
Abstract
Allergic asthma is an inflammatory airway disease caused by T helper type 2 (Th2)-driven immune responses. Recent studies have demonstrated that adipose-derived stromal cells (ASC) have an immunosuppressive effect on T-cell activity. This study was performed to investigate whether ASC can inhibit Th2-dependent allergic airway inflammation in mice. BALB/c mice were sensitized to ovalbumin (OVA) by intraperitoneal injection. To investigate the effect of ASC on the development of asthma phenotypes, 2 × 106 ASC were injected intravenously before OVA challenge. We evaluated the airway hyperresponsiveness (AHR), the proportion of eosinophils and cytokine production in bronchoalveolar lavage fluid (BALF), airway inflammation, and the intracellular cytokine staining of T cells in the BALF and spleen. Airway hyperresponsiveness, airway eosinophilia, and mucus production were markedly reduced after ASC administration before OVA challenge. The increased interleukin (IL)-4, IL-5, and transforming growth factor (TGF)-β1 levels in the BALF after OVA challenge were significantly reduced by the administration of ASC. This inhibition was accompanied by decreased IL-4+ CD4+ T cells and increased interferon (IFN)-γ+ CD4+ T cells in the BALF and spleen. The results of this study suggest that ASC administration before an allergen challenge inhibits AHR, lung inflammation, and Th2 cytokine production induced by an allergen challenge through inhibition of Th2 cell activity.
Introduction
A
Mesenchymal stromal cells (MSC) are multipotent cells found in several adult tissues; they can be induced to differentiate into various components of the bone marrow (BM) microenvironment, such as bone, adipose, and stromal tissues [4]. Mesenchymal stem cells represent a nonhomogeneous population of multipotent cells with a limited true stem cell component [5]. Stem cells have been viewed as a potential source of cells that could be used for replacement purposes in any tissue due to their capacity to differentiate into virtually any type of cell. In addition to their differentiation capability, MSC have been recently demonstrated to suppress T-cell, B-cell, and dendritic cell activities, thus exerting an immunoregulatory influence both in vitro and in vivo [6 –10]. Several studies have demonstrated that MSC affect the function of immunocompetent cell types, lending support to the possibility that they could be used to treat autoimmune disorders [11,12]. Until recently, the BM has been the major source of human MSC; however, it is difficult to obtain sufficient quantities of MSC from BM. Subcutaneous adipose tissues are accessible, abundant, and a potential alternative source of MSC present in every individual. Adipose tissue represents a source of MSC that has the potential for clinical application in the repair of damaged tissues as well as immunoregulatory activity. Similar to the MSC from BM (BM-MSC), the adipose-derived stromal cells (ASC) can differentiate along multiple lineage pathways and have immunosuppressive properties [13,14]. These immunosuppressive effects might have particular impact on transplantation procedures; similar to the BM-MSC, the infusion of ASC might support transplant tolerance and decrease graft-versus-host disease (GVHD) in allogeneic BM transplantation [15 –17].
MSC can exert profound effects on immune responses, primarily through the inhibition of effector functions; this suggests the possibility of using MSC for the treatment of immune-mediated disorders including not only GVHD, but also autoimmune diseases and organ transplantation. The immunological outcome, resulting from the in vivo administration of MSC, is not currently completely predictable; environmental factors are likely, in part, responsible for shaping their functional properties. However, the immunological outcome resulting from the in vivo administration of ASC, in allergic asthma, has not been reported to date. Based on the data suggesting that ASC have immunosuppressive effects and that these effects are determined by the microenvironment, we investigated the inhibitory effects of ASC on the development of allergic airway inflammation in a mouse asthma model.
Materials and Methods
Animals
Female BALB/c mice, 8 weeks of age and free of murine-specific pathogens, were obtained from Hyochang Science (Daegu, Korea). The study protocol, for the animal experiments in the present study, was approved by the Institutional Animal Care and Use Committee (IACUC) of the Pusan National University School of Medicine.
ASC isolation and culture
Adipose tissue was obtained from the abdominal fat of the BALB/c mice. To isolate homogeneous ASC, the adipose tissue was washed extensively with equal volumes of phosphate-buffered saline (PBS), and digested with 0.075% collagenase type I (Sigma, St. Louis, MO) at 37°C for 30 min. Enzyme activity was neutralized with α-modified Eagle's medium (α-MEM), containing 10% fetal bovine serum (FBS) and centrifuged at 1,200g for 10 min to obtain a pellet. The pellet was filtered through a 100-μm mesh to remove cellular debris and incubated overnight at 37°C/5% CO2 in control medium (α-MEM, 10% PBS, 100 unit/mL of penicillin, 100 μg/mL of streptomycin). Following incubation, the plate was washed extensively with PBS to remove residual nonadherent red blood cells. The resulting cell population was maintained at 37°C/5% CO2 in control medium. After 1 week, when the monolayer of the adherent cells reached confluence, the cells were trypsinized (0.05% trypsin; Sigma) resuspended in α-MEM containing 10% FBS and subcultured at a concentration of 2,000 cells/cm2. The cultured ASC were prepared by incubation with Try–ethylenediaminetetraacetic acid (EDTA) in a 37°C incubator for 5 min. The cells were washed with PBS and incubated with Cell Tracker CM-Dil (Molecular Probes, Eugene, OR) 2 μM in a 37°C incubator for 5 min, and then for an additional 15 min at 4°C. The cells were washed with PBS and suspended in PBS at a concentration of 2 × 106 cells/mL. For the experiments, we used the third or fourth passage of ASC. Flow cytometric analysis was performed to characterize the phenotypes of the ASC. At least 50,000 cells (in 100 μL PBS/0.5% bovine serum albumin (BSA)/2 mmol/L EDTA) were incubated with fluorescein isothiocyanate (FITC)-labeled monoclonal antibodies against mouse CD105, CD90, CD45, and CD11b (BD Biosciences, San Jose, CA) or with the respective isotype control. After washing, the labeled cells were analyzed by flow cytometry using a FACSCalibur flow cytometer and Cell Quest Pro software (BD Biosciences).
Asthma induction and treatment protocols
To generate a mouse model of asthma, the mice were sensitized with 2 intraperitoneal injections of 25 μg of ovalbumin (OVA) (Sigma) emulsified in 1 mg of aluminum hydroxide (Pierce Chemical Co., Rockford, IL) in a total volume of 200 μL on Days 1 and 14, and then challenged with an aerosol of 5% OVA in PBS using an ultrasonic nebulizer (NE-U12, Omron, Japan) for 30 min per day on Days 21–23. To evaluate the inhibitory effects of ASC, 0.1 mL of purified stem cells (2 × 106 cells/mL) were injected via the tail vein once a day on Days 18–20. The injected cells were labeled with a cell Tracker CM-Dil (Molecular Probes). For the sham-treated mice, PBS was injected instead of the ASC in the same manner. To evaluate the effects of the ASC on the negative control mice, the ASC were administered via the tail vein in the PBS-sensitized mice (Fig. 1A).
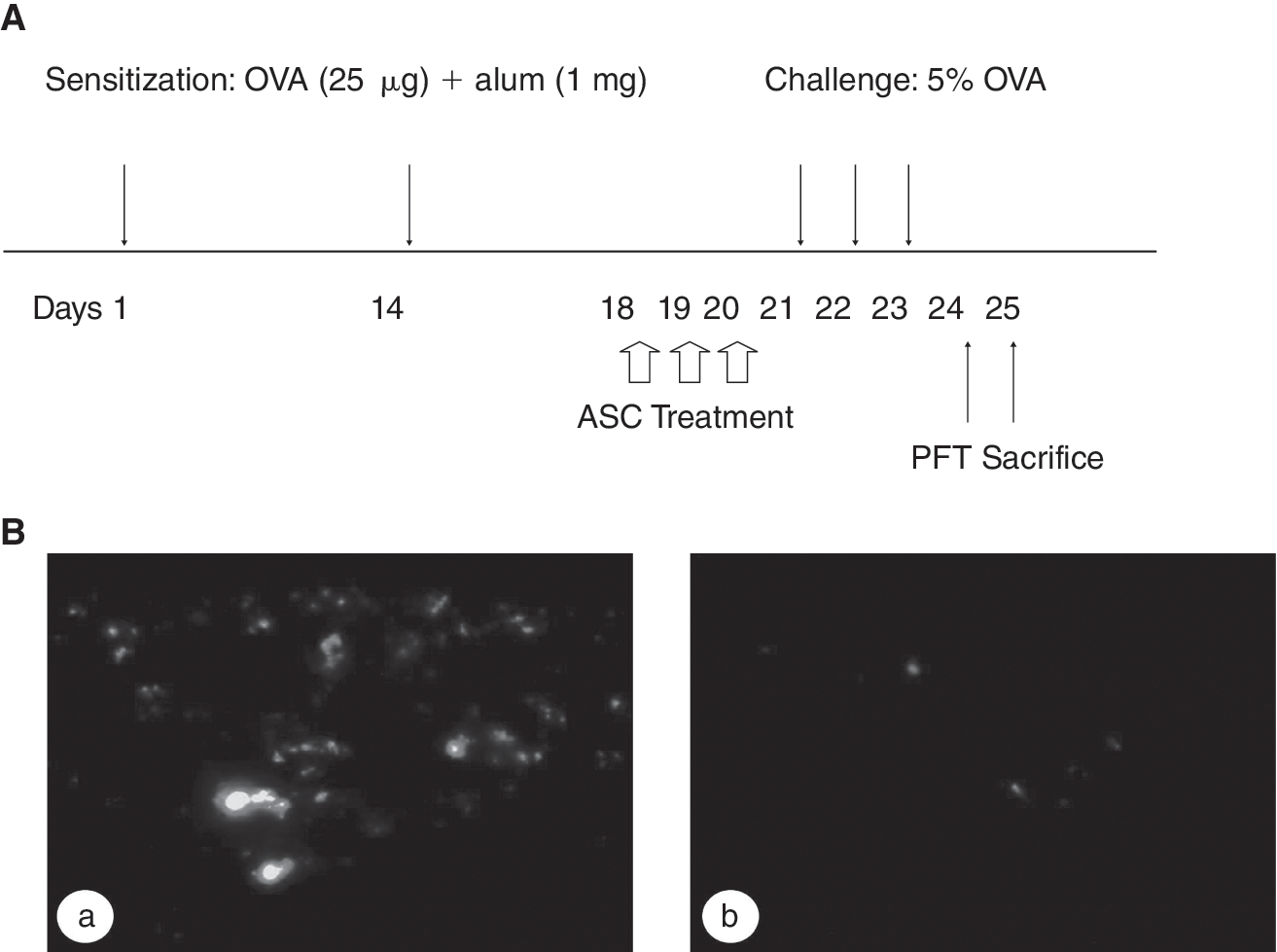
Study protocol (
Measurements of methacholine AHR
Twenty-four hours after the last challenge, the AHR was assessed in conscious, unrestrained mice using noninvasive whole-body plethysmography (Allmedicus, Seoul, Korea) as previously described [18]. In brief, the mice were placed in the plethysmography chamber and exposed to increasing concentrations of aerosolized methacholine at 6.25, 12.5, 25, and 50 mg/mL for 3 min. The enhanced pause (Penh) was calculated automatically based on the mean pressure generated in the plethysmography chamber during inspiration and expiration combined with the time of each phase. The Penh values calculated during each 3-min interval were then averaged.
Analysis of bronchoalveolar lavage fluids
Lung and bronchoalveolar lavage (BAL) samples were obtained as described previously [19]. The mice were anesthetized, and the trachea was isolated by blunt dissection. A small-caliber tube was inserted and secured in the airway. Two successive volumes of 0.75-mL PBS were instilled and aspirated gently, and then these 2 volumes were pooled. Each BAL sample was centrifuged, and the supernatants were stored at −70°C until use. The total numbers of inflammatory cells were counted after dilution of cell pellets with 50 μL of PBS. After Diff-quick staining (Dade Behring, Dudingen, Switzerland) of BAL pellets in a cytospin preparation, the types of inflammatory cells were determined by counting 300 cells, which were classified as macrophages, lymphocytes, neutrophils, or eosinophils. The levels of interleukin (IL)-4, IL-5, interferon (IFN)-γ, transforming growth factor (TGF)-β, and IL-10 in each supernatant were determined using commercial ELISA kits (R&D Systems, Minneapolis, MN).
Lung histology
For the histopathological assessment, the lungs were removed, fixed in 4% formalin overnight, and embedded in paraffin. Hematoxylin and eosin stain was used on the lung sections. The same microscopic magnification was used in all sample slides for comparison. Periodic acid–Schiff (PAS) staining was also performed to evaluate mucus secretion of the lung sections. Lung inflammation was assessed by the degree of peribronchial and perivascular inflammation, which were evaluated on a subjective scale of 0–3 as previously described [20]. In brief, a value of 0 was assigned when no inflammation was detectable, a value of 1 for occasional cuffing with inflammatory cells, a value of 2 when most bronchi or vessels were surrounded by a thin layer (1–5 cells) of inflammatory cells, and a value of 3 when most bronchi or vessels were surrounded by a thick layer (>5 cells deep) of inflammatory cells. Total lung inflammation was defined as the average of the peribronchial and perivascular inflammation scores. As 10–15 tissue sections per mouse were scored, inflammation scores were expressed as a mean value.
Intracellular cytokine staining
To stain intracellular IFN-γ and IL-4, BAL cells and splenocytes were cultured in plate-coated anti-CD3 (145.2C11, 10 μg/mL) for 12 h. The cultured cells were treated with brefeldin A (10 mg/mL; Sigma-Aldrich) for 4 h and washed with PBS. The cells were first stained for CD4 or CD8, subsequently fixed, and permeabilized using a Cytofix/Cytoperm Kit (BD Biosciences), according to the manufacturer's instructions, and incubated with PE- or FITC-conjugated anti-IFN-γ or FITC- or PE-conjugated anti-IL-4. Fluorescence was measured using a FACSCalibur (BD Biosciences), and data analysis was performed using CellQuest pro software (BD Biosciences).
Statistical analysis
All data were expressed as means ± SEMs. Comparisons of parameters among the groups were made by the Mann–Whitney or Kruskal–Wallis tests using SPSS software (SPSS 12.0, Chicago, IL). Statistical significance was defined as a P value of <0.05.
Results
Isolation of stable ASC
ASC was obtained after 3–4 cycles of culture from the adipose tissue of BALB/c healthy mice as described previously in our study [21]. The cultured ASC from adipose tissue was negative for CD45 and CD11b but positive for CD90 and CD105 on flow cytometry. The percentage of CD90+ cells in ASC was 54% and the percentage of CD105+ cells in ASC was 20.1%. The multipotency of the ASC isolated from mouse subcutaneous adipose tissue was demonstrated by incubating the cells in media that promoted differentiation into the osteogenic and adipogenic cell lineages (data not shown).
OVA challenge enhanced ASC migration into the lung
To demonstrate localization of the injected ASC to the lung, we identified the cells by labeling them with a fluorescent dye. The ASC labeled with a fluorescent dye were more intensely distributed within the lung in the OVA-challenged mice compared with the PBS-challenged mice (Fig. 1B).
ASC treatment inhibited AHR, lung inflammation, and mucus production induced by an allergen challenge
To evaluate the inhibitory effects of ASC, the ASC was injected intravenously before an allergen challenge in the mouse asthma model. Methacholine AHR was significantly reduced in the OVA-challenged mice treated with ASC compared with the OVA-challenged mice in the sham-treated group (P < 0.05; Fig. 2).
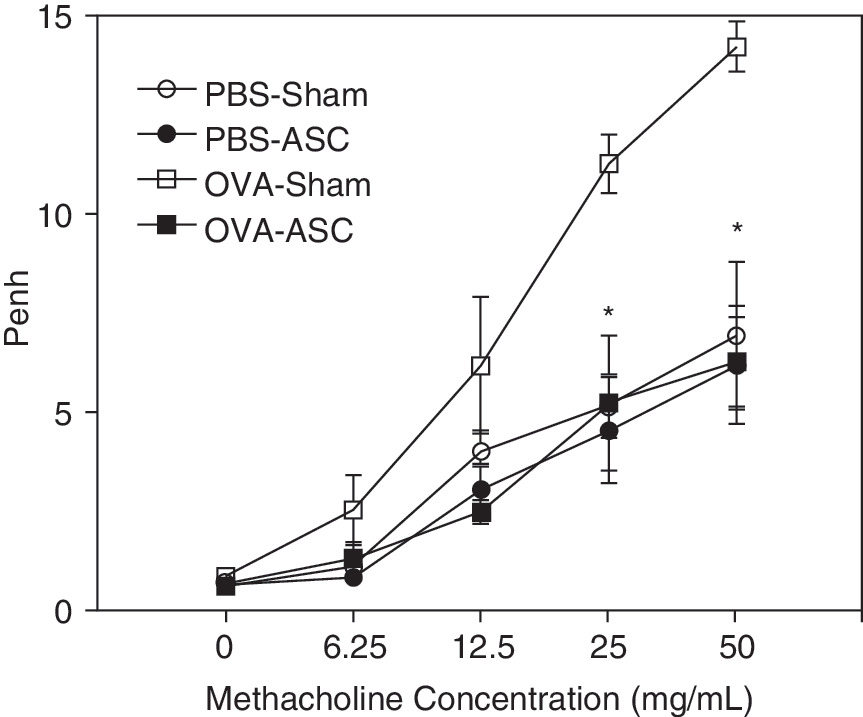
Effects of adipose-derived stromal cells (ASC) on airway hyperresponsiveness (AHR). Airway hyperresponsiveness was assessed in conscious, unrestrained mice using noninvasive whole-body plethysmography at 24 h after the last challenge. Methacholine AHR was significantly reduced in the ovalbumin (OVA)-challenged mice treated with ASC compared with the sham-treated group. These are representative results from 1 of 5 experiments (n = 5 each group per experiment). *P < 0.05 compared with OVA-challenged and sham-treated mice.
An examination of the inflammatory cells in the BAL fluids revealed that the OVA sensitization and challenge induced a significant increase in the total cell numbers, and in the proportion of eosinophils. However, the BAL cellularity was significantly lower in the OVA-challenged mice treated with the ASC than in the OVA-challenged mice in the sham-treated group (P < 0.05; Fig. 3A). The lung histology showed that intense lung inflammation was composed of a mixture of eosinophils, neutrophils, and mononuclear cells in the OVA-challenged mice compared with the sham-treated group. Even though there was a significant amount of peribronchial and perivascular infiltrate in the OVA-challenged and ASC-treated mice, the lung inflammation, especially the eosinophilic infiltration, was significantly reduced compared with the sham-treated mice (Fig. 3B). Moreover, PAS staining of the lung tissue showed that the mucus production was also markedly decreased in the ASC-treated mice compared with the sham-treated mice (Fig. 3B). The scores for the peribronchial and perivascular inflammation were significantly decreased in the ASC-treated mice compared with the sham-treated mice (P < 0.05; Fig. 3C). These results suggest that the ASC inhibited antigen-induced inflammation in the lung, including the influx of eosinophils and mucus production, in this mouse asthma model.
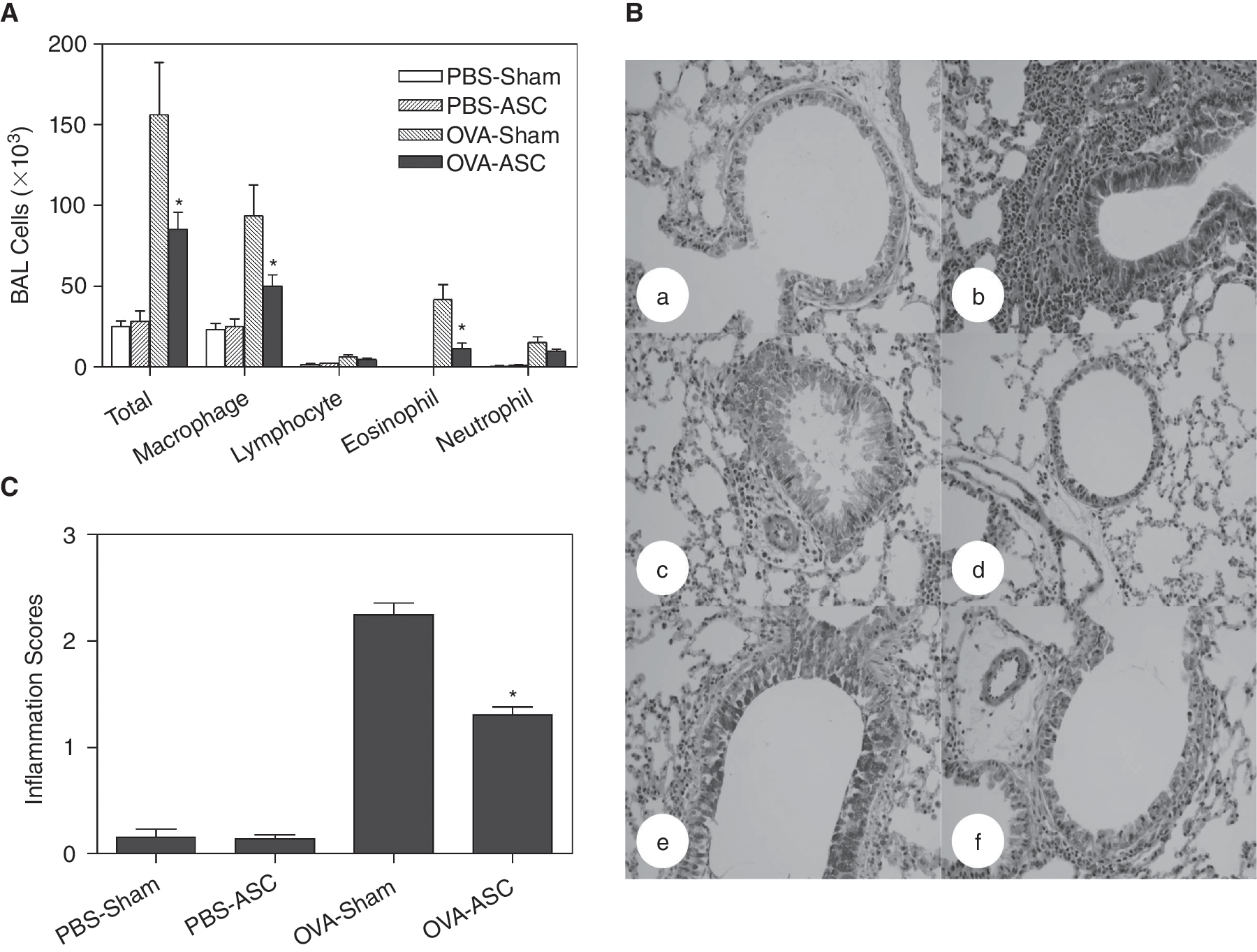
Effect of adipose-derived stromal cells (ASC) on lung inflammation. The numbers of each cellular component of bronchoalveolar lavage fluid (BALF) and lung inflammation were evaluated at 48 h after the last challenge. Inflammatory cell numbers in BALF (
ASC treatment decreased the IL-4 and IL-5 levels, but increased IFN-γ level in BAL fluid
We evaluated the immunomodulatory effects of ASC in the mouse asthma model. Bronchoalveolar lavage IL-4, IL-5, and TGF-β1 levels were significantly reduced in the OVA-challenged mice treated with ASC compared with the OVA-challenged mice in the sham-treated group (P < 0.05; Fig. 4). However, IFN-γ level in BAL fluid (BALF) was significantly increased in the OVA-challenged mice treated with the ASC compared with the sham-treated mice (P < 0.05; Fig. 4).
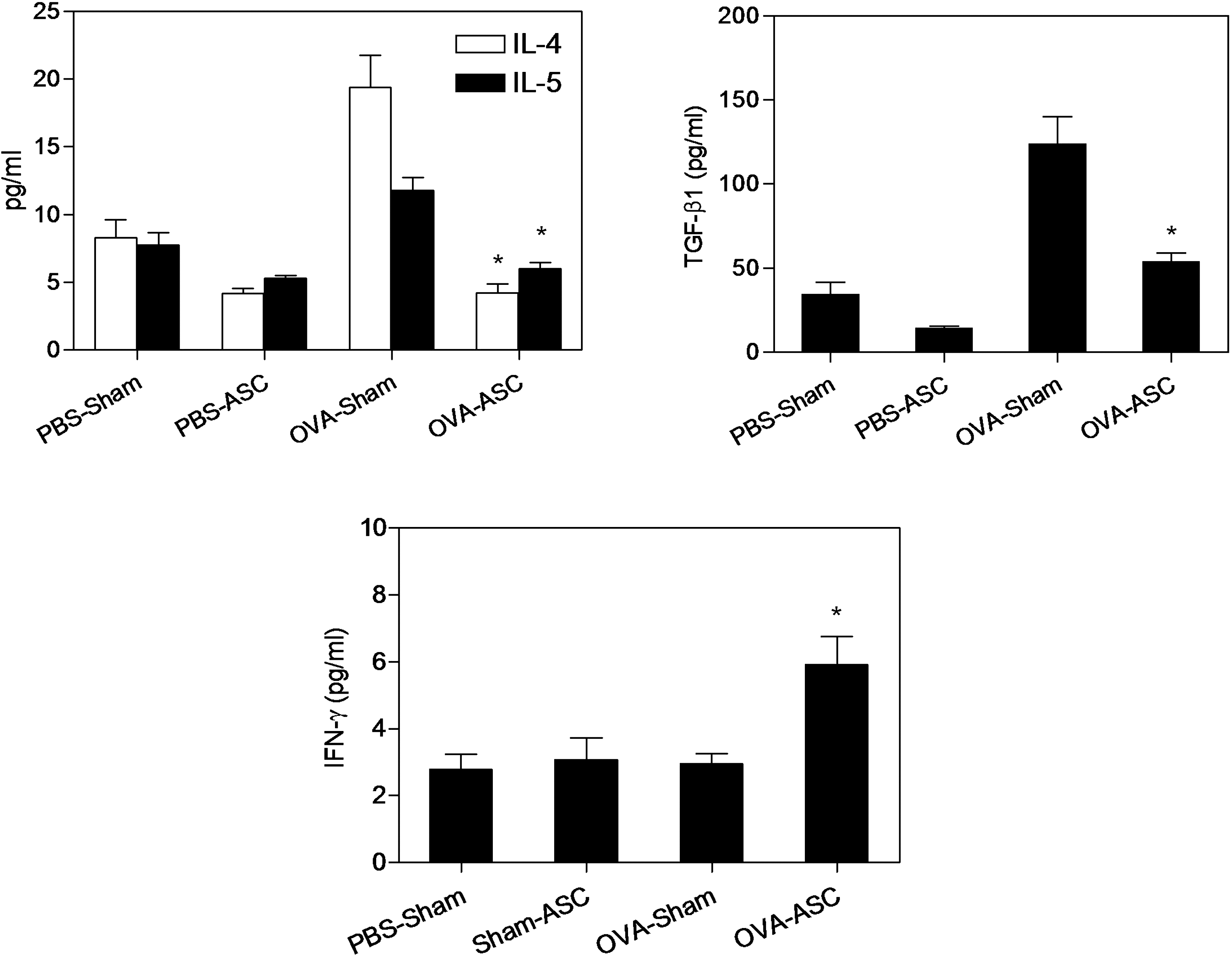
Effects of adipose-derived stromal cells (ASC) on bronchoalveolar lavage (BAL) cytokine levels. Bronchoalveolar lavage sampling was performed at 48 h after the last challenge. Cytokine levels in BAL fluid (BALF) were measured using enzyme-linked immunosorbent assay (ELISA). Interleukin (IL)-4, IL-5, and transforming growth factor (TGF)-β1 levels in BALF were significantly reduced after ASC treatment; however, interferon (IFN)-γ level in BALF was increased after ASC treatment. These are representative results from 1 of 5 experiments, each performed in triplicate (n = 5 each group per experiment). *P < 0.05 compared with OVA-challenged and sham-treated mice.
ASC treatment decreased IL-4+ CD4+ T cells in BALF and spleen
IL-4+ CD4+ T cells in BALF were decreased in the OVA-challenged mice treated with ASC (Fig. 5A); however, IFN-γ+ CD4+ T cells in BALF were increased in the OVA-challenged mice treated with ASC compared with the OVA-challenged sham-treated mice (Fig. 4). IL-4+ CD4+ T cells and IL-4+ CD8+ T cells in spleen were decreased in the OVA-challenged mice treated with ASC (Fig. 5B). These results suggest that ASC down-regulates the Th2 response to OVA in the OVA-induced asthma model.

Effects of adipose-derived stromal cells (ASC) on intracellular cytokine staining of T cells from (
Discussion
The multipotent nature of MSC makes them promising candidates for cell-based therapy. Mesenchymal stem cells isolated from the BM and adipose tissues have the same morphology, phenotype, in vitro differentiation capability, and similar gene expression profiles in humans [22]. Furthermore, both BM-MSC and ASC have immunosuppressive properties [13,14]. Because of the difficulties in obtaining sufficient amounts of autologous stem cells from the BM, ASC from allogeneic donors might provide an alternative source of stem cells for therapeutic use.
The results of this study demonstrated that migration of systemically administered ASC into lung tissue occurs at low levels in normal mice; however, the migration significantly increased in response to an OVA challenge in the OVA-induced mouse asthma model. We also showed that intravenous administration of ASC significantly inhibited the eosinophilic lung inflammation in the mouse asthma model. Moreover, this inhibition of ASC was accompanied by the down-regulation of IL-4, IL-5, and TGF-β1 production in BALF and decreased IL-4+ CD4+ T cells in the BALF and spleen. These results suggest that ASC might be a useful option for the treatment of asthma.
A previous study showed that MSC are distributed at varying levels in different organs [23]. Mesenchymal stem cells appear to migrate to sites of injury, suggesting that they are responsive to the local microenvironment and detect signals associated with inflammation and hypoxia [6]. After fibrotic tissue injury, MSC have been shown to migrate to the injured lung [24]. These findings suggest that MSC might provide an ideal cell source for repair of injured organs including the lung. The results of this study showed that the ASC were more intensely distributed in the lung in the OVA-sensitized and challenged mice compared with the PBS-sensitized and challenged mice. These results suggest that ASC migration into the lung was enhanced by the response to allergen exposure in the OVA-induced mouse asthma model. At present, the mechanism by which OVA-induced allergic inflammation augments ASC migration into the lung is unclear. However, release of cytokines by inflammatory cells may influence ASC migration. In addition, increased airway mucosal blood flow and microvascular leakage in the asthma mouse model may also induce increased ASC migration into the lung tissue.
MSC have recently been reported to have anti-inflammatory effects in experimental models of acute pulmonary and renal injury [24,25]. In addition, MSC have been shown to cause attenuation of experimentally induced autoimmune encephalomyelitis [11], and multiple sclerosis through the induction of T-cell anergy. Mesenchymal stem cells are emerging as a therapeutic modality in various inflammatory diseases including asthma. Weiss et al. found that adult mouse BM-derived MSC attenuate Th2-mediated inflammation and AHR as well as the number of eosinophils in BALF after OVA challenge in an asthma mouse model [26]. The anti-inflammatory effects were seen with both syngeneic and allogeneic BM-MSC administration, consistent with the concept of BM-MSC immunoprivilege. However, there are no data on the immunosuppressive effects of ASC in allergic asthma. Asthma is a chronic inflammatory airway disorder [1]. Allergen-specific Th2-driven allergic responses are important in the development of asthma [2]. In this study, the ASC treatment reduced eosinophilic inflammation and inhibited IL-5 production in BALF, an important cytokine involved in eosinophilic inflammation in asthma. Transforming growth factor-β, an important downstream molecule of eosinophilic inflammation in asthma, was reduced in the BALF after ASC treatment. In addition, the IL-4+ CD4+ cells in the BALF and spleen were also reduced after ASC treatment in the mouse asthma model. Furthermore, the IFN-γ level in the BALF and IFN-γ+ CD4+ T cells were significantly increased in the OVA-challenged mice treated with ASC. In our previous study, IgE and the IgG1/IgG2a ratio, which represent the Th2 immune response, were significantly decreased by ASC treatment in a mouse allergic rhinitis model [21]. In contrast, OVA-specific IgG2a levels, which indicate a Th1 immune response, were significantly increased by ASC pretreatment [21]. These findings suggest that ASC had immunomodulatory effects in the mouse asthma model, one of which was down-regulating the Th2 response and inducing a shift to a Th1 response. Despite the general consensus that MSC inhibit T-cell proliferation, there is little known on the exact molecular mechanisms responsible for this effect. Cell–cell contact and soluble factors such as TGF-β1 and prostaglandin E2 are thought to be responsible for this effect [27,28]. In part, T-cell suppression by MSC appears to depend on cross talk between the 2 cell populations, leading to the production of IFN-γ with increased indoleamine 2,3-dioxygenase activity, which in turn inhibits the proliferation of activated T or NK cells [29]. Moreover, the effects of MSC on T-cell activation might differ according to the specific microenvironment. The exact mechanism responsible for the inhibitory effects of ASC, in the mouse asthma model, requires further elucidation.
In summary, this is the first study to show the beneficial effects of ASC on allergic lung inflammation induced by an allergen challenge in mice. The findings show that ASC had inhibitory effects, decreased AHR, lung inflammation, mucus production, and Th2 cytokine productions in BALF in the mouse asthma model. These results suggest that ASC may have immunomodulatory effects on allergic airway inflammation, and might be potentially useful as a new candidate therapeutic agent for the treatment of asthma in the future.
Footnotes
Acknowledgment
This study was supported by Medical Research Institute Grant (2007–19), Pusan National University.
Author Disclosure Statement
No competing financial interests exist.