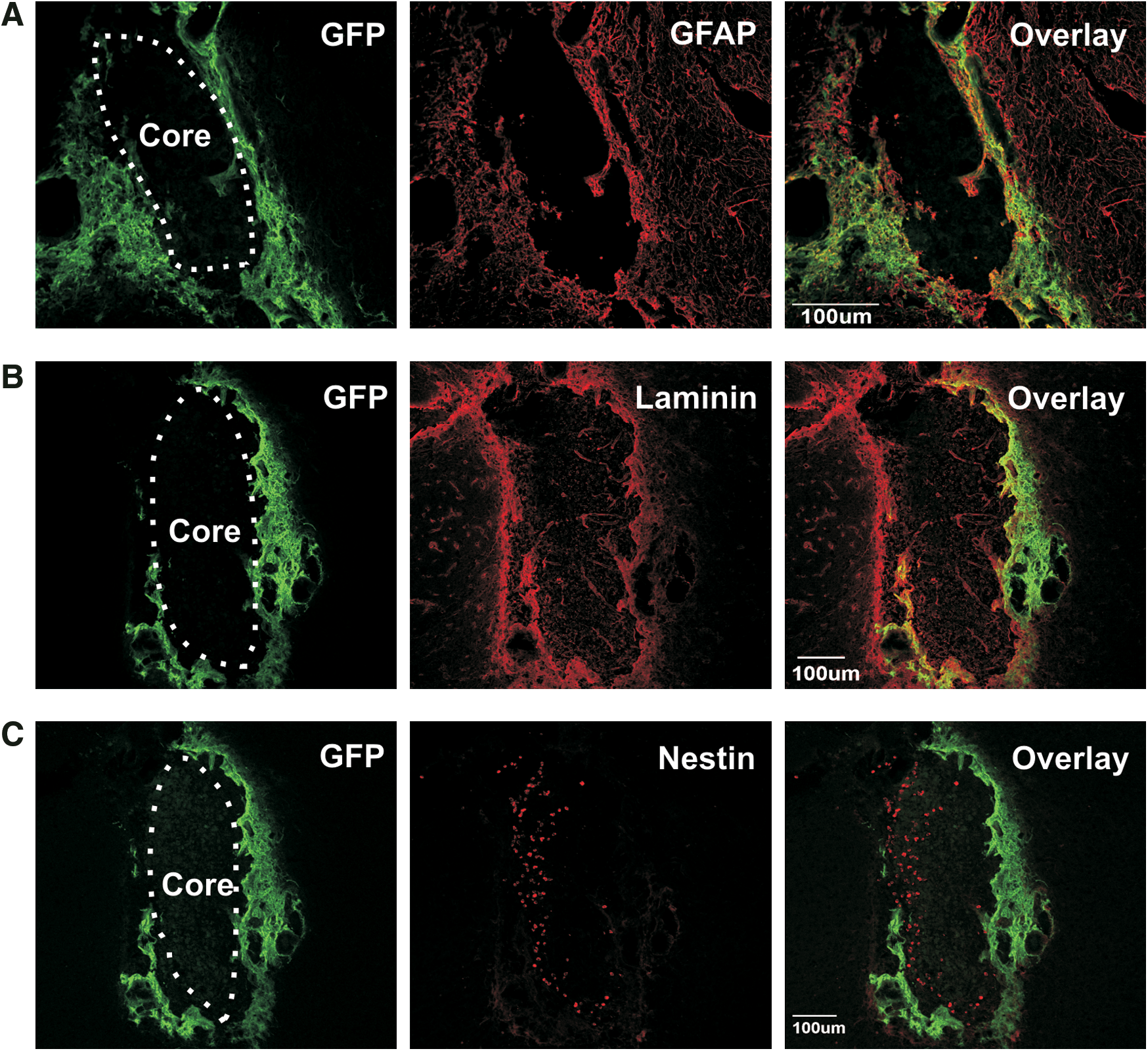
Confocal fluorescence photomicrographs showing graft morphology. GFP+ graft cells
Eight weeks after transplantation, grafts consisted of an outer shell of donor GFP+ cells (Fig. 2A–C) and an inner core of host-derived (GFP) cells (Fig. 2C). Cells in the core of the graft were small and round with a large nucleus, suggesting an immature phenotype (Fig. 2C). Similar to the basement membrane of the SVZ, a laminin layer separated graft cells from core cells (Fig. 2B) and graft cells closest to the core also expressed laminin (Fig. 2B).
Characterization of graft-derived GFP+cells
Most GFP+ graft cells were spindle shaped with long extensions, similar to fibroblasts and expressed mesenchymal stem cell markers, including CD29 and CD44. Each graft varied in the extent to which graft cells expressed these markers, with almost all GFP+ cells expressing these markers in some animals (Fig. 3Ai, Aii, Ci, and Cii), whereas few GFP+ cells coexpressed CD29 and CD44 in other animals (Fig. 3B, D). Immunoreactivity to CD29 and laminin also highlighted the rich vascularization of the grafts (Fig. 3Ai, arrow). Expression of CD29 and CD44 by graft-derived cells suggests characteristics similar to mesenchymal stem cell found in stem cell niches of the intestinal tract. Approximately 20% of graft cells also expressed GFAP (Figs. 2A and 4Ei, Eii), a common marker of the slowly dividing stem cells found within stem cell niches in the CNS. No graft cells coexpressed TH.

Confocal fluorescence photomicrographs showing expression of mesenchymal markers. GFP+ grafted cells expressed CD29
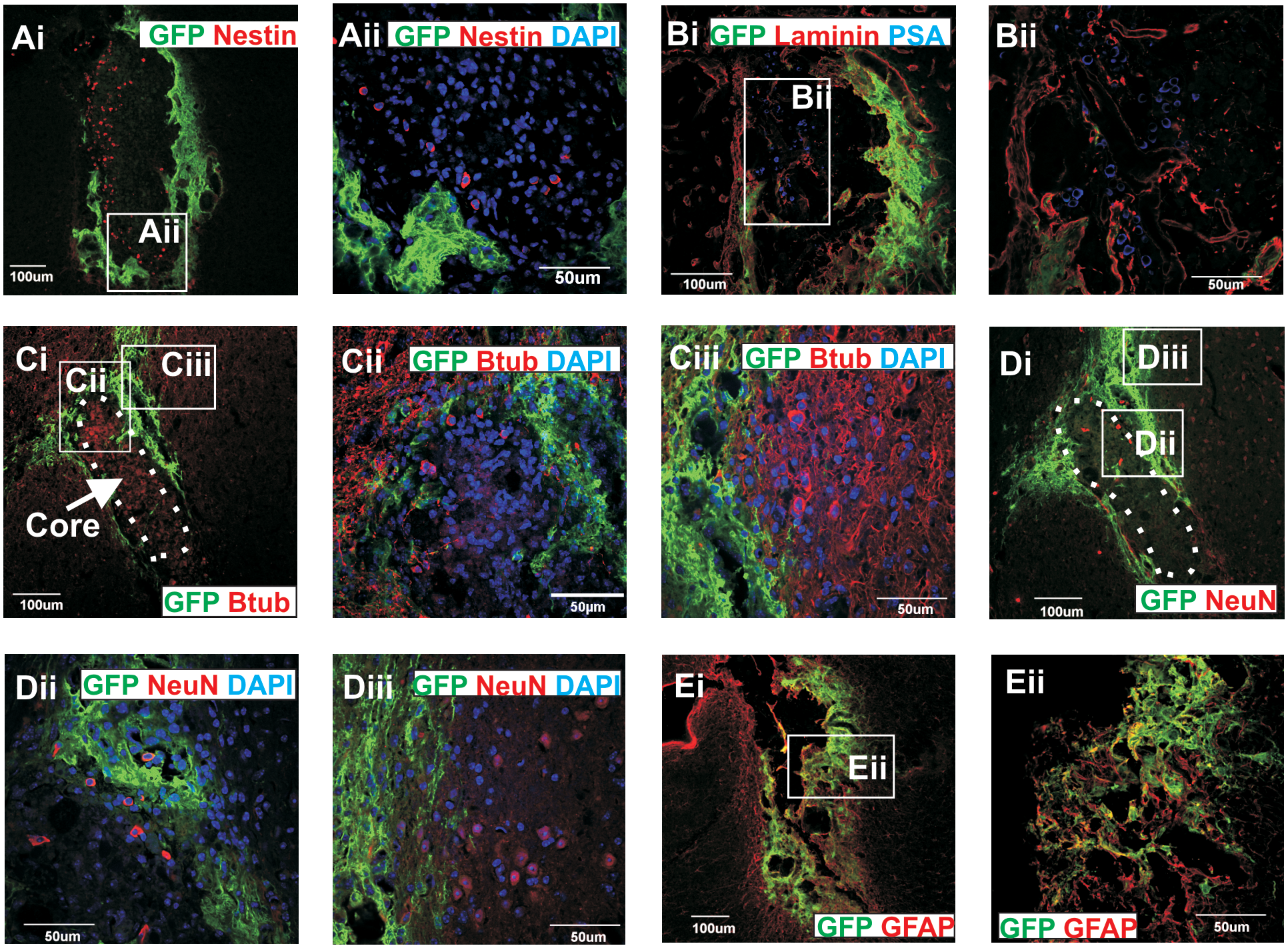
Confocal fluorescence photomicrographs showing phenotype of host-derived cells within core of the graft. The cells in the middle of the graft did not express GFP and were immunoreacted for
Characterization of the host-derived core cells
Several core cells expressed markers of NSC (nestin; Fig. 4Ai, Aii), immature neuronal-committed progenitor cells (PSA-NCAM; Fig. 4Bi, Bii), immature neurons (βIII-tubulin; Fig. 4Ci, Ciii), and mature neurons (NeuN; Fig. 4Di, Dii). More mature neuronal subtypes (βIII-tubulin+ and NeuN+) were also found at the external margins of the core (Fig. 4Ci, Cii, Di, Diii). Interestingly, at 4 and 8 weeks after grafting TH+ cells could be identified within the core (Fig. 5A, B) but did not coexpress DAT, Nurr1, GIRK2, doublecortin, or GFAP (data not shown). No TH+ cells could be identified in the core at 12 or 20 weeks after grafting (Fig. 5C, D, respectively). Some of the small, round GFP cells within the graft core were also immunoreactive for CD45, a hematopoietic cell marker (data not shown).
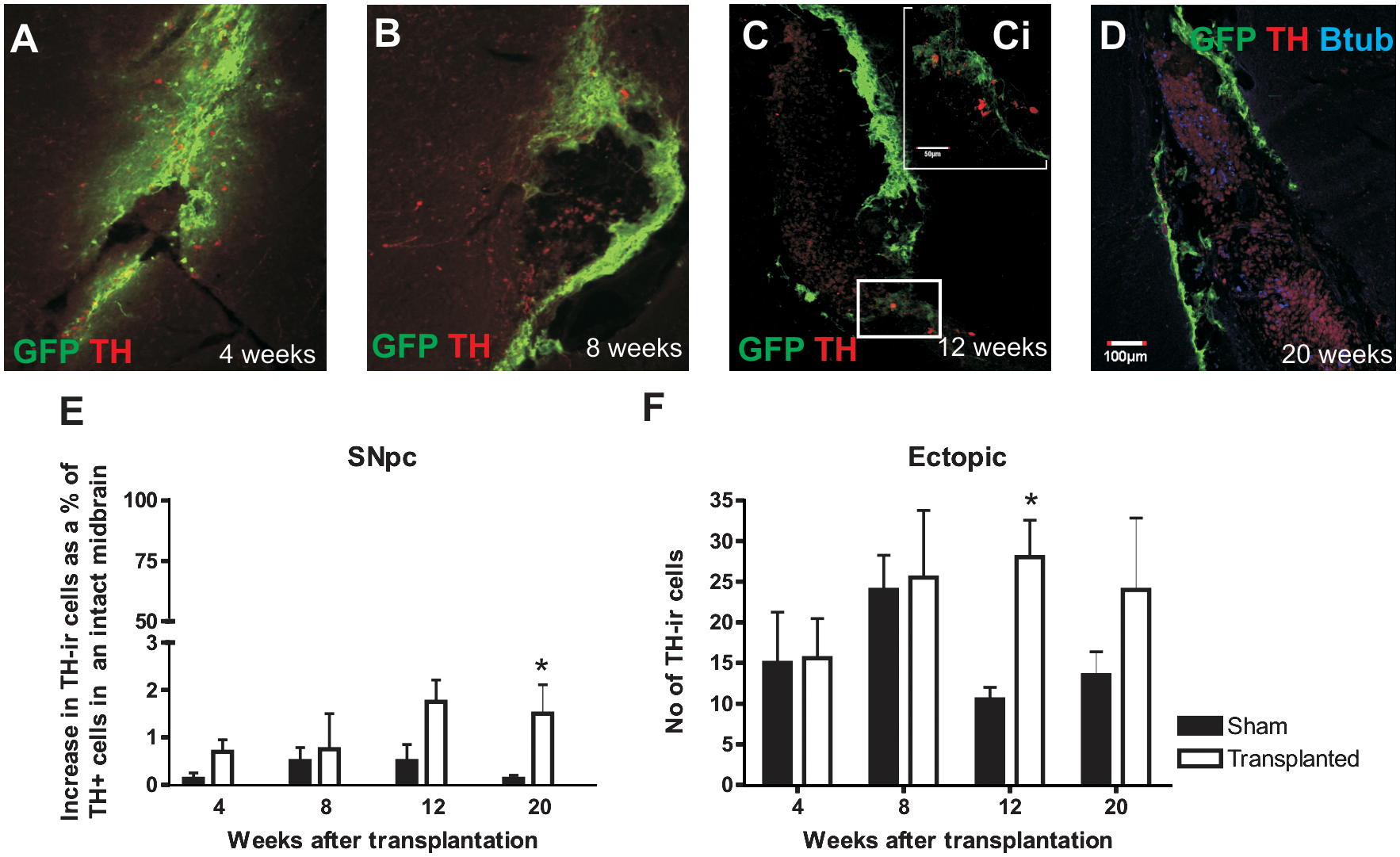
Fluorescence photomicrographs of the GFP+ graft cells and TH+ cells.
Time course of graft development and maturation
Grafts were examined at 4, 8, 12, and 20 weeks after transplantation. Four weeks after transplantation, the graft consisted of a sheet of GFP+ cells with unorganized laminin interlaced throughout. The central core was usually absent or rudimentary when present (Fig. 5A), and a few TH+ cells with neuronal morphology could be observed surrounding (and occasionally within) the grafts. By 8 weeks posttransplantation, the graft cells and laminin had formed into a tear shaped outer shell (Fig. 5B) surrounding round immature host-derived cells in the core of the graft (Fig. 5B). Figure 6 summarizes the components of the graft.

A cartoon representation of the components of the graft with grafted cells (green), laminin (orange), immature host-derived cells (purple), and more mature cells surrounding the graft (red). Color images available online at
Although sham grafting elicited a modest response, consisting of laminin, GFAP+ as well as CD45+, nestin+, and NCAM+ cells (data not shown), these cells were rare compared to the core of VM neurosphere grafts. Further, by 12 weeks, the number of CD45+, nestin+, and NCAM+ cells had diminished further. None of the small round cells in the sham grafts expressed TH at any time point. Transplantation of dissociated VM neurospheres resulted in a response similar to sham grafts (data not shown).
Effect of grafting on TH cell numbers in the midbrain
We also quantified the time course of TH expression by cells in the SN and ectopic to the SN. TH+ cell counts were expressed as a percentage of TH+ cells in the midbrain of SN sections containing the third nerve of normal mice (Fig. 5E). The numbers of TH+ cells within the SN of grafted animals increased sharply between 8 and 12 weeks and remained elevated at 20 weeks and significantly greater than sham-grafted animals (Fig. 5E). Although TH+ cells were absent from the core of 12- and 20-week-old grafts (Fig. 5C, D), ectopic TH+ cells steadily increased in VM-grafted animals until a plateau level was reached at 8 weeks posttransplantation, whereas ectopic cells declined over that period in sham-grafted animals (Fig. 5F). It is important to point out that our counting methods would underestimate SN and ectopic TH+ cells. Although the morphology of TH+ cells around the graft was neuronal (Fig. 1E, F), they did not coexpress NeuN and were smaller than classical SN TH+ cells (Fig. 1A). In addition, they did not express markers of SN DA neurons, including Nurr1, DAT, and GIRK2 (data not shown). These data illustrate that VM grafts induce greater increase in TH+ numbers in the midbrain of PD mice than do sham grafts.
Is there proliferation within the graft?
The presence of nestin+, PSA-NCAM+, and βIII-tubulin+ cells within the core of the graft led us to examine an analogy with proliferating cell types found in the SVZ, that is, rapidly dividing neuroblasts (type A cell), transit amplifying cells (type C cells), as well as the slowly dividing NSC (type B cell). The regenerative capacity of these cells was investigated by infusing the anti-mitotic agent Ara-C into the midbrain of mice transplanted 8 weeks earlier with VM neurospheres. In most animals, a 7-day infusion of Ara-C eliminated host-derived cells from the core of the graft (Fig. 7A–C, implying that these cells were proliferating cells. The numbers of PSA-NCAM+, nestin+, and βIII-tubulin+ cells and fibers were maintained in animals infused with saline.
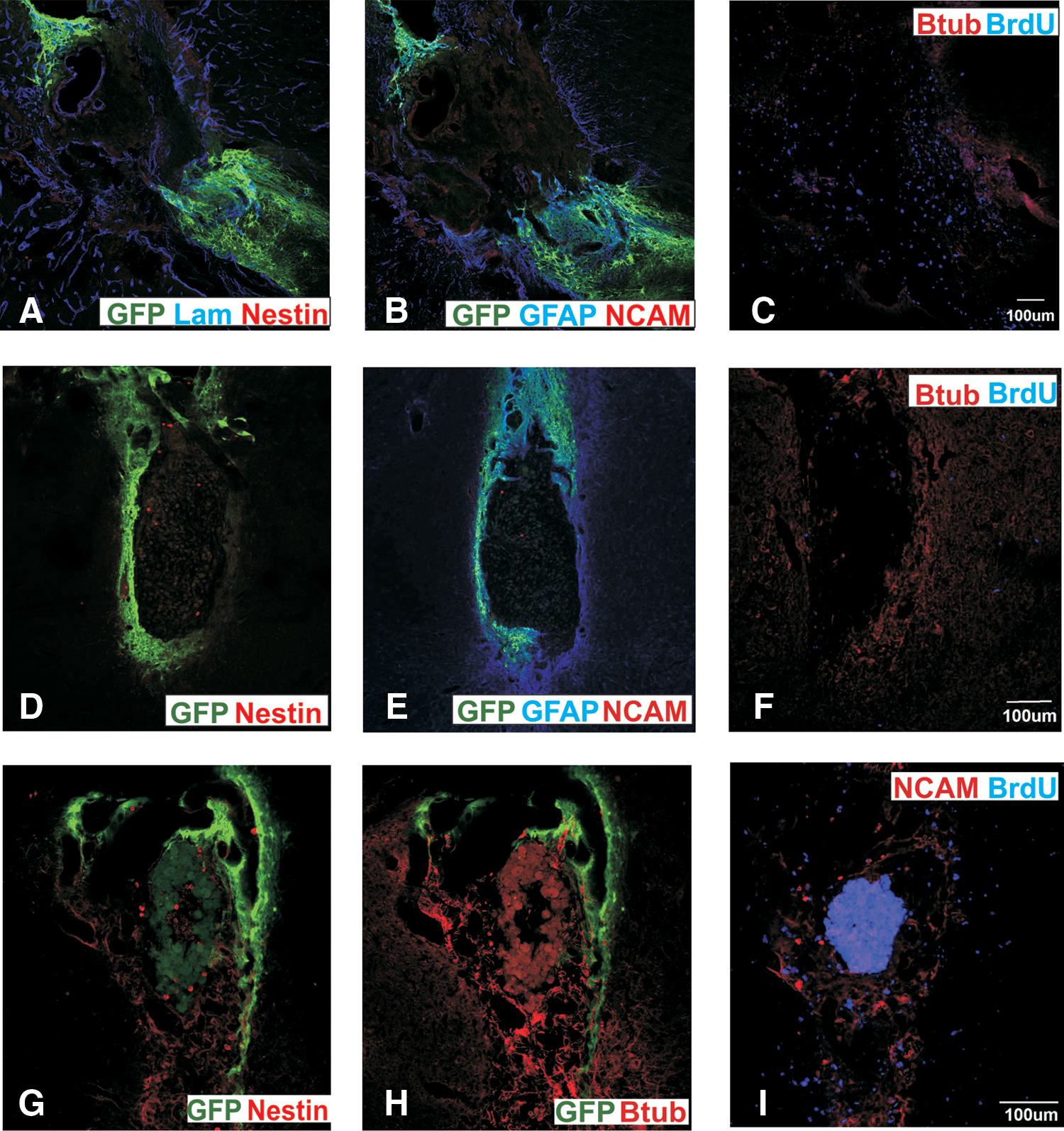
Fluorescence photomicrographs showing the effects of treatment with Ara-C. (
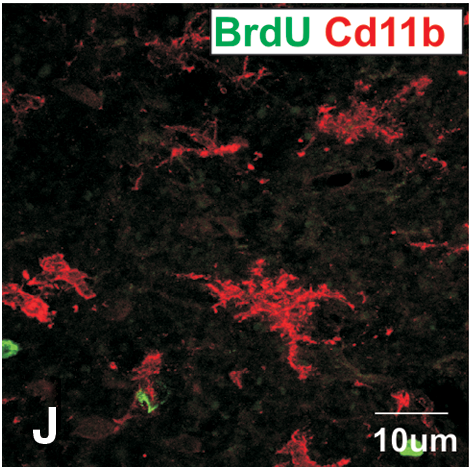
Subsequently, animals were killed at varying intervals after pump removal to assess the time course of regeneration. Nestin+, PSA-NCAM+, and βIII-tubulin+ cells were absent, 0–4 days after pump removal. By 25 days after pump removal, PSA-NCAM+ and nestin+ cells were present in most grafted animals; however, numbers were extremely low (averaging 108 nestin and 96 nestin+ cells/graft) and suggesting that the rate of cell division was extremely low (Fig. 7G–I). When the number of nestin, PSA-NCAM, and βIII-tubulin cells in the core of 25 day group was counted, there were an average of about 108 nestin, 96 PSA-NCAM, and 42 βIII-tubulin+ cells in an animal, indicating that the rate of cell division was very low. To determine when dividing cells reappeared, BrdU was injected every 8 h in the 24 h before killing the mice. There were many BrdU+ cells in and around the graft and cannula tract of animals killed immediately after pump removal (day 0) (Fig. 7C); however, these cells did not express PSA-NCAM, nestin, βIII-tubulin, CD11b, or GFAP. Five days after pump removal, BrdU+ cells around the graft were few and scattered (Fig. 7F); however, substantially more were present in animals killed 25 days after pump removal (Fig. 7I). No BrdU+ cells found at 5 and 25 days coexpressed any neural markers. It is probable and likely that the reappearance of new cells (PSA-NCAM and nestin) that did not costain for BrdU was due to the BrdU pulsing regime and the slow turnover of the proliferating cells.
Effect of grafting LGE-derived neurospheres
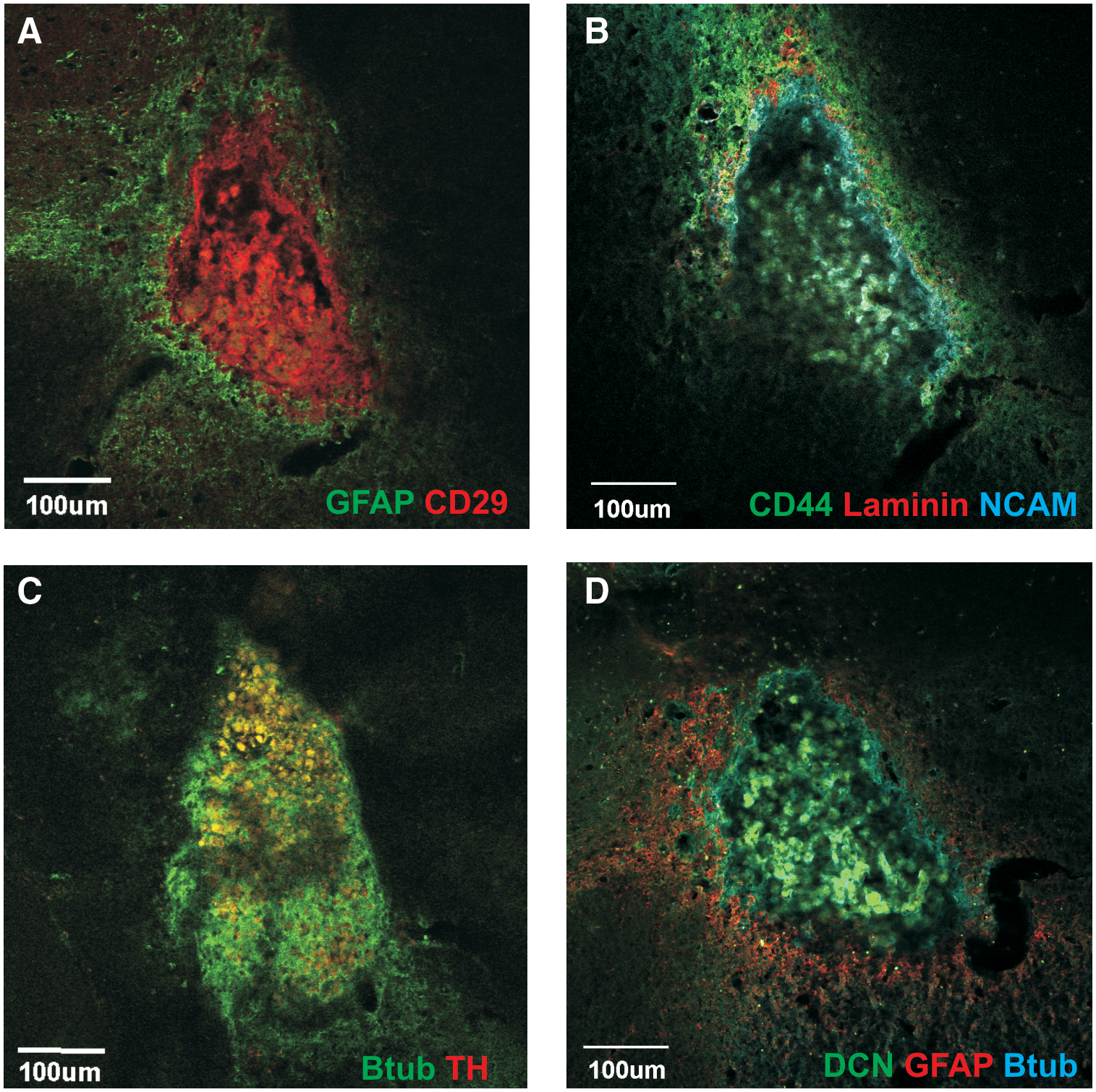
Transplantation of LGE-derived neurospheres into midbrain of mice with large 6-OHDA lesions similarly formed tear-shaped aggregates when transplanted into the dorsal mesencephalon. The graft cells expressed GFAP (Fig. 8E) and other stromal markers such CD29 and CD44. Host-derived cells in the core of the graft expressed TH (Fig. 8A; but did not colocalise with NeuN), NeuN (Fig. 8A, B), nestin (Fig. 8C, D), PSA-NCAM (Fig. 8E, F), and βIII-tubulin (Fig. 8G, H). Similar to VM grafts, laminin separated the graft cells from the host-derived cells in the core (Fig. 8C, D). These findings suggest that LGE-derived neurospheres could successfully generate a stem cell niche-like environment, similar to VM-derived neurospheres. Counts of TH+ cells showed the response in generating ectopic TH+ cells in the midbrain with LGE grafts was similar to that of VM grafts (Fig. 8I).

Effect of transplanted LGE-derived neurospheres into the 6-OHDA lesioned SNpc. GFP LGE grafts formed tear-shaped aggregates similar to VM grafts. Host-derived cells expressing
Effect of grafting PA6 cells
VM-grafted cell develop mesenchymal-like characteristics, which may be important in developing the features that were similar to a stem cell niche. This possibility was examined by transplanting PA6 cells, a bone-marrow-derived stromal cell line, into the midbrain of 6-OHDA lesioned mice. Mice were killed 12 weeks after transplantation and the graft identified by immunostaining for CD29 and CD44 (Fig. 9A, and B, respectively). GFAP+ cells were found lining CD29+ and CD44+ cells (Fig. 9A, D). The graft also consisted of PSA-NCAM+ cells (Fig. 9B) and βIII-tubulin+ cells and fibers (Fig. 9C, D). Expression of laminin around the graft was scant compared with VM neurosphere grafts (Fig. 9B). No TH+ cells were found within or around the graft (Fig. 9C). Hence, although transplantation of PA6cells produced grafts with some similarities to VM grafts, importantly they failed to induce TH+ cells.
Phenotype of PA6-grafted cells. The PA6 graft was identified by expression of CD29 (red;
Discussion
We present evidence that grafting intact E14.5 VM neurospheres into the midbrain of mice with large SN lesions results in a structure with features of a stem cell niche. An adult stem cell niche is a specific site (eg, gonadal, epithelial, and gut) where stem cells reside and undergo self-renewal and differentiation [36]. They share a relatively simple structure that entails stromal partner cells, attached to a basement membrane and adjacent proliferating precursors [16,17], which were also present in the structures described in this study. The morphology of the grafted cells was reminiscent of stromal cells and expressed appropriate markers (GFAP, CD29, and CD44) [37]. Proliferating cells were mostly found within the core and thus in a position to be anchored by laminin. Typical of a stem cell niche, blood vessels were abundant within the graft.
The host-derived cells at the core expressed nestin, PSA-NCAM, βIII-tubulin, or NeuN suggest but do not prove a proliferative capacity. In the SVZ, slowly dividing type B or stem cells are identified by GFAP, vimentin, and nestin immunoreactivity [38]. Type C cells (or transit amplifying cells) express nestin [38], which in turn, generate type A cells (or migratory neuroblasts) that are identified by expression of PSA-NCAM, nestin, and βIII-tubulin [20,38]. Thus cells at the core of the graft express markers consistent with type B, C, and A cells, and their morphology is also consistent with proliferating or immature cells. Further confirmation of their proliferative capacity was provided when they were eliminated by infusing Ara-C into the region of the graft. Rapidly dividing cells would be most susceptible to an antimitotic agent and the reappearance of cells in the core of the graft suggests that they may be present there. The rather indolent reappearance of the cells over 25 days was much slower than in the Ara-C-treated SVZ [20] and suggests very few dividing cells with a low rate of division. On average, 108 nestin+, 96 PSA-NCAM+, and 42 βIII-tubulin+ cells were found within the graft core 25 days after pump removal. These cells did not begin to appear until day 5, indicating that the 108 nestin+ cells were generated within 20 day. This slow cell division (∼5 nestin+, 5 PSA-NCAM+, and 2 βIII-tubulin+ cells/day) could explain the difficulty in colabeling with BrdU given that the bioavailability of BrdU after injection is only 3 h [39] and that we pulsed only 3 times in the last 24 h. This slow turnover of cells is consistent with other reports [8,12]. The capacity to regulate the size and number of cells is a further feature of a niche that sets it aside from either a tumor or a modified repair process. The time-course studies provide evidence that the number of cells in the core of the graft was stable over a long period (Supplementary Figure S1), arguing for regulation of the rate of proliferation within the core. The rather slow return of cells following Ara-C treatment suggests that some signaling mechanisms (such as Sonic hedgehog) required for regulating cell numbers may prevent the complete regeneration of the niche-like structure [40,41].
The origin of the proliferative cells within the graft core is a matter of speculation. They could either be resident midbrain cells [8] or have migrated into the site [42]. Although the question of whether the SN can be repopulated by endogenous host-derived stem cells [9,11] is debated, actively dividing NPC have been described in the adult SN, which, in vitro, differentiate into neurons [8]. These cells also form neurons when transplanted into the hippocampus, where neurogenesis readily occurs. However, when transplanted into the mesencephalon, they gave rise to glia, suggesting that, normally, the midbrain does not provide the appropriate signals to support neurogenesis. We therefore speculate that the graft-derived stromal-like cells provide these signals to support proliferation of these endogenous cells previously identified by Lie et al. [8]. It is therefore of interest in this context that sham grafts and dissociated neurospheres also caused an evanescent and modest increase in TH+ cells, even though there are no dorsolateral mesencephalic TH+ cells in normal animals.
Expression of CD45+ by core cells raises the possibility that cells of hematological origin have migrated into the midbrain as a result of trauma. The argument against this is the length of time after grafting for development of the graft structure and the appearance of CD45+ cells. Four weeks posttransplantation the grafts are predominantly a sheet of GFP+ cells with little or no laminin. It is not until 8 weeks that graft cells develop a more stromal-like appearance and abut a basal membrane consisting of laminin and together surround the core. The time required to organize into a mature niche argues strongly that the core cells are not merely a result of trauma. Trauma immediately recruits activated microglia and macrophages [43], peaking 7 days postinjury and often subsiding by 4 weeks [44]. Cavitation and glial scarring then ensue and the scar tends to reduces in size as the inflammatory infiltrates diminish and the region is repopulated with astrocytes [45,46]. In contrast to the inflammatory responses to trauma, the organization of the grafts took ∼8 weeks to form and remained stable for at least 20 weeks.
The presence of ectopic TH+ cells around the graft may suggest that some host cells produced within the core of the graft had subsequently migrated out and matured. TH+ cells surrounding the graft began increasing in number 4 weeks after transplantation and numbers continued to increase until 12 weeks (note that these TH+ cells have neuronal morphology and do not express neuronal markers). The disappearance of TH+ cells from the core by 12 weeks, when ectopic TH+ cells were continuing to increase, suggests migration of TH+ cells out from the core of the graft. However, none of the ectopic TH+ cells coexpressed BrdU (Supplementary Figure S2). This is not surprising in view of the very low rate of division of core cells. Properly organized niches have guidance pathways and trophic gradients (eg, transforming growth factor 3β) to chaperone the regulated exit of these cells. Presumably, these factors are lacking in the present environment; otherwise more immature cells may have escaped from the core.
It is also possible that ectopic TH cells and the increase in SNpc TH+ neurons result from phenotype shift [47] in cells in the midbrain surrounding the graft rather than for cell proliferation. Factors released by VM neurospheres appear to play an important role in the endogenous response [24] and also in maintaining the DA phenotype in vitro [34], and in this context it is relevant that LGE neurospheres transplanted into the midbrain also induced TH+ cells and generated a core, while PA6 cells transplantated into the lesioned midbrain failed to generate either TH+ cells or a significant core. These results suggest that neural signal provided by the neurospheres derived from the LGE and VM are important for the formation of a stem cell niche-like environment and for inducing TH+ cells. Adjustments in host nigral neurons through the release of neurotrophic factors are consistent with reports that transplantation of human NSC derived from the VM induce host-derived TH+ cells [48,49]. Moreover, transplantation of neurospheres rather than a cell suspension (like the PA6 cells that were transplanted) may also aide in increasing the plasticity of the brain, by providing an environment similar to a stem cell niche.
A previous study demonstrated that transplantation of intact E14.5 VM neurospheres into the midbrain of mice with large SN lesions resulted in partial repopulation of the SN with TH+ cells of host origin, accompanied by increased DAT terminal density in the dorsal striatum and subsequent functional recovery [24]. They also reported an increase in TH+ cells of host origin in the dorsal mesencephalon in the immediate vicinity of the graft. Although the SNpc TH+ cells all appear to have a normal neuronal phenotype, the ectopic TH+ cells were smaller than SN TH+ cells, and did not express markers of SN neurons such as DAT, Nurr1, or GIRK2.
In summary, we provide evidence that intact neurospheres grafted into the midbrain are capable of forming a niche reminiscent of the adult stem cell niche as seen in the SVZ and SGZ of the hippocampus. Although not all functional aspects of a stem cell niche are evident, our niche appears to be capable of providing trophic support in the neighboring region resulting in the generation of TH+ cells and will provide a good model to understand how more functional niches can be created. Further efforts to address the formation of a niche may provide avenues for exploiting the proliferative properties of the adult brain in repair.
Footnotes
Acknowledgment
Funding for this research was through the National Health and Medical Research Council (NHMRC) of Australia.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.