
Abstract
We have introduced 1 to 2 copies of a deletion mutant (βΔC) of the human retinoic acid receptor β into mouse embryonic stem (ES) cells. The βΔC-expressing cells were 10 to 100 times less sensitive to RA-induced differentiation in comparison with their parental cells. In the presence of 10−7 M RA in monolayer culture, they showed no growth arrest or differentiation, but remained pluripotent. Embryoid bodies (EBs) derived from βΔC-expressing cells differentiated into cardiomyocytes rather than neurons after treatment with 10−6 M RA, and became neurons upon exposure to 10−5 or 10−4 M RA. Remarkably, after 10 passages of continuous culture in the presence of 10−7 M RA, they still were able to form chimeras after injection into blastocysts. These data suggest that appropriate levels of normal retinoid receptors are crucial for lineage-specific differentiation of mouse ES cells in vitro. The βΔC mutant protein may prove to be useful in promoting “stemness” of ES cells in culture.
Introduction
R
One of the 3 RAR genes, RARβ, is autoregulated and strongly induced by RA in EC/ES cells and during early mouse embryogenesis [8 –11]. Previously, we have established stable ES clones expressing a lacZ reporter gene under the control of a 1.8 kb mouse RARβ2 promoter containing a RARE [11]. The reporter gene was expressed in a similar manner to the endogenous RARβ2 gene during ES cell differentiation in vitro and during embryonic development in vivo [11]. In the current study, a subclone, 2E5-E9, expressed the lacZ transgene in all cells after 2 days of RA treatment (10−8 M), was used as a reporter cell line to monitor RA concentration and ES cell differentiation.
RARs and RXRs are members of the steroid/thyroid/vitamin D receptor superfamily, and bind to RAREs in heterodimers. A deletion mutant of human RARβ (βΔC) lacking the DNA-binding domain was shown to act in a dominant negative fashion and to repress RA-dependent RARβ2 gene transactivation in transient transfections [12]. To further investigate roles of retinoid receptors during ES cell differentiation, we have constitutively expressed 1 to 2 copies of the βΔC in the 2E5-E9 cells under the control of the SV40 early promoter. The βΔC-expressing cells displayed a significant increase of cell proliferation and a reduction of apoptosis in the presence of RA, and remained as pluripotent stem cells in the presence of 10−7 M RA for over 10 passages. They could be differentiated into mesodermal (instead of ectodermal) derivatives under 10−6 M RA, and could be induced to become neurons in the presence of 10−5 or 10−4 M RA. Remarkably, 10−7 M RA-treated βΔC-expressing cells remained pluripotent in vivo and formed chimeric embryos after blastocyst injection. These data suggest that appropriate levels of RAR/RXR proteins are essential for lineage-specific cell differentiation in vitro.
Materials and methods
Cell lines and culture conditions
The 2E5 clone was established by transfecting E14 ES cell line of 129 mouse origin with a mRARβ2-lacZ-Neo cassette [11] (Fig. 1A). A subclone, 2E5-E9 expressing β-gal in all cells after 2 days of RA treatment (Fig. 1B) was chosen as parental cells for this study. The cells were electroporated with a dominant negative human RARβ mutant, βΔC, lacking the coding sequence of the DNA-binding domain [12]. The βΔC was under the control of a SV40 early promoter coupled to PGK-hygromycin B resistance gene (Fig. 1C). Nineteen ES clones were initially isolated after 2 weeks in the presence of G418 (200 μg/mL) and hygromycin B (100 μg/mL). Two clones, D1 and D6, containing 1–2 copies of the βΔC transgene (Fig. 1D and 1E) were extensively investigated and compared with the parental 2E5-E9 cells. Cultures in the presence of 10−7 M RA for 10 passages were designated as D1RA and D6RA, respectively. The 2E5-E9, D1, D6, D1RA, and D6RA cells were maintained feeder-free on gelatinized (0.1%) plates in buffalo rat liver cell-conditioned media as described [13], and passaged at ∼70% confluency by trypsinization.
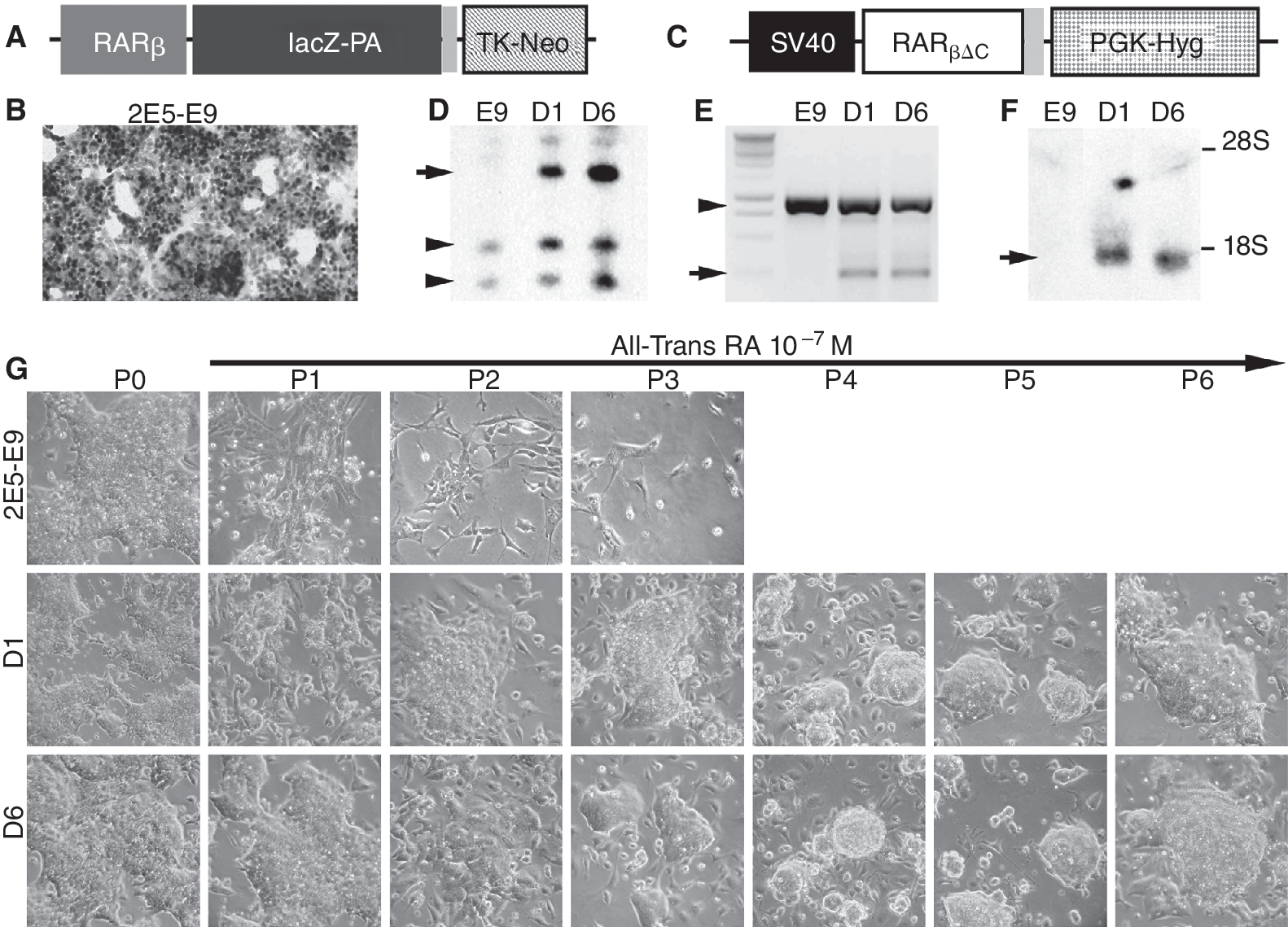
Embryonic stem (ES) cells expressing βΔC retain ES cell morphology in the presence of 10−7 M RA. (
Embryoid body (EB) formation and RA-induced differentiation
The protocols of EB formation and differentiation were described previously [13]. In brief, cultures were trypsinized and diluted to 20,000 cells/mL. Hanging drops (20 μL each, ∼400 cells) were cultured for 2 days on nonadherent culture dishes in EB growth medium, and EBs were washed and grown for one additional day with shaking at 50 rpm. EBs were subsequently treated with RA for 3 days (RAd3) by shaking on nonadherent 10-cm Petri dishes in fresh EB growth medium with all-trans RA at specified concentrations. The RAd3 EBs were harvested and plated on culture dishes precoated with poly-
Southern hybridization and polymerase chain reaction
Genomic DNA was extracted from cultured cells by Proteinase K digestion. Southern hybridization was done with 10 μg of genomic DNA (digested with SacI and BamHI) and hybridized to 32P-labeled human RARβ2 cDNA probe encoding the A/B domains, which detected a 1.2-kb fragment from the βΔC transgene. The transgene copy number was semiquantified based on polymerase chain reaction (PCR) product ratios. PCR was performed with primers RARβFor (5′-CTAAATCGAACTCAGATGCACAATGC-3′) and RARβRev (5′-CTTTGGAAACATGTGAGGCTTGC-3′) for 16, 18, 20, and 22 cycles, which amplified a 985-bp fragment from endogenous mouse RARβ and a 255-bp DNA from the βΔC transgene. The intensity of PCR bands was quantified by ImageJ and adjusted with their lengths. In addition, primers 5′-TCCGTGTGACAGAAGTGGTAGG-3′ and 5′-ACCTACATTGAGCAGTATGCCG-3′ were used to amplify a 630-bp fragment from mouse RARβ2 promoter, and primers 5′-TCCGTGTGACAGAAGTGGTAGG-3′ and 5′-TCTTCGCTATTACGCCAGCTGG-3′ to amplify a 417-bp band from the mRARβ2-lacZ transgene specifically.
Northern hybridization and RT-PCR
Total RNA was extracted from cultured cells using Trizol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's instruction. DNase I (RNase-free; Invitrogen) treatment was applied and RNA concentration was determined with the Quant-It Ribogreen RNA Assay Kit (Invitrogen). Northern blotting was carried out with 20 μg of total RNA and hybridized to 32P-labeled human RARβ2 cDNA, which detected a 1.3 kb of transgene mRNA just below the 18S ribosomal RNA. For reverse transcription, single-strand cDNA was synthesized from 2 μg RNA using a Reverse Transcription System Kit (Promega, Madison, WI) at 42°C for 1 h. Expression of βΔC and endogenous RARβ was carried out by RT-PCR using the same pair of primers (5′-GATGTTCTGTCAGTGAGTCC-3′ and 5′-CTTTTTCTTGTTCCTGTCATTCC-3′) flanking the deleted DNA-binding domain, which amplified 462-bp fragment from the endogenous RARβ and 279 bp from βΔC transcripts. RT-PCR of HPRT was used as a control (352 bp, 5′-CCTGCTGGATTACATTAAAGCACTG-3′, 5′-GTCAAGGGCATATCCAACAACAAAC-3′). Quantitative Real-Time PCR reagents were supplied by Qiagen House (Crawley, UK) and Applied Biosystems (Woolston, UK). PCR reactions were carried out using TaqMan® PCR probe sets (Applied Biosystems) coupled with Lightcycler 480 Probe mastermix (Roche Diagnostics, Indianapolis, IN). The TaqMan primer sets included Mm00450205_m1 for Nestin (72 bp, Tm = 60°C), Mm00485230_m1 for MAP-2 (91 bp, Tm = 60°C), Mm00657783_m1 for Nkx2.5 (117 bp, Tm = 60°C), and primers (5′-TCTGCGAAATAGACGTTCGGGCTTGC-3′ and 5′-GCCATCGTTCTCGCATGGGTTAGGG-3′) for Pref-1 (305 bp, T m = 62°C). All quantitative RT-PCR were analyzed in duplicates and repeated with templates from 3 independent experiments. Quantification was performed using the Opticon 3 software.
Immunocytochemistry
Immunocytochemistry was carried out in a dark-humidified chamber at room temperature. Cultures on 24-well plates were fixed with 1% paraformaldehyde (Sigma, St. Louis, MO) and 0.05% glutaraldehyde (Sigma) in phosphate-buffered saline (PBS) for 15 min, washed in PBS for 3 × 10 min, and incubated overnight with a primary antibody. After 3 × 10 min washing in PBS, cells were incubated for 2 h with secondary antibodies conjugated to fluorophores. They were washed for 3 × 10 min, and counterstained for 30 min with the DNA-binding dye Hoechst 33342 (5 μg/mL; Invitrogen, Carlsbad, CA). The primary antibodies included mouse anti-Nestin (DHSB, 1:10), mouse anti-Vimentin (DHSB, 1:100), rabbit anti-β-Tubulin III (Tuj1, 1:1,000), rabbit anti-MAP-2 (1:400; Sigma), rabbit anti-GFAP (DAKO, 1:1,000), and mouse anti-Cardiac Myosin Heavy Chain (1:200; Abcam, Cambridge, MA). Secondary antibodies (Invitrogen) included Alexa Fluor 488 donkey anti-rabbit IgG, Alexa Fluor 594 donkey anti-mouse IgG, Alexa Fluor 488 donkey anti-mouse IgG, Alexa Fluor 594 donkey anti-rabbit IgG, Alexa Fluor 594 donkey anti-goat IgG, and Alexa Fluor 594 donkey antimouse IgM.
Proliferation and apoptosis assays
A pulse of 5-bromodeoxyuridine (BrdU; BD Biosciences, San Jose, CA) at a final concentration of 10 μM was applied to the cultures for 1 h and cultures were fixed with 1% paraformaldehyde (Sigma, St. Louis, MO) and 0.05% glutaraldehyde (Sigma) in PBS for 2 min. After 3 washes in PBS, cells were treated with 2 N HCl for 45 min at RT, neutralized with 0.1 M NaB4O7 for 10 min, and subsequently incubated with pepsin solution (0.04% w/v in 0.01 N HCl; Sigma) for 30 min at room temperature. Cells were washed in PBS before application of anti-BrdU antibody. Incorporation of BrdU was assayed according to the manufacturers' instructions (BD Biosciences). To determine programmed cell death during RA differentiation, cells were fixed and examined for the percentage of active Caspase-3-positive cells with rabbit anti-Caspase-3 antibody (1:100; Cell Signalling, Danvers, MA) according to the manufacturers' instructions.
Blastocyst injection and X-gal staining
MF1 females (∼8 weeks) were superovulated and timed matings carried out. E3.5 post-coitum blastocysts were injected with ∼15 ES cells/each, and transferred into E2.5 pseudopregnant recipients mated with vasectomized males. Embryos were recovered 8–10 days later. X-gal staining of cells and embryos was processed as previously described [11].
Results
ES cells expressing a mutant RARβ (βΔC) are resistant to 10−7 M RA-induced differentiation
Previously, we have electroporated mouse ES cells with a lacZ reporter gene under the control of a 1.8 kb mouse RARβ2 promoter containing the RARE (Fig. 1A) [11]. A subclone, 2E5-E9, which expressed β-gal in all cells after 2 days of RA treatment and gave rise to healthy germline transmittable chimeras [11], was chosen as a reporter cell line for this study (Fig. 1B). They were subsequently transfected with a dominant negative RARβ mutant (βΔC) [12], lacking the coding sequence of the DNA-binding domain (Fig. 1C). Integration of the βΔC transgene was determined by southern hybridization (Fig. 1D) and PCR (Fig. 1E). Karyotyping was performed on 2 clones D1 and D6 containing 1–2 copies of the βΔC transgene (Fig. 1D and 1E) with their parental 2E5-E9 cells. The majority of cells in clones E9 (20/25, see Supplementary Fig. 1; Supplementary materials are available online at
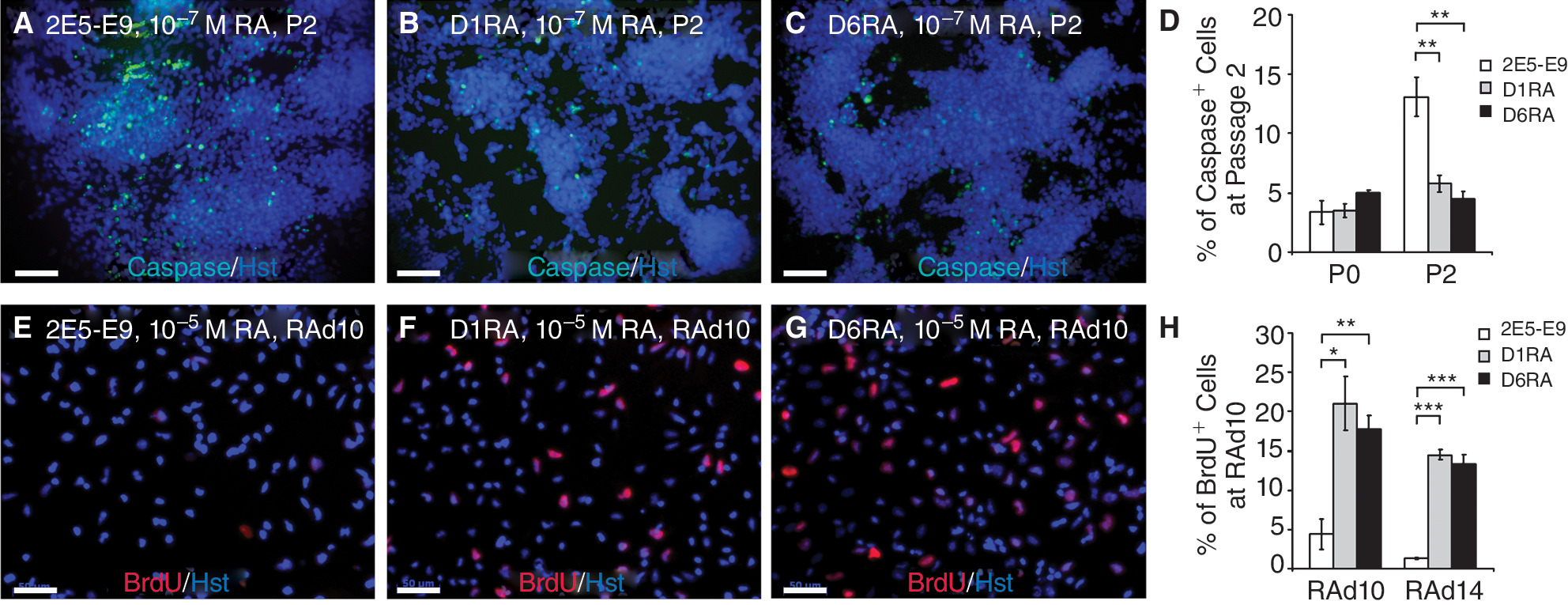
Reduced apoptosis and increased proliferation of D1RA and D6RA cells in the presence of RA. (
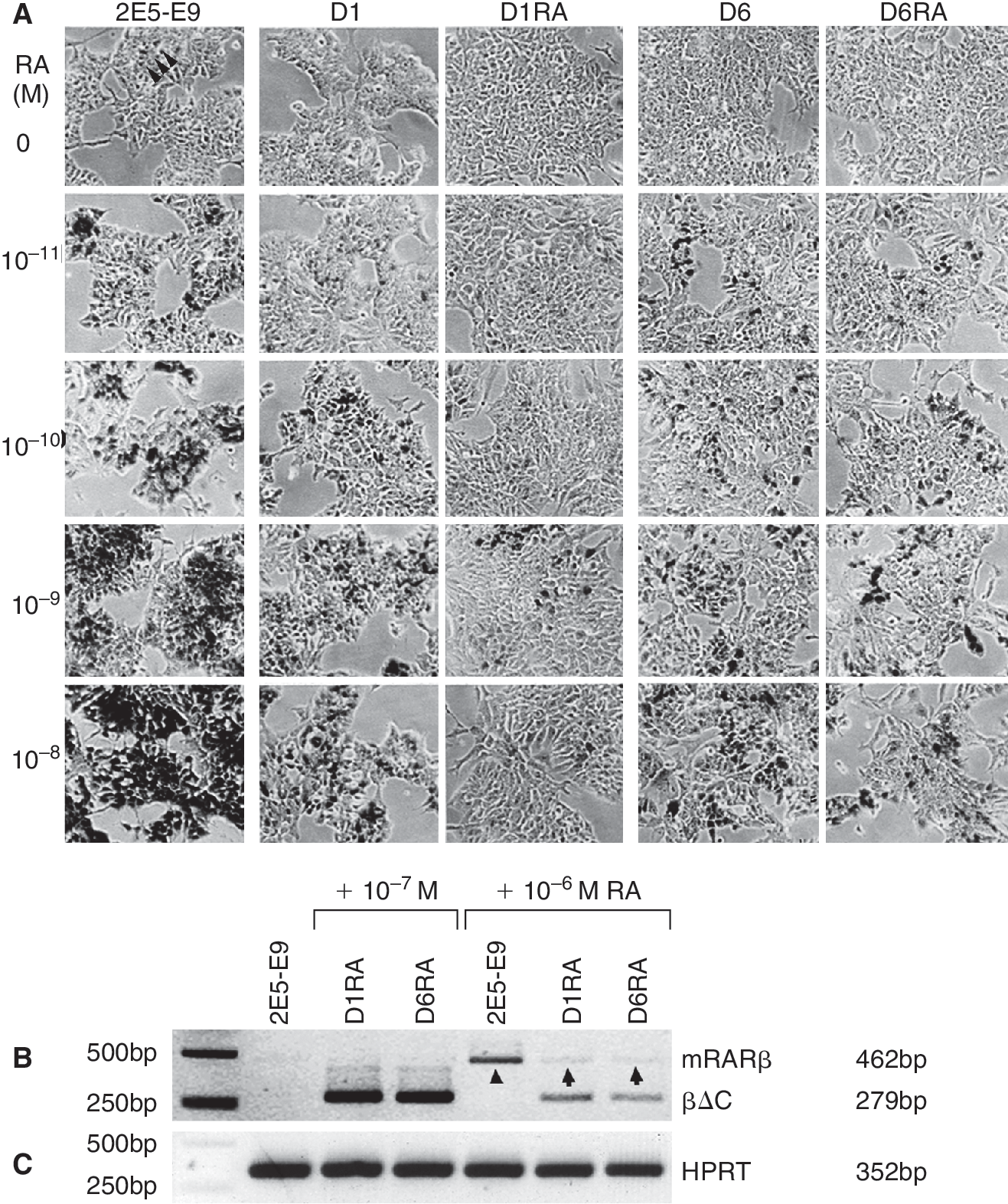
Reduced induction of RA-responsive gene expression in D1RA and D6RA cells. (
To examine the effect of the βΔC mutant on RA-induced differentiation, we added 10−7 M RA to the culture medium for monolayer differentiation. The majority of the parental cells began to differentiate in passage 1 (P1, Fig. 1G). Their growth was severely inhibited, and cultures were completely lost by passage 4 (P4, Fig. 1G). However, D1 and D6 cells retained stem cell morphology and grew in colonies in the presence of 10−7 M RA (Fig. 1G), showing that cells expressing 1–2 copies of βΔC under these conditions were resistant to 10−7 M RA-induced differentiation.
βΔC increases cell proliferation and reduces apoptosis in the presence of RA
Normally, RA-induced differentiation in 2E5-E9 cells was accompanied by cell detachment and death of a sizable cell fraction upon passaging. To determine if RA decreases cell viability by apoptosis, we tested the Caspase-3 activity by immunocytochemistry (Fig. 2A–2C). Cells, cultured in the absence of RA (P0), or in the presence of 10−7 M RA for 1 or 2 passages, were fixed and stained for active Caspase-3, and proportions of Caspase-3-positive cells were calculated. In the absence of RA, there were low but comparable rates of cells undergoing apoptosis among the 3 clones (3.5%–5.0%). A significant increase of apoptotic cells was observed in 2E5-E9 cultures after 2 passages of RA treatment (13.1% ± 16.0%). However, in both D1 and D6 cells, the proportions of Caspase-3-positive cells remained relatively stable (4.5%–5.8%). These data suggest that the βΔC expression counteracts RA-induced apoptosis.
To evaluate the effect of βΔC expression on cell proliferation, embryoid bodies (EBs) from these 3 ES clones were programmed for differentiation with 10−5 M of RA and examined for BrdU incorporation at RAd10 (Fig. 2D–2F). A small proportion of 2E5-E9-derived cells (4.4% ± 1.9%) were BrdU-positive. However, significantly higher proportions of BrdU-stained cells were found in D1RA (21.0% ± 3.4%) and D6RA (17.7% ± 3.4%) cells, demonstrating that 1–2 copies of βΔC expression significantly increased cell proliferation at 10−5 M of RA.
βΔC expression attenuates RARβ induction
RARβ gene expression is undetectable in ES cells, but rapidly induced by RA. We examined the effect of βΔC on RA responsiveness by lacZ expression after 48 h of RA treatment (1 μM). In contrast to robustly induced β-gal activity in all 2E5-E9 cells, reduced proportions of X-gal staining were found in D1 (∼30%) and D6 cells (∼70%). We have subsequently treated cells for 48 h with different RA concentrations (10−11 to 10−8 M, Fig. 3A). The β-gal activity in 2E5-E9 cells was induced in a RA concentration-dependent manner, and the majority of the cells strongly expressed mRARβ2-lacZ in the presence of 10−8 M RA (Fig. 3A). Under the same conditions, the β-gal activity was substantially decreased in D1 and D6 clones (Fig. 3A). The X-gal staining was further reduced in D1RA and D6RA cells after long-term exposure to 10−7 M of RA. To achieve comparable β-gal induction, 10-to 100-fold higher concentrations of RA are required for the dominant negative clones. Additionally, RT-PCR analysis was performed with RNA from 2E5-E9, D1RA, and D6RA ES cells before and after RA treatment (Fig. 3B). The RARβ mRNA was almost undetectable in 2E5-E9 cells cultured in the absence of RA, or in D1RA and D6RA cells maintained with 10−7 M of RA. RA treatment at 10−6 M for 3 days in the absence of LIF resulted in a significant up-regulation of endogenous RARβ expression in 2E5-E9 cells (arrowhead, Fig. 3B). Under the same conditions, RARβ induction in D1RA and D6RA cells was considerably reduced (arrows, Fig. 3B). These results clearly indicate that expression of the βΔC significantly attenuates induction of RA-responsive gene expression.
D1RA and D6RA cells differentiate into cardiomyocytes rather than neurons in the presence of 10−6 M RA
RA differentiates ES cells into cell types of 3 germ layer origins in an incubation time- and concentration-dependent manner [3]. High concentrations (∼1 μM) of RA during EB formation drive neuronal differentiation, whereas low RA concentrations (∼10 nM) induce cardiomyocyte and adipocyte differentiation [3]. To examine the differentiation potential of dominant negative ES clones, we used a RA-induction protocol similar to that previously shown to generate radial glia [14]. 2E5-E9, D1RA, and D6RA ES cells were plated onto nonadherent dishes for 3 days for EB formation (RAd0). RA (at 1 μM) was added to the suspension cultures for 3 days (RAd3). RA-treated EBs were transferred to poly-
As shown previously [13], 2E5-E9 EBs differentiated into neurons with clusters of cells migrating away from the EBs and formed a dense neuronal network with extensive neuronal processes (Fig. 4A). The neuronal identity was confirmed with neuronal markers (Figs. 4 and 5). 2E5-E9 derivatives widely expressed 3 neuronal markers Nestin (a neural progenitor marker, Fig. 4D), β-tubulin III (Tuj1, a pan-neuronal marker, Fig. 4D), and MAP-2 (another neuronal marker, Fig. 4G) at RAd10. A limited number of GFAP-positive cells were found at RAd10 (Fig. 4G), while significantly increased Map-2 mRNA was expressed in 2E5-E9-derived cultures by RAd14 (data not shown).
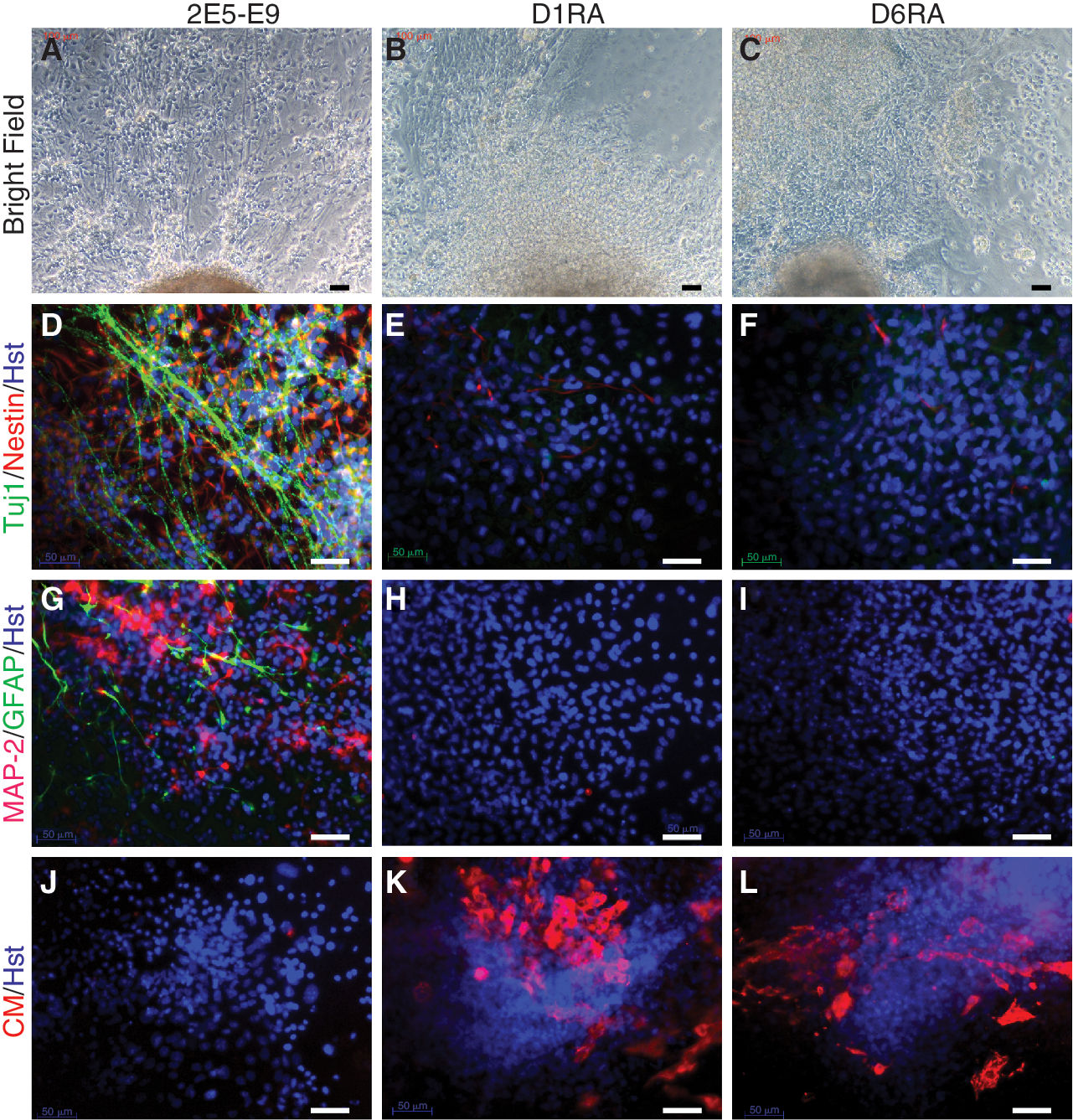
D1RA and D6RA cells differentiate into cardiomyocytes under 10−6 M RA. 2E5-E9, D1RA, and D6RA cells were programmed for differentiation with 10−6 M RA. At RAd10, cells were imaged in bright field (
In contrast, under 10−6 M RA, D1RA- and D6RA-derived cells did not show typical neuronal morphology (Fig. 4B and 4C), but developed increasing areas of cells with spontaneous, continuous, and regular beating during RAd7 and RAd14 (supporting information online videos SV1–SV6). They barely expressed any of the neuronal markers (Fig. 4E, 4F, 4H, and 4I), but strongly stained with anti-cardiac myosin heavy chain (Fig. 4K and 4L), a marker for cardiomyocytes (and responsible for the conversion of the chemical energy through the hydrolysis of ATP into mechanical force that results in contraction). 2E5-E9-derived cells, on the other hand, were negative for cardiac myosin heavy chain under these conditions (Fig. 4J), but did express the protein and induce beating muscle differentiation at a lower RA concentration (10−8 M RA, data not shown).
To further verify the nature of differentiated cells, we carried out quantitative RT-PCR with neuronal markers Nestin and MAP-2, and 2 additional mesodermal markers, Nkx2.5 (a transcription factor that initiates cardiac fate) and pre-adipocyte factor-1 (Pref-1, a member of the EGF transmembrane protein family highly expressed in pre-adipocytes of mesodermal origin). 2E5-E9-derived cells expressed Nestin and MAP-2 mRNA in a time-dependent manner from RAd7 to RAd14 (Fig. 5A and 5B), with no significant induction of Nkx2.5 and Pref-1 (Fig. 5C and 5D). In D1RA- and D6RA-derived cultures, Nestin and MAP-2 were not abundantly expressed throughout differentiation (Fig. 5A and 5B). However, Nkx2.5 was highly expressed in RAd7 and RAd10 (Fig. 5C), and Pref-1 mRNA was progressively induced and peaked at RAd14 (Fig. 5D). Taken together, these data clearly demonstrate that while 10−6 M of RA induces neuronal features in normal ES cells, it differentiates D1RA and D6RA cells into cardiomyocytes.
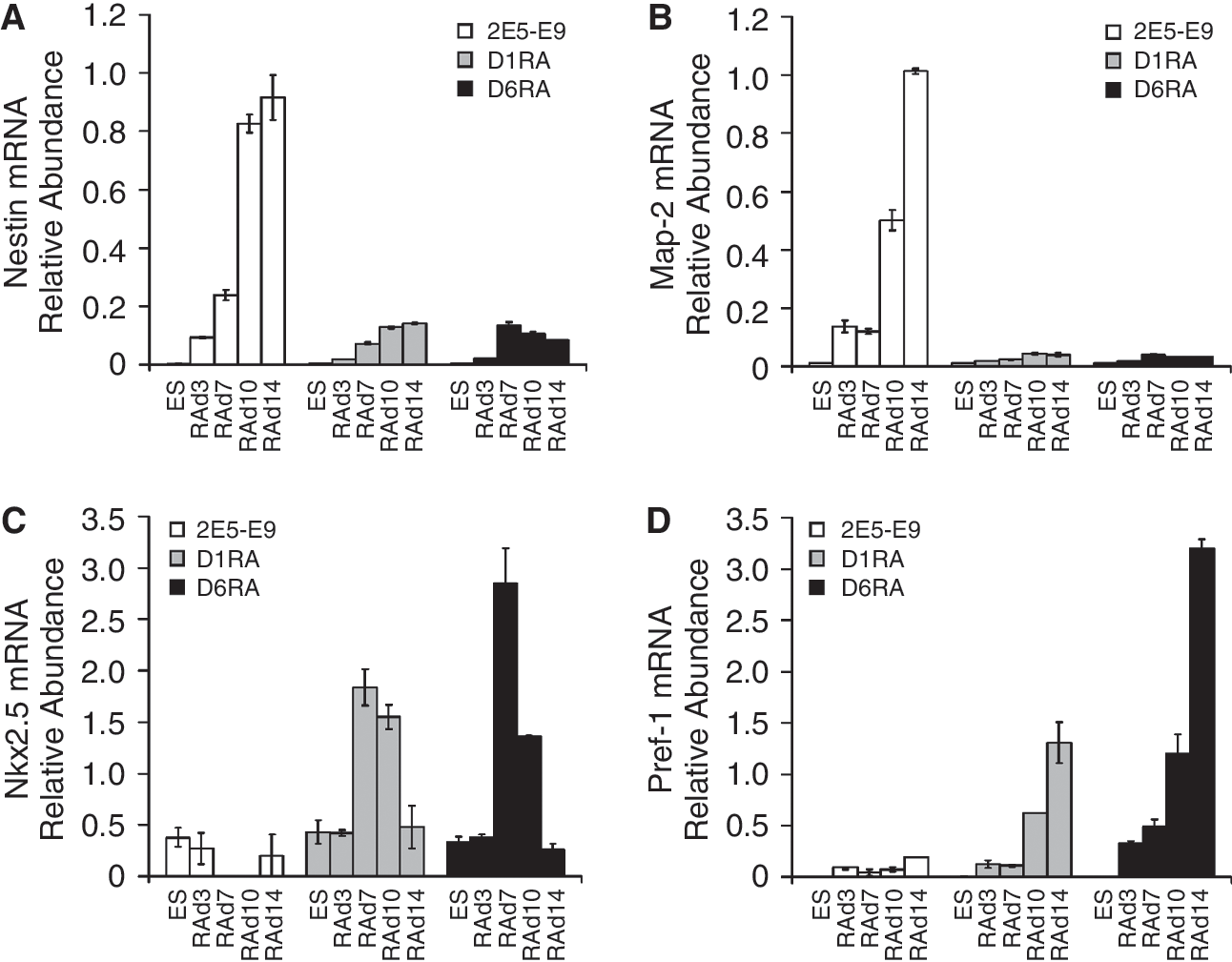
Expression of neuronal (Nestin and Map-2), cardiac (Nkx2.5), and adipocytic (Pref-1) markers. 2E5-E9, D1RA, and D6RA embryoid bodies (EBs) were treated with 10−6 M for 3 days (RAd3) and plated out for 4 (RAd7), 7 (RAd10), and 11 (RAd14) days. Total RNA was extracted and quantitative RT-PCR was carried out to determine the mRNA relative abundance for Nestin (
D1RA and D6RA cells can be differentiated into neurons, by 10−4 M RA
The finding that 10−6 M RA induced mesodermal differentiation in D1RA and D6RA cells, which is achieved by ∼10−8 M RA in parental ES cells, prompted us to examine their neuronal differentiation potential at 10−4 M RA (Fig. 6). After 7 days of RA induction, differentiating cultures from 2E5-E9, D1RA, and D6RA were morphologically indistinguishable; they all contained clusters of cells exhibiting a rosette-like morphology, characteristic of neuroepithelial or radial glia cells, with neurite-like processes (Fig. 6A–6C). These rosette-like clusters, also called radial arrays, were immunopositive for both Nestin and Tuj1, suggesting that these were neural progenitors committed to a neuronal fate (Fig. 6D–6F). Cells with long and fine processes were labeled only for Tuj1, and they appeared to be over-layered with the radial arrays.
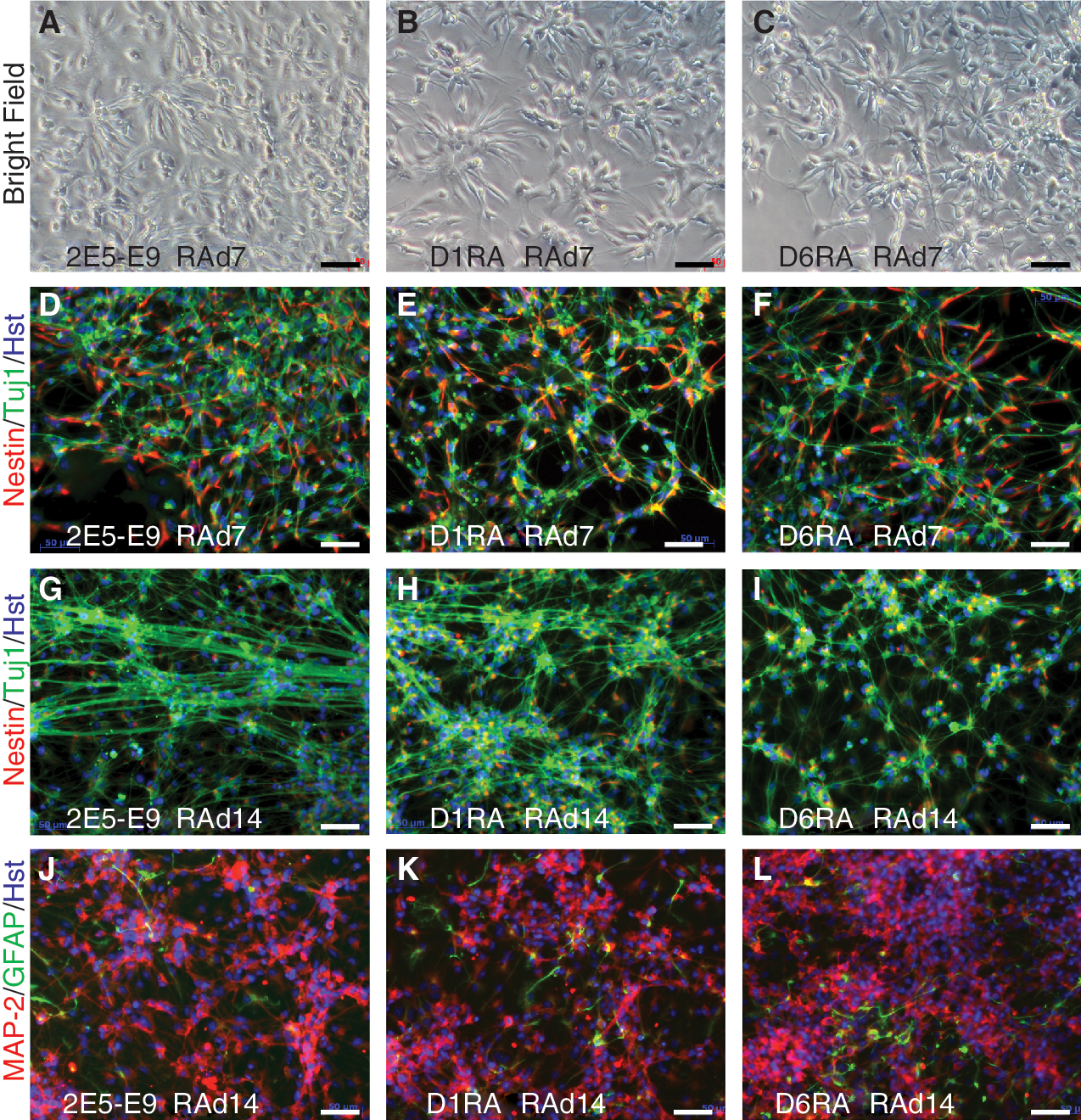
Neuronal differentiation of ES cells with 10−4 M RA. RA at 10−4 M was applied to EBs derived from 2E5-E9 (
Between RAd7 and RAd14, the cultures of all 3 cell lines formed more and more dense neuronal networks with increasing and elongating neurites. This increase was coincident with a decrease in the number of Nestin-positive cells (Fig. 6G–6I) and increasing staining of Tuj1. They also abundantly express Map-2, another neuronal marker (Fig. 6J–6L), with few cells stained by anti-GFAP antibody, a marker for cortical radial glia cells and astrocytes. These data demonstrate that D1RA and D6RA ES cells are capable of neuronal differentiation, but require much higher RA concentrations.
Pluripotency of βΔC-transfected ES cells in vivo
The most important feature of mouse ES cells is that they can participate in embryonic development in vivo, following introduction into host embryos. D1RA and D6RA cells retained stem cell morphology in the presence of 10−7 M RA for at least 10 passages. They were then cultured for 2 passages in the absence of RA, and microinjected into blastocysts, to compare their developmental potential with 2E5-E9 cells in vivo. Embryos were dissected at ∼E10.5 post-coitum, fixed, and stained with X-gal. As we showed before [11], 2E5-E9 cells participated in all parts of embryos including the head region with no X-gal staining (Fig. 7E and 7F), but the X-gal staining was largely restricted to the developing spinal cord where the endogenous RARβ2 was expressed (Fig. 7A).
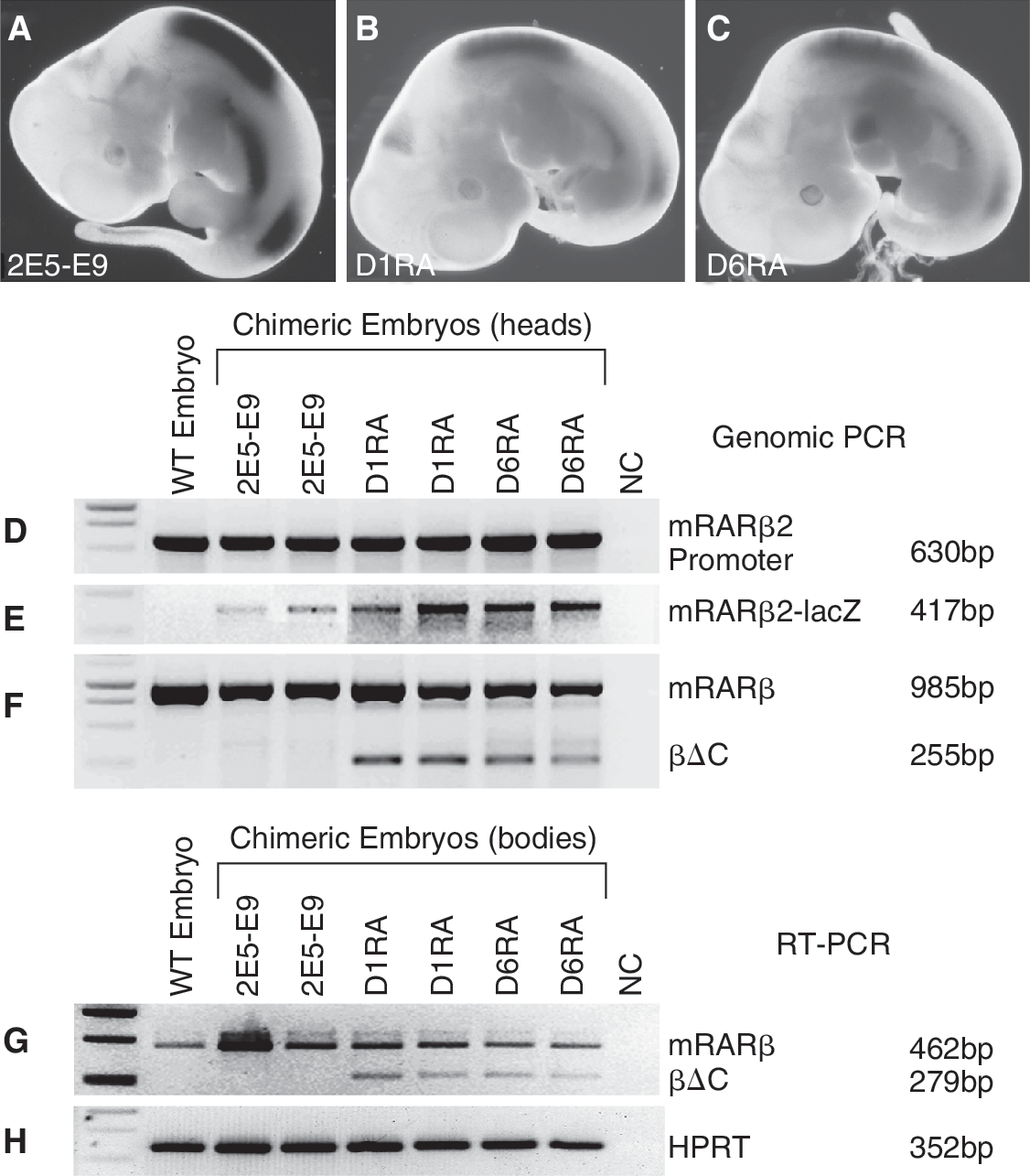
Participation of βΔC ES cells in embryo-genesis. X-gal staining of E10.5 embryos developed from blastocysts injected with 2E5-E9 (
Unexpectedly, D1RA and D6RA cells, after 10 passages in the presence of 10−7 M RA, could participate in development and form chimeric embryos (Fig. 7B and 7C). The X-gal staining patterns among the chimeric embryos resulting from microinjection of 2E5-E9, D1RA, or D6RA are largely the same. Strong β-gal activity was found in 2 regions of spinal cord close to the forelimb and hind limb buds, in addition to its expression in the developing eye and the lateral body wall. Consistent with weaker mRARβ2-lacZ induction in D1RA and D6RA cells in vitro, the intensity of staining in D1RA and D6RA chimeric embryos (Fig. 7B and 7C) was much weaker in comparison with 2E5-E9 chimeric embryos (Fig. 7A). D1RA and D6RA derivatives expressed βΔC in chimeric embryos (Fig. 7G). D1RA and D6RA chimeric embryos did not have gross abnormalities or tumor formation, indicating that D1RA and D6RA cells could be differentiated normally in an embryonic environment, and long-term treatment of 10−7 M RA did not appear to alter the pluripotency of D1RA and D6RA cells. This is the first time to our knowledge that RA-treated cells have been shown to form chimeras.
Discussion
We showed that ES cells expressing 1–2 copies of a deletion mutant of human RARβ, βΔC (lacking the DNA-binding domain), were resistant to 10−7 M RA-induced differentiation and remained as pluripotent stem cells in monolayer culture. EBs derived from βΔC-expressing cells differentiated into cardiomyocytes rather than neurons in the presence of 10−6 M RA, but became neurons after induction with 10−5 to 10−4 M RA. Remarkably, after 10 passages of culture with 10−7 M of RA, they retained multipotency in vivo and formed chimeras after being introduced into blastocysts. To our knowledge, this is the first report that cells are able to participate in normal embryonic development after prolonged RA treatment.
Previously, mutants of RARs or RXRs were generated and transfected into EC cells. They caused RA nonresponsiveness and repressed RA-inducible gene expression in some but not all studies. A deletion mutant of the human RARα encoding the N-terminal 186 amino acids was transfected to F9 EC cells, and 2.0%–3.4% of clones were resistant to 0.5 μM RA-induced growth arrest [15]. Such a mutant, comprising the DNA-binding domain but lacking the ligand-binding/dimerization domain, may occupy RAREs as inactive monomers, and thus prevent binding of active dimers. Later, a truncated RARα encoding the first 391 amino acids and lacking the last 70 amino acids was found to be responsible for RA nonresponsiveness in a naturally mutated P19 EC cell line, RAC65 [16,17]. Introduction of a human RARα encoding the N-terminal 394 amino acids of the receptor into P19 cells indeed blocked RA-induced differentiation in a similar way [9, 18]. A comparable human RARβ deletion mutant was characterized to dominantly repress RA-induced gene activation in transfected EC cells [12]. These mutants lack the C-terminal transactivation domain only. They can associate with wild-type receptors and bind to RAREs as inactive dimers [18].
We previously characterized a human RARβ mutant with an internal deletion of the DNA-binding domain, which also acted as a dominant negative mutant [12]. Consistent with this finding, a mutant mouse RXRβ lacking the DNA-binding domain caused resistance of RA-induced differentiation in P19 cells [19]. These mutants lacked the DNA-binding capacity, but were able to dimerize with wild-type RXRs to form inactive dimers [19]. Among the 3 RXR genes, RXRα was the main functional RXR subtype during embryogenesis [7] and RA-induced F9 EC cell differentiation [20]. Among the 3 RAR genes, RARα was the most abundant RAR expressed in EC [21,22] and ES cells [23], whereas RARβ was induced upon RA stimulation during neuroectodermal differentiation [21,22]. Induction of RARβ expression was sufficient and necessary to trigger commitment of mouse ES cells to adipocyte differentiation [24], whereas targeted deletion of RARβ resulted in the loss of RA-associated growth arrest in F9 cells [25].
Our study suggests that appropriate levels of normal RARs and RXRs are crucial for ES cell differentiation. Expression of 1–2 copies of the βΔC mutant is sufficient to alter threshold levels of the receptors required for gene activation and lineage-specific differentiation. It appeared that 10- to 100-fold increased RA concentrations were required to achieve mRARβ2-lacZ transgene induction and lineage-specific differentiation in βΔC-expressing cells, in comparison to their parental cells. Exposure of growing EBs to nanomolar RA was shown to induce mesodermal differentiation, whereas micromolar RA was required for neuronal differentiation [3]. This was also the case for the parental cells 2E5-E9 used in this study. However, the βΔC-expressing cells did not show growth arrest or differentiation after prolonged exposure to 10−7 M RA in monolayer culture, a condition that normally differentiated ES cells into a mixture of mesodermal and ectodermal cells. In the presence of 10−6 M RA, they became beating cardiomyocytes (instead of neurons) expressing mesodermal markers including cardiac myosin heavy chain, Nkx2.5 and Pref-1, which were induced by ∼10−8 M RA normally [3]. The βΔC-expressing cells retained the ability to become neurons, and this could be achieved by applying 10−5 or 10−4 M RA. These data suggest that βΔC mutant protein may effectively form inactive dimers and effectively compete for the availability of retinoids.
We also demonstrated that the βΔC mutant can effectively increase proliferation and attenuate RA-induced apoptosis. Spontaneous differentiation in prolonged culture was one of the major problems in the maintenance of stem cells including human origin, which hindered the isolation of pure and undifferentiated clonal human ES cell lines [26]. It was also suggested that the presence of minor populations of differentiated cells might lead to the complete loss of stem cells [27]. However, factors causing spontaneous differentiation remain unknown. It is worth noting that weak X-gal staining appeared in some of the 2E5-E9 cells containing mRARβ2-lacZ transgene under normal culture conditions (Fig. 3A). This indicated that a low level of RA might be present in culture medium. On the other hand, expression of 1–2 copies of βΔC was sufficient to maintain pluripotency of ES cells in vitro, in the presence of 10−7 M RA for over 10 passages. Although RAR/RXR genes are essential for differentiation of EC cells [20] and ES cells [24] in vitro, viable mice were developed with single and/or compound disruption of the RARs and RXRs, despite various phenotypes being associated with the knockouts [7]. Obviously, the in vivo cell differentiation process differs from the in vitro situation. When RA signaling is not available, other signaling pathways take over to ensure embryonic morphogenesis. Therefore, it is not completely surprising that ES cells expressing a mutant RAR or RXR are able to participate in embryogenesis in vivo. What is remarkable in the current study is that cells undergoing over 10 passages of culture in the presence of 10−7 M RA are still able to form chimeras after blastocyst injection. This demonstrates that production of the βΔC mutant protein from 1 to 2 copies of the transgene is sufficient to counteract 10−7 M RA to maintain their stemness. This study implies that βΔC protein may bind retinoids in culture medium. Supplement of culture medium with βΔC protein may prove to be efficient in preventing contaminated retinoids from entering into cells, and therefore benefit the maintenance of cultures in purer stem cell status.
Footnotes
Acknowledgments
We thank the UK Medical Research Council, NHS Grampian, University of Aberdeen and Tenovus Scotland for funding the project. We thank Mr. David AJ Stevenson, Ms. Doreen Massie, and Mrs. Helen Cran at NHS Grampian for the assistance of ES cell karyotyping.
Author Disclosure Statement
Authors indicate no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.